脂肪代谢重要基因网络对呼伦贝尔羊脂肪分布和脂肪酸组成的影响
2022-06-02郭月英赵丽华孙雪峰
要 铎,张 月,郭月英,苏 琳,赵丽华,孙雪峰,靳 烨,
(1.内蒙古农业大学食品科学与工程学院,内蒙古 呼和浩特 010018;2.内蒙古自治区质量和标准化研究院,内蒙古 呼和浩特 010018;3.乌拉特中旗畜牧工作站,内蒙古 巴彦淖尔 015300)
我国有丰富的绵羊品种和悠久的绵羊遗传育种历史,呼伦贝尔草原地处亚洲中部的蒙古高原,是世界著名的高原牧场,呼伦贝尔羊繁衍于此,根据其尾巴大小的不同分为“巴尔虎”和“短尾”两种品系。脂尾性状尤为重要,是一种逆境生存所必需的生物性状,而这两种尾型的呼伦贝尔羊在自然条件下均可度过呼伦贝尔的寒冬,且在同一饲养条件下具有不同的肉用品质,其机制值得进一步探究。从脂肪代谢相关基因角度探究脂肪沉积机制的技术手段已趋于成熟,其中脂肪量与肥胖相关基因(fat mass and obesity associated,)、-甲基腺苷甲基转移酶基因(methyltransferase 3,)、过氧化物酶体增殖剂激活受体基因(peroxisome proliferators-activated receptor,)、解耦联蛋白1基因(uncoupling protein 1,)、腺苷酸活化蛋白激酶基因(AMP-activated proteinkinase,)、乙酰辅酶A羧化酶基因(acetyl-CoA carboxylase,)和肉碱脂酰转移酶1基因(carnitine palmitate transferase 1,)对动物脂肪代谢具有重要影响,如和基因可影响不同肥胖表型猪的脂肪沉积,和基因可调控小鼠和猪的脂肪细胞分化,信号通路的激活可减少肉羊的肌内脂肪沉积。
上述基因并非单独存在,而是存在一定的基因网络(图1):和催化-甲基腺苷(-methyladenosine,mA)的去甲基化和甲基化,从而改变脂质相关基因的mRNA的加工、成熟和翻译,可通过胆固醇调节元件结合蛋白1c(sterol-regulatory element binding proteins 1c,)途径抑制介导的脂肪酸氧化,从而促进脂质贮存,也可通过改变表达增强CCAAT增强子结合蛋白(CCAAT enhancer binding proteins,/)的表达,促进脂肪沉积,另外,可通过抑制信号通路以减少骨骼肌中的脂质利用,同时可激活生热作用,共同发挥重要的脂肪代谢调节作用。该基因网络最终改变了动物的脂肪分布与脂肪酸组成,进而影响其肉制品的营养、风味、大理石花纹、胴体等级和加工效率。本研究选择呼伦贝尔羊两种品系为实验材料,探索脂肪代谢重要基因网络对其脂肪分布和脂肪酸组成的影响机制,结合前期的研究,旨在为改善羊肉品质和生产效率提供一定的理论支撑。

图1 脂肪代谢重要基因网络Fig.1 Important gene networks related to fat metabolism
1 材料与方法
1.1 材料与试剂
实验动物来源于内蒙古呼伦贝尔市鄂温克族自治旗羊场,其中巴尔虎羊和短尾羊各50 只,共计100 只。在相同的饲养管理条件下,以玉米秸、苜蓿和燕麦秸按质量比6∶6∶5混合饲料喂养,日摄入量约为500 g/只。从中选择5 月龄发育正常、健康无病且体质量相近的巴尔虎公羊与短尾公羊各10 只,共计20 只羊。现场屠宰并称质量,采集背最长肌、臂三头肌、股二头肌、尾部脂肪和胸腹部皮下组织样品置于-20 ℃冰箱或液氮中保存,用于后续脂肪分布与脂肪酸组成以及脂肪代谢相关基因表达的测定。
甲醇、正己烷(均为色谱纯),三氯甲烷、三氟化硼-乙醚络合物、氯化钠、氢氧化钠、无水硫酸钠、乙醚(均为分析纯) 国药集团化学试剂有限公司;37 种脂肪酸甲酯混标 美国Sigma公司;RNAiso Plus、PremixVersion 2.0(Loading dye mix)、PrimeScriptRT reagent Kit with gDNA Eraser(Perfect Real Time)、SYBRPremix Ex™ II、6×loading buffer、Marker DL2000 大连宝生物工程有限公司;RNase-free水 北京天根生物技术有限责任公司;焦碳酸二乙酯 美国Intrivogen公司;核酸染料 无锡百泰克公司。
1.2 仪器与设备
Scion气相色谱仪 天美(中国)科学仪器有限公司;UMC-5型斩切机 德国Stephan公司;Foss Analytical A/S型食品分析仪 福斯华(北京)科贸有限公司;RE-52AA旋转蒸发器 上海亚荣生化仪器厂;HJ-6型多头磁力搅拌加热器江苏荣华仪器制造有限公司;ND-1000型微量紫外分光光度计基因有限公司;5810R离心机 德国Eppendorf公司;ZHJH-C1112C超净工作台 上海智城分析仪器制造有限公司;BG-power5000型稳压稳流电泳仪、水平电泳槽北京百晶生物技术有限公司;凝胶成像系统 美国Bio-Rad公司;LightCycler96实时聚合酶链式反应(real-time polymerase chain reaction,real-time PCR)仪 罗氏诊断产品(上海)有限公司;PCR扩增仪 美国Applied Biosystems公司。
1.3 方法
1.3.1 脂肪分布的测定
将羊胴体分割为尾脂、肌肉组织和胸腹部皮下组织并称质量;根据GB/T 9695.19—2008《肉与肉制品 取样方法》取样,并使用食品分析仪进行扫描分析,测得胸腹部皮下组织、背最长肌、臂三头肌和股二头肌脂肪含量。根据公式(1)~(5)计算:

1.3.2 脂肪酸组成的测定
参照王柏辉等的方法进行脂肪酸的提取和测定,结果为该脂肪酸峰面积与总脂肪酸峰面积之和的百分比(占比)。
1.3.3 重要基因网络表达量的测定
real-time PCR:参照袁倩等的方法,采用TRIzol法提取呼伦贝尔羊不同部位RNA,并使用紫外分光光度计检测总RNA的纯度和浓度,同时通过1%琼脂糖凝胶电泳检测RNA的污染和降解情况。根据TaKaRa公司的反转录试剂盒,进行反转录后cDNA置于-20 ℃保存备用。目的基因和管家基因的引物参照NCBI(http://www.ncbi.nlm.nih.gov/)中提供的序列进行设计(表1),由生工生物工程(上海)股份有限公司合成。按照SYBR Premix ExII试剂盒说明书操作。

表1 引物序列Table 1 Primer sequences used in this study
1.4 数据处理

2 结果与分析
2.1 巴尔虎羊和短尾羊的脂肪分布
脂肪分布是呼伦贝尔羊在生长发育过程中脂肪沉积的结果,而脂肪的沉积是贮存能量的主要方式,脂肪沉积的先后因机体部位而异,其肌内脂肪、皮下脂肪和尾部脂肪作为胴体主要脂肪对肉用品质(屠宰性能,食用品质和营养价值等)影响较大。由表2可知,短尾羊背最长肌脂肪含量、臂三头肌脂肪含量、肌内脂肪含量、肌内脂肪质量、胴体总脂肪质量和胴体脂率均显著大于巴尔虎羊(<0.05),且皮下脂肪含量和质量有此趋势但不显著(>0.05),说明短尾羊的体脂含量较高,主要体现在皮脂和肌内脂肪方面,皮脂是肉类加工的重要原料,而肉类的肌内脂肪含量可以通过影响其大理石花纹、风味等食用品质进而影响消费者的选择,故短尾羊肉用品质优势明显。值得注意的是,巴尔虎羊尾型较大这一性状并未在尾脂质量差异中体现,这与观察和报道的情况不符,分析原因可能是屠宰时正值冬季,处在高纬度地区半封闭式羊圈的巴尔虎羊消耗尾部脂肪以抵御严寒,这与赵伟利和王耀武的研究一致。另外,短尾羊较高的肌内脂肪和皮下脂肪也具备了越冬的条件,故气候对羊肉品质和经济效益具有深远的影响。

表2 巴尔虎羊和短尾羊的脂肪分布Table 2 Fat distribution in Baerhu sheep and short-fat-tailed sheep
2.2 巴尔虎羊和短尾羊的脂肪酸组成
2.2.1 肌肉组织
肉类脂肪中饱和脂肪酸以硬脂酸和棕榈酸居多,不饱和脂肪酸以油酸居多,然后是亚油酸。由表3可知,在肌肉组织中,巴尔虎羊背最长肌和股二头肌饱和脂肪酸占比显著大于短尾羊(<0.05),进一步研究发现巴尔虎羊背最长肌月桂酸和硬脂酸占比显著大于短尾羊(<0.05),同时短尾羊背最长肌多不饱和脂肪酸占比显著大于巴尔虎羊(<0.05),说明短尾羊肌肉组织中饱和脂肪酸占比较小,不饱和脂肪酸所占比例较大,相较于巴尔虎羊,短尾羊肉脂肪酸组成更为合理,更有益人体健康。值得注意的是,近年来反式脂肪酸受到广泛关注,其对人体健康的危害甚至大于饱和脂肪酸,来源多为部分氢化处理的植物油。巴尔虎背最长肌反式油酸和股二头肌反式亚油酸占比均显著大于短尾羊(<0.05),说明巴尔虎羊所占的反式脂肪酸比例较大。虽然有研究表明反式油酸能够体外抑制大鼠H9c2心肌细胞的增殖并诱导其凋亡,而反式亚油酸会诱导细胞氧化损伤对机体产生不良的影响,但是天然来源的反式脂肪酸对机体健康的不良影响尚不明显,相关研究证据并不充分。

表3 巴尔虎羊和短尾羊肌肉组织中的脂肪酸组成Table 3 Fatty acid composition of lipids in muscle tissues of Baerhu sheep and short-fat-tailed sheep
肉类功能性脂肪酸多为不饱和脂肪酸,如亚油酸是风味物质的重要底物,对羊肉的风味形成起着重要作用,亚麻酸是长链脂肪酸合成的前体物质,并且具有广泛的生物学意义。花生四烯酸具有免疫调节、抗炎、抗氧化、降血脂、抗肿瘤和保护皮肤等多种生理功能。二十二碳五烯酸在保护神经、抗癌和抗炎等方面有着非常重要的意义。二十碳五烯酸、二十二碳六烯酸是两种功能性长链脂肪酸,具有抗氧化、抗衰老的作用,提高记忆力及思维能力,对记忆力减退、老年性痴呆有一定疗效。由表3可知,巴尔虎羊臂三头肌亚油酸占比显著大于短尾羊(<0.05),短尾羊背最长肌-亚麻酸、花生四烯酸、二十二碳五烯酸、二十碳五烯酸、二十二碳六烯酸,臂三头肌二十二碳六烯酸、二十二碳五烯酸和股二头肌花生四烯酸、二十二碳五烯酸、二十二碳六烯酸占比均显著高于巴尔虎羊(<0.05),说明整体上看,短尾羊肌肉组织-亚麻酸、花生四烯酸、二十二碳五烯酸、二十碳五烯酸和二十二碳六烯酸占比较高,这些功能性脂肪酸作为人类的重要营养素发挥着重要作用。
2.2.2 脂肪组织
呼伦贝尔羊尾部和皮下脂肪为蓄积脂肪,是一种逆境生存所必需的生物性状,可以储存多余的能量以抵抗恶劣环境,如饥饿和御寒状态下,脂肪酸氧化增加,产生酮体为其他组织供能。由表4可知,在脂肪组织中,短尾羊尾部脂肪饱和脂肪酸占比显著大于巴尔虎羊(<0.05),皮下脂肪也有此趋势,但并不显著(>0.05),进一步研究发现短尾羊尾部脂肪癸酸、月桂酸、肉豆蔻酸和棕榈酸,皮下脂肪癸酸和棕榈酸占比均显著大于巴尔虎羊(<0.05)。同时巴尔虎羊尾部脂肪单不饱和与多不饱和脂肪酸,皮下脂肪多不饱和脂肪酸占比显著大于短尾羊(<0.05),说明巴尔虎羊脂肪组织中饱和脂肪酸占比较小,不饱和脂肪酸所占比例较大,相较于短尾羊,巴尔虎羊脂肪组织脂肪酸组成较好。具体来说,巴尔虎羊尾部脂肪油酸、亚油酸、-亚麻酸和共轭亚油酸,皮下脂肪亚油酸和共轭亚油酸占比显著大于短尾羊(<0.05)。

表4 巴尔虎羊和短尾羊脂肪组织中的脂肪酸组成Table 4 Fatty acid composition of lipids in adipose tissues of Baerhu sheep and short-fat-tailed sheep
2.3 巴尔虎羊与短尾羊脂肪代谢重要基因网络
2.3.1 RNA质量分析
由图2可知,所提取的总RNA可见28S、18S和5S条带清晰明亮,说明RNA完整性好无降解,用微量紫外分光光度计测得/均在1.8~2.1之间,表明质量较好、无污染,可用于后续实验。

图2 总RNA琼脂糖凝胶电泳检测Fig.2 Agarose gel electrophoresis analysis of total RNA

2.3.2 脂肪代谢重要基因网络mRNA表达量
由图3可知,巴尔虎羊背最长肌和臂三头肌的和mRNA相对表达量显著高于短尾羊,皮下脂肪和mRNA相对表达量显著低于短尾羊(<0.05),巴尔虎羊尾部脂肪的mRNA相对表达量显著高于短尾羊,mRNA相对表达量显著低于短尾羊(<0.05),和mRNA在脂肪组织表达较高。mRNA在巴尔虎羊和短尾羊肌肉部位的相对表达量极低,在皮脂中表达量较高,而巴尔虎羊尾部脂肪mRNA相对表达量显著高于短尾羊(<0.05)。mRNA在巴尔虎羊和短尾羊脂肪组织中的相对表达量均高于肌肉组织,其中皮下脂肪mRNA相对表达量显著低于短尾羊(<0.05)。巴尔虎羊臂三头肌、股二头肌和尾部脂肪mRNA相对表达量显著高于短尾羊(<0.05),背最长肌、臂三头肌、股二头肌和皮下脂肪mRNA相对表达量显著低于短尾羊(<0.05),背最长肌、皮下脂肪和尾部脂肪mRNA相对表达量显著高于短尾羊(<0.05),且、和mRNA相对表达量在脂肪部位较高,尤其是皮下脂肪。

图3 巴尔虎羊和短尾羊重要基因网络mRNA的表达量Fig.3 mRNA expression of important gene networks in Baerhu sheep and short-fat-tailed sheep
2.3.3 脂肪分布和脂肪酸组成与脂肪代谢重要基因表达量的相关性分析
由表5可知,皮下脂肪质量与其基因表达量呈显著正相关(<0.05),与基因呈显著负相关(<0.05),说明基因可能正向调控脂肪沉积,基因反之。5 个部位脂肪含量/质量与其基因表达量均呈正相关,反之但不显著(>0.05),说明的表达可能对脂肪沉积有促进作用,与之相反。臂三头肌脂肪含量与其基因表达量呈显著负相关(<0.05),股二头肌脂肪含量与其基因表达量呈显著正相关(<0.05),背最长肌脂肪含量与其基因表达量呈显著负相关(<0.05),说明和基因可能负向调控脂肪沉积,基因可能正向调控脂肪沉积。

表5 脂肪分布与脂肪代谢重要基因表达量相关性分析Table 5 Correlational analysis between fat distribution and the expression of important genes related to lipid metabolism
由表6可知,基因表达量与癸酸、月桂酸、肉豆蔻酸和棕榈酸呈显著负相关(<0.05),说明基因可能下调饱和脂肪酸的组成比例。基因与多不饱和脂肪酸、油酸和二十碳五烯酸呈显著负相关(<0.05),基因表达量与反式油酸、油酸和亚油酸呈显著正相关(<0.05),说明基因可上调不饱和脂肪酸的组成比例,基因反之。基因表达量与油酸呈显著正相关(<0.05),与单不饱和脂肪酸正相关但不显著(>0.05),基因表达量与油酸和单不饱和脂肪酸均呈显著正相关(<0.05),说明和基因可能有助于单不饱和脂肪酸的沉积。
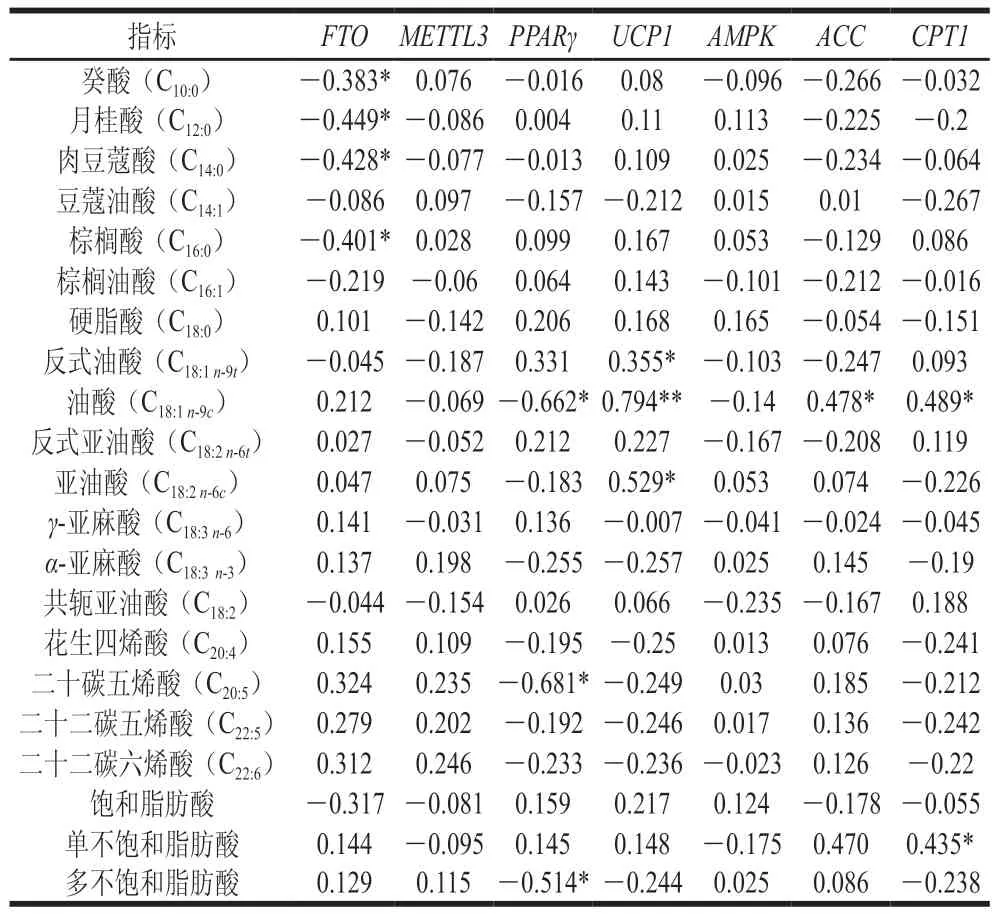
表6 脂肪酸组成与脂肪代谢重要基因表达量相关性分析Table 6 Correlation analysis between fatty acid proflie and expression of important genes related to lipid metabolism
3 讨论
3.1 脂肪代谢重要基因网络对脂肪分布的影响
mA甲基化修饰是高等生物mRNA上最为普遍的修饰,可甲基化mRNA RRACH序列的腺嘌呤,而作为去甲基化酶可逆转甲基化。基因在多种组织中均有表达,其高表达可促进脂肪沉积,而基因的组织表达特异性与类似,通过甲基化mA进而抑制动物脂肪沉积。本实验中,相较于短尾羊,巴尔虎羊肌肉组织中和基因表达量较高,皮下脂肪和基因表达量较低,短尾羊肌肉基因低表达与其高肌内脂肪含量不符,说明在肌肉组织中,尽管基因表达量与脂肪沉积呈正相关,但是需要考虑基因的存在,故肌内脂肪含量和皮下脂肪质量由和基因共同调控,这与Zhao Xu等的研究一致。巴尔虎羊尾部脂肪的基因表达量较高,短尾羊尾部脂肪基因表达量较低,而两个品系尾部脂肪质量并无差异,进一步说明在冬季巴尔虎羊尾部脂肪沉积小于消耗,这与其自身的生热作用密切相关。
生热作用对于温血动物至关重要,产热方式分为战栗性产热和非战栗性产热。战栗性产热是指骨骼肌发生不随意的节律性收缩产热,以维持体热平衡,而非战栗产热通过提高组织代谢率产热,常见于肩胛下区、颈部和腹股沟等处的棕色脂肪组织,其线粒体中可使葡萄糖和脂肪酸分解直接产热。具有脂肪组织特异性,不仅调控脂质代谢酶的表达,而且可以通过促进的表达进而影响机体的生热作用。本实验中,与短尾羊相比,巴尔虎羊皮下脂肪基因表达量较低,皮下脂肪质量也有类似显著差异,结合相关性分析,说明可促进皮下脂肪沉积,与侯海瑞的研究一致。巴尔虎羊和短尾羊肌肉组织基因表达量极低,而巴尔虎羊尾部脂肪基因表达量较高,说明相较于脂肪组织,肌肉组织中棕色脂肪含量较低,故推测肌肉组织御寒时多为战栗性产热,脂肪组织多为非战栗性产热。同时,巴尔虎尾部脂肪的高产热进一步说明其尾脂变小可能与其御寒有关。
可以通过调节动物能量代谢影响脂肪沉积,进而影响肉用品质。低温条件可激活,启动调控脂肪代谢的级联效应,磷酸化后迅速抑制活性,同时降低基因的表达从而减少脂肪酸合成,还可以减少丙二酰辅酶A的表达,减弱其对的抑制作用,故被激活后可促进线粒体中脂肪酸的-氧化,进而加速脂肪代谢产能。本实验中,与短尾羊相比,巴尔虎羊肌肉组织和尾部脂肪和基因表达量较高,基因表达量较低,说明巴尔虎羊肌肉和尾脂中信号通路较为活跃,脂肪酸的-氧化增加使其脂肪沉积减少,进而导致巴尔虎羊肌内脂肪和尾部脂肪减少。另外,Wang Xue等研究发现,呼伦贝尔羊基因正向调控皮下脂肪沉积,而本实验中基因在短尾羊皮下脂肪中表达较高,而表达较低,结合相关性分析,说明短尾羊皮下脂肪中脂肪酸的-氧化减少,促进脂肪沉积。
3.2 脂肪代谢重要基因网络对脂肪酸组成的影响
目前对于基因调控脂质代谢的研究多集中在脂肪合成、分解和转运方面,而对基因调控脂肪酸组成的研究较少,这可能是因为在大多情况下,化学结构的微小变化都可以显著改变脂质信号的活性,例如棕榈酸可以轻微抑制胰岛素介导的蛋白激酶磷酸化,而棕榈油酸对蛋白激酶磷酸化无抑制作用,另外,衍生自花生四烯酸骨架的脂蛋白会激活生热作用,而花生四烯酸本身会抑制线粒体活性。Chalupová等研究认为与基因对脂肪酸组成的影响尚未体现,而在本实验中,经相关性分析,基因下调饱和脂肪酸的组成比例,这与Li Zhenjie的研究一致。
众所周知,与其配体—长链多不饱和脂肪酸共同调控脂肪沉积,Sato等研究表明,的激活降低了单不饱和脂肪酸含量,从而抑制了大多数C或C多不饱和脂肪酸的还原。基因可以调控脂肪酸代谢,而脂肪酸也会对基因产生影响,如油酸和花生四烯酸会增加的表达,亚油酸会激活脂肪酸和葡萄糖吸收到棕色脂肪和肌肉,降低甘油三酯含量,提高耐寒性,促进脂肪组织产热。本实验中,结合脂肪酸组成及相关性分析,说明巴尔虎羊皮脂可以降低单不饱和脂肪酸和多不饱和脂肪酸占比,影响其脂肪酸组成,而尾部脂肪产热可能与其油酸和亚油酸有关。
可通过脂肪酸转位酶调控细胞摄入脂肪酸,尚未发现其对脂肪酸组成影响的研究,而其下游基因和与脂肪酸组成关系密切,如Rubio-Martin等发现基因高表达与饱和脂肪酸和脂肪沉积有关。杨蕾等研究发现基因高表达量有助于单不饱和脂肪酸的沉积,与本研究结果一致。另有研究表明,苏尼特羊尾脂基因表达量与亚油酸含量呈正相关。Ahn和Chang等研究发现,油酸可诱导基因上调,与本研究结果一致,说明和基因差异化表达对脂肪酸组成有重要的影响。
4 结论
即使同属呼伦贝尔羊,脂肪代谢重要基因网络(、、、、、和)表达的差异也最终造成了巴尔虎羊和短尾羊脂肪分布与脂肪酸组成的差异。从脂肪分布上看,巴尔虎羊胴体脂肪含量低于短尾羊,主要体现在高水平的肌内脂肪含量和皮下脂肪质量,同时季节对脂肪分布的具有重要影响,肉羊在抵御严寒时消耗了大量的脂肪,这有可能造成其产肉性能、食用品质、营养价值和饲料报酬的降低;从脂肪酸组成看,短尾羊肌肉组织脂肪酸的组成比例较好,主要体现在不饱和脂肪酸占比较高,不饱和脂肪酸中,多不饱和脂肪酸占比较高,多不饱和脂肪酸中,-亚麻酸、花生四烯酸、二十二碳五烯酸、二十碳五烯酸和二十二碳六烯酸占比较高;巴尔虎羊脂肪组织脂肪酸的组成比例较好,主要体现在不饱和脂肪酸所占比例较大,如亚油酸和共轭亚油酸等。巴尔虎羊和短尾羊脂肪分布与脂肪酸组成的不同可能最终导致了其屠宰性能、食用品质和营养价值的差异。值得注意的是,巴尔虎羊与短尾羊虽然同属呼伦贝尔羊,但是二者生热体系存在差异,在冬季枯草期巴尔虎羊尾部基因高表达量可能致使其脂肪减少,而短尾羊的高体脂含量也从另一角度解释了其御寒能力不只依靠于尾部,这为减少尾脂含量、提高肉羊生产效率提供了一定的理论依据。
