80份陆稻晚稻农家种资源的遗传多样性综合分析
2022-06-02徐志军李亚波欧阳红军徐磊安东升刘洋
徐志军 李亚波 欧阳红军 徐磊 安东升 刘洋
摘 要:为保护和利用陆稻农家种资源,本研究利用SNP標记和表型鉴定对80份陆稻晚稻农家种资源进行遗传多样性综合分析。结果表明:利用水稻1KSNP芯片对80份资源进行检测后共获得739个SNP的基因型数据,基于SNP的群体结构分析、PCA分析和聚类分析,均将80份资源分为云南亚群(63份)、海南亚群(11份)和贵州亚群(6份)共3个亚群,3个亚群间的平均遗传距离分别为0.83、0.63、0.80,其中海南亚群与其他2个亚群的遗传距离较远。贵州的陆稻资源分别属于贵州亚群和云南亚群,表明黔南地区的陆稻资源可能存在不同的来源。云南亚群可进一步分为3个亚群,平均遗传距离分别为0.30、0.28和0.22,表明细分亚群间的遗传距离很近。表型鉴定结果表明,11个表型性状的变异系数在4.09%~38.77%之间,其中有效分蘖数和穗重的变异系数较大,均超过25.00%,株高、种子长、种子宽、种子长宽比和种子圆度均值变异系数较小,均低于10%;遗传多样性指数范围为1.76~2.84,种子长的遗传多样性最丰富。相关分析结果表明,11个表型性状间存在着复杂的相关关系,其中种子长宽比与圆度均值的相关系数最高为0.98。在表型PCA分析中,前5个因子累计贡献率为82.37%,可反映表型性状的绝大部分信息,各成分的贡献率分别为29.08%、23.42%、12.35%、9.77%和7.75%,主要反映了株型和产量相关信息。80份资源的综合得分范围为0.15~0.73,排名前10的种质资源(ZRG36、ZRG101、ZRG58、ZRG31、ZRG1、ZRG102、ZRG104、ZRG63、ZRG29、ZRG11)均来自海拔较高地区,其中2份来源于海南,5份来源于贵州,3份来源于云南,且在穗长、种子长、种子宽、穗重和千粒重表现较好。该研究结果为陆稻农家种资源的保护和利用提供了参考。
关键词:陆稻农家种;SNP;表型;遗传多样性;综合评价中图分类号:S511.6 文献标识码:A
Comprehensive Genetic Diversity Analysis of Eighty Late Upland Rice Landrace
Abstract: The comprehensive genetic diversity of 80 upland rice landrace was evaluated based on SNP genotyping and phenotype identification, aiming to protect and utilize the upland rice landrace. The results showed that a total of 739 genotyping SNP data of the 80 landrace was obtained using a 1K rice SNP array. Based on the SNP genotyping data, the upland rice landrace was divided into Yunnan group, Hainan group and Guizhou group, through population structure analysis, PCA analysis and phylogenetic analysis. And there were 63, 11 and 6 landrace in the group, respectively. The genetic distance of the three groups was 0.83, 0.63 and 0.80, and the genetic distance between Hainan group and the other two groups was far relatively. The upland landrace, collected from Guizhou, were clustered into Guizhou group and Yunnan group respectively, indicating that there may be different sources of upland landrace in southern Guizhou. The coefficient variation of 11 agronomic and yield related traits ranged from 4.09% to 38.77%, and the coefficient variation of panicle weight and effective tiller was larger than 25.00%, while the coefficient variation of plant height, grain length, grain width, grain length/width ratio and mean grain roundness was smaller than 10%. The traits diversity index ranged from 1.76 to 2.94, and seed length had the greatest genetic diversity among the traits. Correlation analysis revealed complex correlation ship between the traits, and the correlation coefficient between grain length/width ration and mean grain roundness was 0.98. The phenotype PCA analysis showed that the first five principal components explained 29.08%, 23.42%, 12.35%, 9.77% and 7.75% phenotypic variation respectively. And the first five principal components totally explained 82.37% of the phenotypic variation, reflecting most information of the 11 traits. The comprehensive score of the landrace, extremely significant correlated with plant height, panicle length, flag leaf length, flag leaf width and effective tiller, was ranged from 0.15 to 0.73. The landrace ZRG36, ZRG101, ZRG58, ZRG31, ZRG1, ZRG102, ZRG104, ZRG63, ZRG29 and ZRG11 had the top comprehensive score all from higher elevations,. And the landrace, two from center region of Hainan Province, five from southern of Guizhou Province and three from Southwest and southeast of Yunnan Province, performed well in terms of panicle length, grain length, grain width, panicle weight and 1000-grain weight. The landrace were potential parent material for upland rice breeding or intermediate material for rice drought resistance breeding. The result will provide references for protecting and utilizing the upland rice landrace.AF2D856E-8F0A-401B-9F4A-53406B05FF5A
Keywords: upland rice landrace; SNP; phenotype; genetic variation; comprehensive evaluation
DOI: 10.3969/j.issn.1000-2561.2022.05.007
陆稻,也称为旱稻,是栽培稻(Oryza sativa)的旱生类型,具有耐旱、耐贫瘠和抗逆性好的特点,是抗旱、氮高效栽培稻遗传改良的宝贵资源。在全球气温升高、极端天气频繁和水资源短缺的背景下,我国稻作产业面临的环境风险不断提高,因干旱或季节性干旱造成的水稻减产面积和范围不断扩大[1]。因此,培育产量高、品质优、节水抗旱的水稻品种对于我国水稻产业发展具有重要意义[2]。云南、贵州、广西、海南等地是我国陆稻的传统种植区,分布有丰富的陆稻资源,这些资源是水稻抗旱、抗逆等功能基因的重要来源[3-5]。然而,随着高产水稻品种的培育和应用,传统的农家陆稻品系由于其产量水平較低导致种植面积急剧下降,陆稻农家种流失严重[1, 6]。此外,陆稻农家种普遍具有植株较高、生育期较长等特点,也限制对这些资源的利用[6]。表型是作物基因型和环境相互作用的综合表现,可直观反映作物的形态特征[3, 7];遗传多样性可反映不同来源种质资源的遗传背景差异及变异水平[8-9]。对作物种质资源开展表型和遗传多样性评价,可以更全面地了解资源的特性,为育种利用提供重要参考依据。本课题组前期收集陆稻资源时,发现有80份陆稻晚稻农家种资源。基于此,本研究拟利用SNP标记对从云南、贵州和海南等地收集的80份陆稻晚稻农家种资源的基因型进行检测,结合农艺性状和产量性状的鉴定,分析陆稻晚稻农家种资源的遗传多样性特点,明确陆稻晚稻农家种遗传的特性和亲缘关系,为陆稻晚稻农家种的保护和利用提供参考。
1 材料与方法
1.1材料
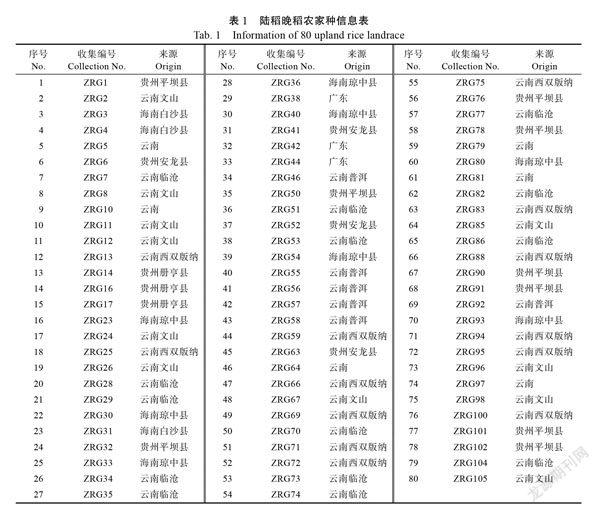
以本课题组收集的来源于云南、贵州、海南和广东的80份陆稻晚稻农家种资源为供试材料(表1),上述材料在广东湛江只能在6—11月种植(晚稻),春季种植均不能完成整个生育期。云南的陆稻晚稻资源主要来源于海拔1000~1400 m的滇东南(文山)和滇西南(临沧、西双版纳和普洱),贵州的陆稻晚稻资源来源于平均海拔1000 m的黔南地区(平坝县、册亨县和安龙县),海南的陆稻晚稻资源来源于平均海拔1000 m的海南中部地区(琼中县和白沙县)。
1.2 方法
1.2.1 SNP检测 使用博瑞迪生物技术有限公司的水稻1KSNP芯片对80份资源进行检测。
1.2.2 表型鉴定 供试材料于2019—2020年连续两年在广东省湛江市中国热带农业科学院湛江实验站基地种植,每份材料种植3个小区,每小区
种5行,每行种10株,株距13.3 cm,行距25 cm,收获前调查各材料的株高、穗长、剑叶长、剑叶宽、有效分蘖数共5个性状,收获后调查穗重、千粒种、种子长、种子宽、种子长宽比、种子圆度共6个性状。所有性状调查按照《水稻种质资源描述规范和数据标准》执行[10]。
1.3数据处理
1.3.1 SNP数据分析 使用Plink(http://www. cog-genomics.org/plink2/)软件对获得的SNP芯片clean数据进行质控,去除缺失≥0.05、最小等位基因频率(MAF)<0.05和=1的SNP位点。
1.3.2 群体结构分析 使用fastSTRUCTURE软件对群体结构进行分析,确定最佳分群[11];使用R软件包进行PCA分析,并绘制群体结构图和PCA图;使用MEGA 7.0软件构建进化树,并计算遗传距离(P-distance)[12],同时使用iTol工具(http://itol2.embl.de/)对进化树进行美化。
1.3.3 多样性分析 参照文献[3, 13-14]中的方法对性状划分等级,即M±kσ(其中k=0、0.5、1.0、1.5、2.0,M为平均值,σ为标准差),将这些性状表型值划分10个等级,每1级中材料所占的比例(Pi)用于计算多样性指数,遗传多样性指数即Shannon-Wiener index(H′)信息指数,其计算公式为:H′=-ΣPi×lnPi,式中Pi为某性状第i级中包含的材料份数所占材料总份数的百分比。
1.3.4 综合评价 使用模糊隶属函数法将80份陆稻晚稻农家种的11个表型性状函数值进行标准化[13];参照综合评价方法[15],使用R软件包对11个表型性状进行PCA分析,计算主成分因子得分系数矩阵,结合权重系数来计算综合评价F值。
2 结果与分析
AF2D856E-8F0A-401B-9F4A-53406B05FF5A2.1 陆稻晚稻农家种资源的群体结构分析
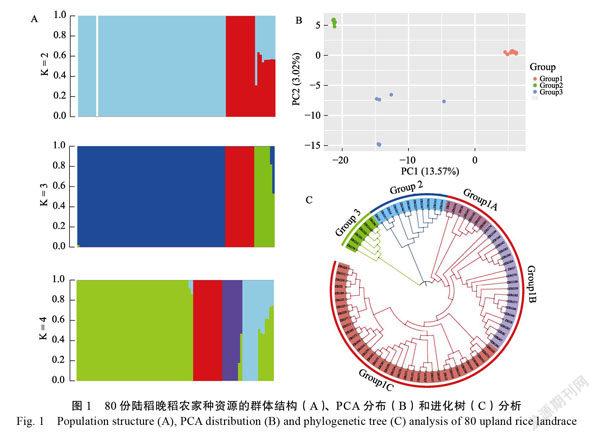
对80份陆稻晚稻农家种资源进行水稻1KSNP芯片检测后,共获得了1107个SNP位点,按照缺失和MAF进行过滤后,共获得739个符合要求的SNP位点。SNP的主基因频率范围为0.55~0.99,杂合度范围为0~0.16,PIC值范围为0~0.37。使用fastSTRUCTURE软件对群体结构进行分析和最佳分组筛选,在K=3时,群体有最佳的分组(图1A)。根据各品种的Q值,按照Q>0.5确定各品种的亚群,研究发现这些资源的遗传背景比较清晰,不同亚群间的基因交流较少。按照地域可将这些资源分为云南亚群、海南亚群和贵州亚群。其中云南亚群共有63份资源,主要以云南的资源为主,包含10份来源于贵州和3份来源于广东的陆稻晚稻农家种资源;海南亚群包含11份海南的陆稻晚稻农家种资源;贵州亚群包含6份来自于贵州的陆稻晚稻农家种资源。分属于不同亚群的贵州陆稻晚稻农家种资源,其Q值差异较大。
2.2 陆稻晚稻农家种资源的PCA分析和系统进化分析
PCA分析结果表明,80份陆稻晚稻农家种资源可分为云南亚群(Group1)、海南亚群(Group2)和贵州亚群(Group3)3个亚群(图1B)。
进化树分析结果表明,80份陆稻晚稻农家种资源可聚为云南亚群(Group1)、海南亚群(Group2)和贵州亚群(Group3)3类,与PCA和群体结构的结果相一致,表明K=3为该陆稻晚稻资源的最佳分群数(图1C)。3个亚群间的平均遗传距离分别为0.83、0.63、0.80,表明云南亚群和贵州亚群的遗传距离较近,海南亚群与其他两个亚群的遗传距离均较远。从图1可看出,3个亚群处于同一水平,可进一步将Group1所代表的云南亚群细分为3个亚群,其中Group1A为7份从贵州收集的陆稻晚稻农家种资源,这些资源与Group3亚群中来自于同一地区的资源遗传距离较远,而与来源于云南地区的资源遗传距离较近,表明黔南地区的陆稻晚稻资源可能存在着不同的来源;Gruop1B和Gruop1C分别包含20份和36份陆稻晚稻农家种资源,主要为云南西南地区和东南地区收集的陆稻晚稻农家种资源,这2个亚群中均包含来自广东的资源,Gruop1B中的2份廣东资源与云南西双版纳地区的2份资源聚在同一个分支,Gruop1C中的1份广东资源与云
南临沧的资源聚在同一个分支;3个细分亚群中均包含滇西南和滇东南地区的资源,西双版纳、临沧、普洱和文山的资源均不能按地区分开,细分亚群两两的遗传距离为0.30、0.28和0.22,表明云南亚群细分亚群间的遗传距离很近,其中从贵州收集的Group1A与Gruop1B、Gruop1C的遗传距离相对较远。
2.3陆稻晚稻农家种资源的表型变异和多样性分析
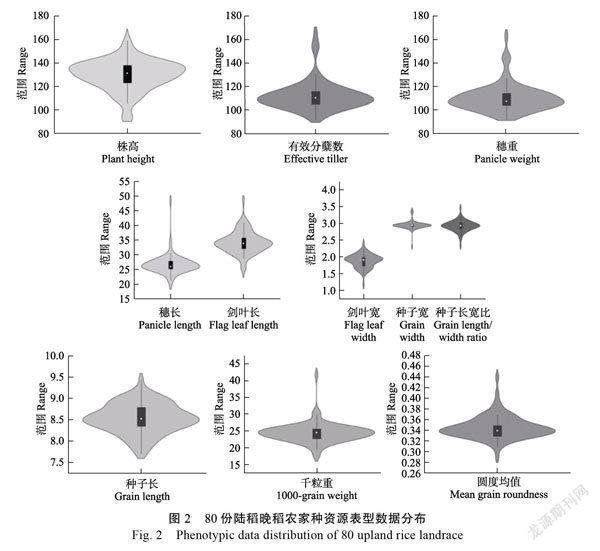
对80份陆稻晚稻农家种资源连续2年进行表型鉴定,结果表明,11个性状的表型分布和变异差异明显(图2和表2)。11个性状的变异系数为4.09%(种子长)~38.77%(穗重),其中有效分蘖数和穗重的变异系数较大,均超过25.00%;株高、种子长、种子宽、种子长宽比和种子圆度均值变异系数较小,均低于10%。11个性状的遗传多样性指数范围为1.76(剑叶宽)~2.94(种子长),遗传多样性丰富,其中种子长(2.94)和株高(2.85)的多样性指数最高。
2.4 陆稻晚稻农家种资源的表型相关分析
对陆稻晚稻农家种11个性状进行相关分析,结果见表3,株高与穗长、剑叶长和剑叶宽呈极显著正相关,穗长与剑叶长和剑叶宽呈极显著正相关。种子长和种子长宽比呈极显著正相关,与圆度均值呈极显著负相关;种子宽与种子长宽比呈极显著负相关,与圆度均值和穗重呈极显著正相关;种子长宽比与圆度均值和穗重呈极显著负相关。株高与有效分蘖数、剑叶长和剑叶宽、种子长和千粒重均呈显著正相关,穗长与千粒重呈显著负相关。其中种子长宽比与圆度均值的相关系数最高为0.98。
2.5 陆稻晚稻农家种资源的表型综合评价
对11个性状进行PCA分析,前5个主成分的累计贡献率达82.37%,可以解释表型性状的绝大部分信息(表4)。第一主成分的特征值最大,为3.19,贡献率为29.08%,其中种子长、宽、长
宽比和圆度均值的特征向量绝对值较大,主要反映产量相关信息;第二主成分的贡献率为23.42%,其中株高、穗长、剑叶长和剑叶宽的特征向量较大,说明该成分主要反映株型相关的信息;第三主成分的贡献率为12.35%,其中种子长、宽、千粒重、穗重的特征值较大,表明该特征值主要反映产量相关信息;第四主成分中有效分蘖数特征值最大为0.82,有效分蘖既与株型相关,又与产量相关,说明该成分主要反映株型和产量相关性状;第五主成分中千粒重的特征值最大,其次为穗重和种子长,表明该成分主要反映产量相关性状。
根据PCA分析获得了5个因子得分公式:
F1=0.04X1+0.14X2+0.19X3+0.06X4+0.11X5–0.34X6+ 0.39X7–0.53X8+0.54X9+0.27X10–0.10X11AF2D856E-8F0A-401B-9F4A-53406B05FF5A
F2=0.52X1+0.45X2+0.45X3+0.50X4+0.05X5+0.05X6–0.11X7+0.12X8–0.10X9–0.13X10–0.14X11
F3=0.11X1+0.02X2+0.12X3–0.01X4+0.28X5+0.55X6+0.42X7+0.08X8–0.05X9+0.34X10+0.55X11
F4=0.17X1+0.04X2–0.22X3–0.18X4+0.82X5–0.18X6–0.29X7+0.10X8–0.06X9+0.24X10–0.18X11
F5=0.16X1–0.14X2+0.24X3–0.14X4+0.09X5–0.41X6–0.27X7–0.08X8+0.10X9–0.33X10+0.71X11
将F1~F5的得分,根据各成分的权重(0.35、0.28、0.15、0.12、0.09),计算出各资源的综合得分F,80份资源的得分范围为0.15(ZRG7)~0.73(ZRG36),平均值为0.44(表5)。F值与11个性状的相关分析表明,F值与株高、穗长、剑叶长、剑叶宽和有效分蘖数呈极显著正相关,而与
其他性狀的相关关系不显著(表6)。综合排名前10位的农家种为ZRG36、ZRG101、ZRG58、ZRG31、ZRG1、ZRG102、ZRG104、ZRG63、ZRG29、ZRG11,这些资源的穗长、种子长、种子宽、穗重和千粒重表型较好,且均分布于海拔较高地区,其中ZRG31和ZRG36来源于海南中部地区,ZRG1、ZRG63、ZRG101、ZRG102 和ZRG104来源于贵州黔南,ZRG11、ZRG29和ZRG58来源于滇西南和东南地区,是潜在的陆稻晚稻育种亲本材料或水稻抗旱育种的中间材料。
3 讨论
我国是栽培稻的起源地之一,稻作历史悠久,稻种资源丰富。陆稻是栽培稻的旱作生态类型,是在自然环境与人工驯化共同作用下形成的一类具有耐旱、耐贫瘠等特性的特殊种质资源,是水稻育种中最直接、最有效的抗旱基因供体来源[16-18]。云南、海南和贵州具有丰富的陆稻资源[3, 19],本研究利用739个SNP标记将从这些地区收集的80份陆稻晚稻资源按照来源划分为云南亚群、海南亚群和贵州亚群,其中海南亚群与其他2个亚群的亲缘关系均较远,可能与海南岛与大陆隔离,与其他地区的资源交流较少有关;云南亚群中包含部分贵州收集的晚稻资源,这些晚稻资源与云南的晚稻资源遗传距离较近,而与同样来源于贵州黔南的贵州亚群的晚稻资源遗传距离较远,推测云南和贵州地区可能发生了晚稻农家种的交流,且贵州地区的晚稻农家种可能有不同的来源和驯化;此外,在云南亚群中,还存在3份来源于广东的晚稻农家种资源,分别与云南临沧和西双版纳的晚稻资源遗传距离较近,而与海南地区的晚稻资源遗传距离较远,由于这3份资源(从国家水稻中期库引进)具体来源地不详,且收集的资源中广东地区的晚稻资源较少,初步推测可能与晚稻农家种的引进交流有关,还需进一步结合本地区其他陆稻晚稻资源进行综合分析。前人研究发现,云南滇西南与滇东南的的陆稻品种之间的差异显著[6],而本研中鉴定的滇东南和滇西南的陆稻晚稻资源差异并不明显,可能与所鉴定的晚稻资源类型较为单一有关。本研究结果还表明,80份陆稻晚稻资源的11个表型性状的表型分布和变异差异明显,遗传多样性指数范围为1.76~2.85,11个性状间形成了复杂的相关关系,80个品种的得分范围为0.15~0.73,这些结果对陆稻晚稻农家种资源的利用和保护具有参考价值。AF2D856E-8F0A-401B-9F4A-53406B05FF5A
遺传多样性评价是认识作物基因型和表型多样性程度,以及挖掘和利用资源中优异基因资源的理论和实践基础[20-21]。表型多样性有助于从整体上认识作物性状的多样性程度,为优异资源的挖掘和利用提供理论和实践依据。本课题组在前期鉴定中发现,收集的陆稻资源中,有80份来源于云南、贵州、广东和海南的农家种晚稻资源,这些资源只有晚稻种植才能完成整个生育期。本研究对这80份陆稻晚稻资源进行表型鉴定,发现所调查的11个性状表现出丰富的表型变异。从地理分布来看,在海南和贵州收集的陆稻晚稻资源均分布在海拔约为1000 m的地区;在云南滇西南和滇东南地区收集的陆稻晚稻资源,主要分布在海拔1000~1400 m的半山区[6]。从气候条件看,海南中部地区属于热带季风气候,日照充足,陆稻晚稻种植期间平均温度在26℃左右;黔南地区属于亚热带湿润季风气候,阴雨天多,日照少;云南滇南陆稻资源分布地区气候与贵州类似,为半山云雾多湿区,日照不充分,陆稻晚稻生长期平均温度在23℃左右。相似的地理和生态环境促进了云南和贵州地区的资源引种和交流,可能是造成贵州黔南地区陆稻晚稻资源遗传差异较大的原因。这些地理和气候环境因素,是陆稻农家种驯化的重要动力,造成了陆稻资源丰富的表型多样性,也从生态方面解释了为何海南的资源与贵州、云南的资源遗传距离较远。
分子标记是开展遗传多样性研究的重要工具,在陆稻研究中,RFLP、RAPD、SSR、SRAP等多种分子标记已经用于分析资源的遗传背景、亲缘关系和基因多态性[17-18, 22-23],如王英等[22]利用22条多态性RAPD引物分析了49分陆稻的遗传基础,并构建了指纹图谱;张立娜等[17]利用39对SSR引物分析了158份不同来源的陆稻资源的遗传相似性,发现籼型旱稻地方品种的基因多样性显著高于粳型旱稻地方品种;徐建欣等[6]利用24对SSR引物分析了131份云南陆稻品种的遗传多样性,引物的等位基因数平均为8.125个,基因多样性指数平均为0.6543,研究发现滇西南和滇南地区存在丰富的遗传变异。而本研究利用739个SNP分析了80份陆稻晚稻农家种资源的亲缘关系,经群体结构分析、PCA分析和聚类分析,均将80份陆稻晚稻农家种资源分成了云南、海南和贵州3个亚群,说明了种群划分的准确性。SNP标记与传统的SSR和RAPD等标记相比,由于只涉及单个碱基的变异,其检测到的等位基因数一般是2~3个,但基于大量水稻资源测序构建的水稻SNP芯片,能够更均匀的检测基因组上的变异,可以更加准确对资源遗传背景和亲缘关系进行判定。随着芯片技术的成熟和测序成本的降低,基于基因芯片的SNP标记展现出高通量、高效、快速的特点,已经越来越多用于水稻[24]、玉米[25]、棉花[26]、油菜[27]等多种作物的遗传多样性研究和指纹图谱构建,将更加高效的用于种质资源评价和品种鉴定,促进优异资源的挖掘和利用。
参考文献
[1] 殷会德, 米铁柱, 刘佳音, 于 萌, 顾晓振, 郭海鹏, 李继明. 早稻研究现状及发展前景[J]. 中国稻米, 2020(4): 22-24.YIN H D, MI T Z, LIU J Y, YU M, GU X Z, GUO H P, LI J M. Research status and development prospects of upland rice[J]. China Rice, 2020(4): 22-24. (in Chinese)
[2] 罗利军, 梅捍卫, 余新桥, 刘鸿艳, 冯芳君. 节水抗旱稻及其发展策略[J]. 科学通报, 2011, 56(11): 804-811.LUO L J, MEI H W, YU X Q, LIU H Y, FENG F J. Water-saving and drought-resistance rice and its development strategy[J]. Chinese Science Bulletin, 2011, 56(11): 804-811. (in Chinese)
[3] 宫彦龙, 雷 月, 闫志强, 刘雪薇, 张大双, 吴健强, 朱速松. 不同生态区粳稻资源表型遗传多样性综合评价[J]. 作物杂志, 2020(5): 71-79.GONG Y L, LEI Y, YAN Z Q, LIU X W, ZHANG D S, WU J Q, ZHU S S. Comprehensive evaluation of phenotype genetic diversity in japonica rice germplasm resources in different ecological zones[J]. Crops, 2020(5): 71-79. (in Chinese)
[4] 王一平. 旱稻种质资源遗传多样性研究[D]. 北京: 中国农业科学院, 2006.WANG Y P. Study on genetic diversity of upland rice resources[D]. Beijing: Chinese Academy of Agricultural Sciences, 2006. (in Chinese)
[5] 张立娜. 中国水旱稻地方品种遗传多样性分析[D]. 北京: 中国农业科学院, 2010.ZHANG L N. Genetic diversity of lowland and upland rice resources in China[D]. Beijing: Chinese Academy of Agricultural Sciences, 2010. (in Chinese)AF2D856E-8F0A-401B-9F4A-53406B05FF5A
[6] 徐建欣, 王云月, 姚 春, 刘云霞, 汤 淼, 陆明兴. 利用SSR分子标记分析云南陆稻品种遗传多样性[J]. 中国水稻科学, 2012, 26(2): 155-164.XU J X, WANG Y Y, YAO C, LIU Y X, TANG M, LU M X. Genetic diversity of upland rice varieties in Yunnan province revealed by SSR markers[J]. Chinese Journal of Rice Science, 2012, 26(2): 155-164. (in Chinese)
[7] 都真真, 李锡香, 宋江萍, 武亚红, 赵 青, 徐 婷, 张晓辉, HELLIER B, HU J G, 王海平. 228份引進大蒜资源的表型多样性分析及适应性初步评价[J]. 植物遗传资源学报, 2019, 20(5): 1186-1196.DU Z Z, LI X X, SONG J P, WU Y H, ZHAO Q, XU T, ZHANG X H, HELLIER B, HU J G, WANG H P. Phenotypic diversity and adaptability analysis of 228 accessions of introduced garlic genetic resources[J]. Journal of Plant Genetic Resources, 2019, 20(5): 1186-1196. (in Chinese)
[8] 万映伶, 刘爱青, 张孔英, 刘 燕. 菏泽和洛阳芍药品种资源表型多样性研究[J]. 北京林业大学学报, 2018, 40(3): 110-121.WAN Y L, LIU A Q, ZANG K Y, LIU Y. Phenotype diversity of herbaceous peony variety resources in Heze, Shandong of eastern China and Luoyang, Henan of central China[J]. Journal of Bjing Forestry University, 2018, 40(3): 110-121. (in Chinese)
[9] 陈 越, 张敦宇, 丁明亮, 王玲仙, 肖素勤, 柯 学, 程在全. 多个省份水稻资源的表型多样性与优异资源的筛选[J]. 浙江农业学报, 2019, 31(11): 1779-1789.CHEN Y, ZHANG D Y, DING M L, WANG L X, XIAO S Q, KE X, CHENH Z Q. Phenotypic diversity of rice resources in multiple provinces and screening of excellent resources[J]. Acta Agriculturae Zhejiangensis, 2019, 31(11): 1779-1789. (in Chinese)
[10] 韩龙植. 水稻种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2006.HAN L Z. Descriptors and data standard for rice (Oryzaspp.)[M]. Beijing: China Agriculture Press, 2006. (in Chinese)
[11] RAJ A, STEPHENS M, PRITCHARD J K. fastSTRUCTURE: variational inference of population structure in large SNP data sets[J]. Genetics, 2014, 197(2): 573-589.
[12] KUMAR S, STECHER G, TAMURA K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution, 2016, 33(7): 1870-1874.
[13] 徐志军, 吴小丽, 胡小文, 刘 洋. 33份引进花生资源表型遗传多样性分析及在粤西地区的适应性初步评价[J]. 热带作物学报, 2021, 42(7): 1885-1895.XU Z J, WU X L, HU X W, LIU Y. Phenotypic diversity of 33 introduced peanut germplasm accessions and preliminary adaptability evaluation in West Guangdong[J]. Chinese Journal of Tropical Crops, 2021, 42(7): 1885-1895. (in Chinese)
[14] 孙珍珠, 李秋月, 王小柯, 赵婉彤, 薛 杨, 冯锦英, 刘小丰, 刘梦雨, 江 东. 宽皮柑橘种质资源表型多样性分析及综合评价[J]. 中国农业科学, 2017, 50(22): 4362-4372.SUN Z Z, LI Q Y, WANG X K, ZHAO W T, XUE Y, FENG J Y, LIU X F, LIU M Y, JIANG D. Comprehensive evaluation and phenotypic diversity analysis of germplasm resources in mandarin[J]. Scientia Agricultura Sinica, 2017, 50(22): 4362-4383. (in Chinese)AF2D856E-8F0A-401B-9F4A-53406B05FF5A
[15] 胡标林, 万 勇, 李 霞, 雷建国, 罗向东, 严文贵, 谢建坤. 水稻核心种质表型性状遗传多样性分析及综合评价[J]. 作物学报, 2012, 38(5): 829-839.HU B L, WAN Y, LI X, LEI J G, LUO X D, YAN W G, XIE J K. Analysis on genetic diversity of phenotypic traits in rice (Oryza sativa) core collection and its comprehensive assessment[J]. Acta Agronomica Sinica, 2012, 38(5): 829-839. (in Chinese)
[16] 王一平, 魏兴华, 华 蕾, 袁筱萍, 余汉勇, 徐 群, 汤圣祥. 不同地理来源旱稻种质资源的遗传多样性分析[J]. 作物学报, 2007, 33(12): 2034-2040.WANG Y P, WEI X H, HUA L, YUAN X P, YU H Y, XU Q, TANG S X. Genetic diversity in upland rice germplasm from different geographic regions[J]. Acta Agronomica Sinica, 2007, 33(12): 2034-2040. (in Chinese)
[17] 张立娜, 曹桂兰, 韩龙植. 中国不同地理来源旱稻地方品种的遗传相似性研究[J]. 中国农业科学, 2010, 43(17): 3481-3488.ZHANG L N, CAO G L, HAN L Z. Analysis of genetic similarity for upland landrace rices from different geographical origins in China[J]. Scientia Agricultura Sinica, 2010, 43(17): 3481-3488. (in Chinese)
[18] 杨学辉, 袁 洁, 陈小均, 阮仁超, 何海永, 吴石平, 王云月. 贵州旱稻种质资源的SSR遗传多样性分析[J]. 分子植物育种, 2009, 7(5): 890-896.YANG X H, YUAN J, CHEN X J, RUAN R C, HE H Y, WU S P, WANG Y Y. Genetic diversity of upland rice germplasm resources from Guizhou based on SSR markers[J]. Molecular Plant Breeding, 2009, 7(5): 890-896. (in Chinese)
[19] 游俊梅, 陈惠查, 金桃叶, 阮仁超. 贵州地方旱稻种质资源遗传多样性评价[J]. 种子, 2005(4): 79-81, 84.YOU J M, CHEN H C, JIN T Y, RUAN R C. Genetic diversity evaluation of upland rice germplasm resources in Guizhou province[J]. Seeds, 2005(4): 79-81, 84. (in Chinese)
[20] 陈 巍, 王力荣, 朱更瑞, 方伟超, 陈昌文, 曹 珂, 张绍铃. 基于SSR标记和生物学性状进行桃遗传多样性的比较分析[J]. 植物遗传资源学报, 2009, 10(1): 86-90, 96.CHEN W, WANG L R, ZHU G R, FANG W C, CHEN C W, CAO K, ZHANG S L. Comparison of genetic diversity among peach cultivars based on biological traits and SSR markers[J]. Journal of Plant Genetic Resources, 2009, 10(1): 86-90, 96. (in Chinese)
[21] CAMPBELL D R. Using phenotypic manipulations to study multivariate selection of floral trait associations[J]. Annals of Botany, 2009, 103(9): 1557-1566.
[22] 王 英, 葉 通, 邱海燕, 庄南生. 49份旱稻种质RAPD标记遗传多样性分析[J]. 中国农学通报, 2011, 27(24): 21-28.WANG Y, YE T, QIU H Y, ZHUANG N S. Analysis on genetic diversity of 49 upland rice germplasm using RAPD markers[J]. Chinese Agricultural Science Bulletin, 2011, 27(24): 21-28. (in Chinese)
[23] 姚宇飞, 王 英, 庄南生, 高和琼. 利用SRAP标记分析海南旱稻地方品种的遗传多样性及其分子身份证的构建[J]. 生物技术通报, 2014(11): 97-106. (in Chinese)YAO Y F , WANG Y, ZHUANG N S, GAO H Q. Analysis on genetic diversity of hainan upland rice local varietiesand establishment of molecular ID with SRAP markers[J]. Biotechnology Bulletin, 2014(11): 97-106. (in Chinese)AF2D856E-8F0A-401B-9F4A-53406B05FF5A
[24] 李梓榕, 袁 雄, 陈 叶, 郑兴飞, 胡中立, 李兰芝. 基于全基因组SNP高效鉴定水稻种质资源并构建指纹图谱[J]. 分子植物育种, 2020, 18(18): 6050-6057.LI Z R, YUAN X, CHEN Y, ZHENG X F, HU Z L, LI L Z. Effective identification for varieties by genome-wide SNPs and establishment of fingerprint for rice germplasms[J]. Molecular Plant Breeding, 2020, 18(18): 6050-6057. (in Chinese)
[25] 田紅丽, 杨 扬, 王 璐, 王 蕊, 易红梅, 许理文, 张云龙, 葛建镕, 王凤格, 赵久然. 兼容型maizeSNP384标记筛选与玉米杂交种DNA指纹图谱构建[J]. 作物学报, 2020, 46(7): 1006-1015.TIAN H L, YANG Y, WANG L, WANG R, YI H M, XU L W, ZHANG Y L, GE J R, WANG F G, ZHAO J R. Screening of compatible maizeSNP384 markers and the construction of DNA fingerprints of maize varieties[J]. Acta Agronomica Sinica, 2020, 46(7): 1006-1015. (in Chinese)
[26] 孙正文, 匡 猛, 马峙英, 王省芬. 利用CottonSNP63K芯片构建棉花品种的指纹图谱[J]. 中国农业科学, 2017, 50(24): 4692-4704.SUN Z W, KUANG M, MA Z Y, WANG X F. Construction of cotton variety fingerprints using CottonSNP63K array[J]. Scientia Agricultura Sinica, 2017, 50(24): 4692-4704. (in Chinese)
[27] 赵仁欣, 李森业, 郭瑞星, 曾新华, 文 静, 马朝芝, 沈金雄, 涂金星, 傅廷栋, 易 斌. 利用SNP芯片构建我国冬油菜参试品种DNA指纹图谱[J]. 作物学报, 2018, 44(7): 956-965.ZHAO R X, LI S Y, GUO R X, ZENG X H, WEN J, MA C Z, SHEN J X, TU J X, FU T D, YI B. Construction of DNA fingerprinting forBrassica napusvarieties based on SNP chip[J]. Acta Agronomica Sinica, 2018, 44(7): 956-965. (in Chinese)AF2D856E-8F0A-401B-9F4A-53406B05FF5A
