不同种植方式对连作土壤细菌群落结构和多样性的影响
2022-05-24宋秀丽黄瑞龙柯彩杰黄蔚章武陶波
宋秀丽,黄瑞龙,柯彩杰,黄蔚,章武*,陶波
1. 岭南师范学院,广东 湛江 524048;2. 东北农业大学,黑龙江 哈尔滨 150030
土壤微生物是土壤中生命活性物质,参与土壤中物质循环和能量代谢,对维持农田土壤肥力,保持土壤健康和农业可持续生产具有重要作用(宋长青等,2013;蔡元锋等,2014;潘孝晨等,2019)。基于16S rRNA基因的分子生物技术能够较全面和准确地反映土壤微生物群落结构和物种遗传多样性,目前已被广泛地应用在土壤微生物群落的研究上(Edgar,2013;夏围围等,2014)。
大豆(Glycine max)是中国重要粮食作物之一,也是东北黑土区旱地主要种植作物之一。由于土地资源有限、经济利益驱动及种植习性,大豆连作为当地主要种植方式。在连作过程中出现的连作障碍,增加植物的发病率,影响作物的健康,其中大豆根腐病和包囊线虫成为制约大豆生产的重要病害(Mo et al.,2016;Xiong et al.,2016;宋杰,2016)。选用大豆抗病品种,将大豆与其他作物进行轮作种植是目前防治病害的主要措施(Perez-Brandan et al.,2014;王芳等,2018)。不同的农业种植方式影响着土壤中的化学、物理和生物过程,并引发土壤微生物群落组成的变化(Fierer et al.,2012)。增加作物类型和轮作能够促进土壤养分的合理利用,协调土壤微生物的丰富度和均匀度,增加作物抗病性(Tian et al.,2015:Wang et al.,2015;Benitez et al.,2017)。也有研究表明,大豆长期连作可以减轻连作障碍。大豆连作14年,土壤中大豆根腐病潜在病原菌镰孢菌属的物种丰度显著降低(Hamid et al.,2017;Liu et al.,2019)。
土壤细菌是养分活化的驱动者,是土壤微生物中的重要群体,同时是一些植物病原物的拮抗菌(Mendes et al.,2011)。国内外学者对不同种植方式下大豆细菌群落多样性的研究表明,种植方式对土壤中细菌丰度和多样性都产生了影响,并且土壤中有效 N和有机质(OM)对微生物群落影响较大(Fan et al.,2011;周岚等,2013)。目前,关于作物轮作缓解连作障碍的研究报道较多(Tan et al.,2016:李锐等,2015),但在大豆长期连作的土壤上,不同种植方式对土壤细菌群落的影响尚不明确。因此本研究基于 Illumina MiSeq第二代高通量测序平台,设置休耕、休耕-大豆、玉米-大豆、小麦-大豆和大豆连作 5个处理,研究5种种植方式对土壤细菌群落结构组成、物种多样性的影响,并与土壤化学性质指标进行相关性分析,以期为优化大豆种植,改善土壤微生态提供理论依据。
1 材料与方法
1.1 试验地概况
本试验于 2005年开始,试验地位于中国北方黑龙江省黑河市嫩江中储粮公司四场第五中队。嫩江是中国大豆之乡,是麦豆主要生产区,土壤类型为黑土。试验地气候属于寒温带大陆性季风气候,地理坐标为 125°27′E,49°33′N。当地年平均气温为0.8—1.4 ℃,平均降雨量为480—512 mm,冬季寒冷、漫长、干燥,最低气温为-47.3— -43.7 ℃;夏季雨热同季,最高气温为33.9—37.4 ℃。
1.2 试验设计
2005年3月之前实验地为大豆连作种植区域,2005年4月开始设置5种种植方式处理:休耕地块(CK)、大豆连作种植(SC)、休耕-大豆轮作种植(FS)、玉米-大豆轮作种植(CS)和小麦-大豆轮作种植(WS),每个处理设3个重复小区,每个小区面积666.7 m2,采用完全随机区组设计。作物种植前使用尿素、磷酸二铵和硫酸钾3种肥料。大豆每年施用 33.1 kg·hm-2N、24.1 kg·hm-2P 和 21.6 kg·hm-2K,玉米施用 59.0 kg·hm-2N、40.1 kg·hm-2P 和 43.1 kg·hm-2K,以及小麦施用 73.0 kg·hm-2N、30.1 kg·hm-2P 和 21.6 kg·hm-2K。在玉米的孕穗阶段额外施用59.0 kg·hm-2N。
1.3 土壤取样
于2016年10月30日秋季大豆成熟后进行土壤样品采集。每个小区采用5点法在作物根部周围用土钻采耕层土(直径5 cm,深20 cm),装入密封袋,重复3次,混合为一个土壤样品,装入干净的无菌自封袋中。将共15份土壤样品运回实验室,剔除作物根系残体和杂物,取一份样品于-80 ℃冰箱中保存,用于土壤微生物分析,另一份样品用于土壤化学性质的测定。
1.4 土壤化学性质测定
土壤样本室温风干后磨碎,过0.2 mm筛。微量元素有效态铜(Cu)、铁(Fe)、锰(Mn)和锌(Zn)用0.005 mol·L-1二乙三胺乙酸提取后,用原子吸收分光光度计测定;有效态硼(B)用钾亚胺比色法测定;土壤pH按水土比2.5∶1进行前处理,然后用pH计测定;土壤有机质(OM)使用重铬酸钾容量法检测;有效磷(P)用0.5 mol·L-1NaHCO3浸提-钼锑抗比色法检测;有效氮(N)采用扩散法检测;有效钾(K)采用1 mol·L-1NH4OAc浸提-火焰光度法检测(鲁如坤,2000)。
1.5 土壤细菌总DNA的提取及16S rRNA扩增测序
使用土壤基因组DNA试剂盒(Power Soil™ DNA Isolation Kit,MoBio Laboratories Inc.,CA)提取 0.5 g土壤样品的总DNA,利用1.0%琼脂糖凝胶电泳和超微量紫外分光光度计检测DNA质量。以前端引物343F(5′-TACGGRAGGCAGCAG-3′)和后端引物798R(5′-AGGGTATCTAATCCT-3′)对 16S rRNA 基因的V4—V5区进行PCR扩增。PCR反应条件为:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s,25个循环;72 ℃延伸20 min。扩增结束后,使用2%琼脂糖凝胶对PCR扩增产物进行电泳,检查扩增效果。切取目的片段后,用Axygen凝胶回收试剂盒回收。将回收的目的片段进行测序。每个样本设置3个重复实验,将同一个样本的3个平行扩增产物混合得到一个样本的原始文库。采用Illumina Miseq二代高通量测序平台2×250 bp的双端测序策略对文库进行测序。土壤微生物测序服务委托上海天昊生物科技有限公司完成。
1.6 数据处理与分析
首先对原始测序序列进行过滤,得到有效序列,然后对有效序列进行去杂优化,采用UPARSE聚类方法将经过质控后的序列按序列间 97%的相似性进行聚类,形成不同的分类单元(operational taxonomic units,OTUs)。利用 mothur(Version 1.33.3)进行Alpha多样性分析(Chao、ACE等物种丰富度统计,Shannon、Simpson等物种多样性统计)、Venn图绘制,Beta多样性分析(PCA分析)。利用Excel软件对数据进行整理、汇总及相关系数等计算分析,利用SPSS 19.0软件中的Duncan分析法对数据进行单因素方差分析,P<0.05为差异显著。所有样品相似度树图、Venn图、PCA分析图、微生物群落组成结构等制图通过R语言软件的ggplot 2程序包完成。
2 结果与分析
2.1 种植方式对土壤化学性质的影响
由表1可知,不同种植方式对土壤化学性质产生了显著影响(P<0.05)。与CK相比,SC土壤中OM、有效N、K、Zn、Fe和Mn含量显著增加,有效P显著降低。FS土壤中有效Cu、Mn和P含量显著降低。WS土壤有效P和Cu含量显著增加,土壤中OM、有效N和Fe显著低。CS土壤中有效K和Zn含量显著降低。CK土壤中OM、有效K和Mn的含量显著高,有效Cu和Fe含量显著低。可见,不同种植方式显著改变了土壤化学性质。

表1 不同种植方式对土壤化学性质的影响Table 1 Soil chemical properties of different cropping systems
2.2 不同种植方式土壤细菌群落结构比较分析
对各样品细菌群落物种丰度及分布进行PCA,由图1a可知,主成分1和主成分2分别占51.807%和 17.979%,样本的细菌群落结构呈现显著差异(P<0.05)。5种种植方式下土壤细菌群落分成3组,其中休耕和轮作处理细菌群落显著不同于 SC处理,FS与CK细菌群落结构相似,WS和CS细菌群落结构相似,并且WS和CS细菌群落结构相似程度较高(图1b),说明不同轮作方式对土壤细菌群落结构影响显著。

图1 不同种植方式土壤细菌群落的相似度分析Figure 1 Differences of soil bacterial community structure in different cropping systems
2.3 不同种植方式对土壤细菌群落丰度和多样性的影响
如表2所示,不同种植方式下土壤细菌菌群丰度及菌群多样性差异显著(P<0.05)。FS土壤中细菌菌群丰度(Chao1/ACE)和菌群多样性(Shannon)显著增高(表1)(P<0.05),其他种植方式下细菌群落丰度和多样性差异不显著。菌群丰度总体表现为FS>CK>CS>WS>SC,菌群多样性总体表现为FS>WS>CS>CK>SC。与 SC 相比,FS、CS和 WS种植方式能够增加土壤中细菌菌群丰度和菌群多样性。
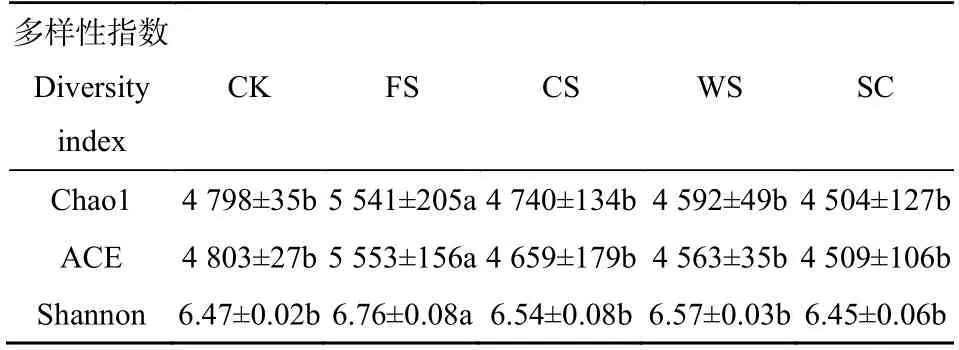
表2 不同种植方式土壤细菌群落的丰度及多样性Table 2 Effects of different cropping systems on bacterial community abundance and diversity
如图2所示,5种种植方式下土壤环境中共有细菌OTUs数量为2376个。CK土壤中细菌OTUs有4965个,其中特有OTUs有747个。FS土壤中细菌OTUs有5788个,其中特有OTUs有535个。CS土壤中细菌OTUs有4893个,其中特有OTUs有183个。WS土壤中细菌OTUs有4789个,其中特有OTUs有225个。SC土壤中细菌OTUs有4835个,其中特有OTUs有268个。可见,不同种植方式土壤环境中细菌OTUs呈现差异,并且不同种植方式改变了土壤特有OTUs的数量,进而影响了土壤细菌群落丰度和均匀度。

图2 不同种植方式土壤细菌OTUs的Venn图Figure 2 Venn diagram of soil bacteria OTUs under different cropping systems
2.4 不同种植方式对土壤细菌群落组成的影响
由图3可知,东北黑土土壤中细菌群落主要分布在变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)和放线菌门(Actinobacteria)中。在所有种植方式的土壤中,Proteobacteria相对丰度显著最高,占细菌群落的23%—37%。与其他处理相比,SC土壤中变形菌门的相对丰度显著增高,酸杆菌门的相对丰度减少。拟杆菌门的相对丰度占比为13%—20%,其中WS土壤中拟杆菌门的相对丰度最低。放线菌门的相对丰度占比为4%—14%,与其他处理相比,在FS土壤中相对丰度显著降低。因此,不同种植方式显著改变了土壤细菌优势类群的相对丰度,影响细菌群落结构的组成。

图3 不同种植方式土壤细菌在门水平上的物种组成变化Figure 3 Changes in species annotation to OTUs of soil bacteria from different cropping systems at the phylum level
由图 4可知Gp4、Gp6、Gemmatimonas、Gp1 Gaiella、WPS-1_genera_incertae_sedi、Flavobacteriu、Gp3、Rhizomicrobium、Rhodanobacter、Terrimonas、opitutus和Pedobacter菌群在土壤中相对丰度较高,是东北黑土土壤细菌群落的主要菌属。SC土壤中Gp4相对丰度为2.51%,与SC相比,CK、FS、CS和 WS土壤中Gp4的相对丰度显著增高,分别为9.43%、4.51%、6.35%和5.28%。Gp6的相对丰度在SC中为2.13%,与之相比,其相对丰度在CK、FS、CS和WS土壤中显著增高,分别为5.37%、4.99%、4.03%和4.51%。Gemmatimonas的相对丰度在SC中为1.97%,与之相比,其相对丰度在CK、FS、CS和WS土壤中显著增高,分别为2.61%、2.40%、2.91%和4.56%。可见休耕和轮作处理显著改变了优势菌属在群落中的相对丰度。

图4 不同种植方式土壤细菌在属水平上的物种组成变化Figure 4 Changes in species annotation to OTUs of soil bacteria from different cropping systems at a genus
硝化螺旋菌属(Nitrospira)、厌氧蝇菌(Anaerolinea)、固氮菌(Azotobacter)及产甲烷菌的甲烷八叠球菌属(Methanosarcina,厌氧菌)是土壤中的潜在有益菌,可促进土壤化学循环,保持土壤肥力。根瘤菌(Rhizobium)有利于作物对氮的吸收。芽孢杆菌(Bacills)是增加植物抗病性的潜在有益菌。在FS土壤中,硝化螺旋菌属、厌氧蝇菌、固氮菌及甲烷八叠球菌属相对丰度显著最高(表3)。与SC相比,FS处理的硝化螺旋菌属和固氮菌的相对丰度分别增加了0.47%和0.37%。在5个处理中,根瘤菌在SC、WS和CS土壤中显著增高,在CK和FS中显著降低(P<0.05)。芽孢杆菌的相对丰度在WS中显著最高,在CK和FS土壤中最低(表 3)。可见,不同种植方式显著改变了土壤土壤中有益细菌的相对丰度。

表3 不同种植方式下潜在有益菌的相对丰度Table 3 Relative abundance of potentially beneficial bacteria in different cropping systems
2.5 土壤细菌主要类群相对丰度与土壤化学性质的相关性
将所有样品的细菌菌群与土壤化学性质进行相关分析,发现土壤中Proteobacteria菌群与土壤中P和Cu极显著正相关;Acidobacteria与OM显著正相关,与P、Cu极显著负相关;Bacteroidetes与有效P、Cu、pH和Mn含量显著正相关;Actinobacteria与OM、有效K和Mn呈极显著负相关(表4)。进一步分析发现,土壤中优势细菌属与土壤化学性质存在显著相关性(P<0.05)(表 5)。Gp4与有效Cu和Zn含量极显著负相关;Gp6与OM、有效N极显著正相关,与有效P、Cu含量极显著负相关;Gemmatimonas与有效P和Cu含量极显著负相关;Gp1与有效Fe含量极显著正相关,与pH极显著负相关。同时,土壤中其他优势菌属与土壤OM和有效P、Cu、Mn等呈现显著相关性。土壤中Nitrospira与土壤 OM和 N、Zn、Fe的有效含量显著相关;Rhizobium与土壤OM和N、K、Mn有效含量显著相关;Azotobacter与土壤N、Zn和Fe有效含量显著相关;Bacills与土壤pH和K、Mn有效含量显著相关。可见,土壤中细菌有益菌群与不同土壤化学性质呈现显著相关性(表6)。

表4 土壤细菌优势菌门与土壤化学性质相关关系Table 4 Correlations between dominant phyla of soil bacteria and soil chemical properties

表5 土壤细菌优势菌属与土壤化学性质相关关系Table 5 Correlations between potentially beneficial bacteria and soil chemical properties

表6 潜在有益菌与土壤化学性质的相关性Table 6 Correlations between potentially beneficial bacteria and soil chemical properties
3 讨论
3.1 不同种植方式对细菌群落结构和多样性的影响
土壤微生物群落结构及组成与土壤环境密切相关,种植方式会引起土壤理化性质、水汽热条件的变化,进而改变微生物群落结构。研究证明,变形菌、酸杆菌、拟杆菌和放线菌是农田土壤细菌中的优势类群(Liu et al.,2017)。不同种植方式在一定程度上改变了变形菌、拟杆菌、酸杆菌门的相对丰度(Li et al.,2014)。在本研究中,5种种植方式下变形菌是土壤中最丰富的的类群,相对丰度在23%—37%,这与前人研究结果一致(沈冰洁等,2015;朱琳等,2017)。与连作相比,轮作处理显著降低了土壤中变形菌的相对丰度,增加了酸杆菌的相对丰度,这与前人研究结果不一致。朱英波等(2014)对黑龙江富锦地区两个相邻的大豆土壤细菌多样性分析发现,轮作土壤以变形菌为优势类群,连作以酸杆菌为优势类群。研究结果不同表明,不同门类细菌在不同生态区和不同种植方式下所占比例不同。
大豆连作障碍一直是研究的热点问题,针对不同土壤、不同种植方式下大豆田土壤微生物变化已有诸多研究。由于生态环境、种植方式以及种植年限不同,微生物类群及其丰度变化的结果也不尽相同。众多研究证明,大豆连作后大豆根际分泌物及单一的田间管理导致土壤生态环境向着利于病原真菌积累的方向偏移(Li et al.,2014)。研究证明黑龙江西部地区大豆经过短期连作后,大豆根际细菌群落多样性和丰富度增加(王芳等,2018)。轮作和大豆短期连作(2年和4年连作)土壤多样性高于长期连作(连作7年)(陈雪丽,2015)。因此,大豆土壤细菌群落丰度主要是随着大豆连作年限的增加呈现先增加而后降低的现象。也有研究证明长期连作导致健康土壤向致病土壤再向抑病土壤的转变(Schreiner et al.,2010)。研究证明,土壤抑病功能是由不同功能生物类群结构的转变导致的,包括细菌优势菌群丰度变更和土壤中有益菌的增加(王闯进,2014)。研究证明长期连作(20年)土壤的优势细菌与轮作土壤的相似度更高,并且一些有益菌的丰度高于轮作土壤的,如与固氮有关的根瘤菌,以及促进植物生长和对病原菌有防控作用的芽孢杆菌的丰度在连作4年、10年土壤中都高于在轮作土壤中。硝化螺旋菌属、厌氧蝇菌、固氮菌和甲烷八叠球菌属有助于土壤中的亚硝酸氧化成硝酸盐,进而提高土壤的氮肥(Li et al.,2018)。本研究中大豆连作促进了土壤根瘤菌相对丰度的增高,小麦-大豆轮作促进了芽孢杆菌相对丰度的增高,而休耕-大豆轮作促进了硝化螺旋菌属、厌氧蝇菌、固氮菌和甲烷八叠球菌属相对丰度的增高。同时玉米-大豆和小麦-大豆轮作土壤中的硝化螺旋菌属也显著高于大豆连作土壤中的。这一现象说明在大豆连作土壤上实施休耕-大豆轮作及其他轮作更利于增加土壤中有益菌群的丰度,进而提高土壤地力。
土壤微生物群落结构和多样性决定了土壤微生物群落的稳定性及其抗逆能力,是土壤生态系统可持续发展的重要指标(Smith et al.,2008)。Yin et al.(2010)研究显示,与连作相比,轮作体系显著改变微生物的群落结构。然而,Venter et al(.2016)发现轮作条件下微生物群落并不发生改变,且不同的研究手段、轮作方式、种植年限等因素会导致研究结果产生差异。本研究证实,大豆连作土壤中细菌菌群丰度(Chao1/ACE)和菌群多样性最低,3中轮作种植方式都改变了土壤细菌群落结构,其中休耕-大豆轮作能够显著增加土壤中细菌菌群丰度(Chao1/ACE)和菌群多样性(Shannon)。通过分析韦恩图发现,休耕显著增加了土壤中特有 OTUs的数量,因此提高了土壤微生物多样性数值。研究表明休耕有利于耕层中水分与温度的保持,因此促进了土壤微生物的生长繁殖(徐光辉等,2018)。本研究中小麦-大豆和玉米-大豆轮作土壤中的细菌群落丰度和多样性也高于大豆连作土壤中的。轮作通过增加作物的种类和生物量的方式增加土壤微生物的食物来源,进而导致土壤微生物多样性增加(Wang et al.,2011)。因此本研究证实,在大豆连作土壤上增加休耕和轮作制度可以改善土壤细菌群落结构,增加群落多样性(Navarro et al.,2013;Ai et al.,2015)。
3.2 不同种植方式土壤化学性质和微生物的相关性
据报道,不同的作物种植方式使得土壤中营养元素利用和转化效率不同,进而使土壤形成不同的肥力性质(Xun et al.,2015)。连作可能导致土壤养分失调(刘株秀等,2019)。研究证明,大豆连作胁迫导致土壤有效Zn、Mn和Fe增加(韩丽梅等,2000)。研究发现,与大豆连作相比,种植玉米、小麦对土壤P的需求相对减少,利于土壤有效磷的积累(李玉洁等,2015)。也有研究证明轮作5年和大豆连作13年显著增加了土壤pH和有效养分含量(刘株秀等,2019)。本研究发现大豆连作土壤中OM和有效N、K、Zn、Fe、Mn显著增加,有效P显著降低;休耕-大豆轮作消耗了土壤中有效Mn含量;小麦-大豆轮作消耗了土壤OM和有效N、Fe的含量,而促进土壤有效P和Cu含量的积累。因此,不同生态环境、种植方式和种植时间会显著影响土壤化学性质。
研究证明,土壤化学性质影响土壤微生物群落结构,土壤中OM和有效N含量的增加可以促进细菌多样性的形成(Liu et al.,2014;Li et al.,2018)。随着土壤养分的积累,变形菌的相对丰度增加(Zeng et al.,2017)。通过相关性分析,本研究土壤中变形菌与土壤中有效P和Cu含量显著相关;拟杆菌与土壤有效P、Cu和Mn的含量显著相关;放线菌与土壤中OM和有效Mn含量显著负相关;酸杆菌与OM正相关,与有效Cu负相关。一些研究证明土壤 pH是决定细菌群落演替的主导因子(Pii et al.,2015),而本研究证明OM、P、Cu、Mn与土壤细菌群落相关性更大,这可能是由于本研究的土壤样本间pH差异不显著,而微量元素差异显著。实验结果证明,种植方式、土壤化学性质和土壤微生物群落结构是具有相关性(Ofek et al.,2014;Pang et al.,2017)。以上研究结果表明,不同种植方式及土壤化学性质对各类土壤细菌群落影响不一致。
4 结论
与大豆连作处理相比,休耕与轮作显著改变了土壤细菌群落结构。休耕-大豆能够显著促进土壤细菌群落相对丰度和多样性的形成,小麦-大豆轮促进效果作次之;休耕与休耕-大豆细菌群落结构相近,并且促进了土壤中特有OTUs的形成。休耕、轮作处理显著降低了变形菌门的相对丰度,增加了酸杆菌门的相对丰度;同时休耕和轮作处理显著改变了土壤中细菌优势菌属在群落中的相对丰度,尤其是显著增加了Gp4、Gp6和Gemmatimonas的相对丰度。休耕-大豆轮作处理显著增加了土壤中硝化螺旋菌属、厌氧蝇菌、固氮菌及产甲烷菌的甲烷八叠球菌属的相对丰度,小麦-大豆轮作能够促进土壤中芽孢杆菌的相对丰度,有利于增加土壤抗病性。因此在大豆连作地区实施休耕-大豆轮作和小麦-大豆轮作更利于土壤细菌群落丰度和多样性的形成。同时,本研究发现不同种植方式显著改变了土壤化学性质,并且土壤中优势菌群与土壤OM,有效P、Cu、Mn等化学性质的含量呈现显著相关性。因此,种植方式和土壤化学性质能够调控土壤细菌组成结构。
