增温和植物去除对青藏高原东部高寒灌丛土壤不同形态氮的影响
2022-05-24刘美马志良
刘美,马志良
1. 绵阳师范学院/生态安全与保护四川省重点实验室,四川 绵阳 621000;2. 西华师范大学生命科学学院,四川 南充 637009
高寒生态系统植物生长发育、物质合成和转运等生命活动均需要氮素参与(Miller et al.,2003)。由于高寒生态系统土壤生态过程长期受低温影响,土壤氮转化过程缓慢,土壤有效氮含量通常不能满足植物生长发育的需求,土壤氮素有效性已成为植物群落生长最主要限制因子之一(Du et al.,2020)。土壤中含量最丰富的大分子有机氮不能被植物根系直接吸收,但能通过土壤微生物和酶矿化过程分解为无机氮和氨基酸等有效形态被植物根系吸收利用(Chen et al.,2019)。同时,土壤微生物生物量氮也是土壤氮素的生物贮存库,在氮循环中具有不可替代的作用(Buchkowski et al.,2015)。相关研究表明,土壤温度和水分条件、冻融循环、大气氮沉降以及植物群落、土壤生物活动、酶活性等非生物与生物因素共同调控土壤氮转化过程,决定着土壤中不同形态氮含量(Dawes et al.,2017a;Nie et al.,2017)。
岷江源区位于青藏高原东部,该区域土壤氮库容量巨大,维持着区域氮生物地球化学循环(Zhao et al.,2018)。高寒灌丛是岷江源区最主要的植被类型之一,该植被类型对气候变暖的响应十分敏感(IPCC,2014)。气候变暖导致该区域高寒灌丛分布上限不断爬升,分布面积逐步扩张,从而使高寒灌丛在区域生态系统氮循环中发挥着越来越重要的作用(Miehe et al.,2018)。同时,随着全球温度的持续升高,凋落物分解和细根周转速率提高,土壤有机质输入、矿化过程加快,将加速该区域土壤氮转化过程(Ma et al.,2018)。植物群落作为影响土壤氮转化和有效氮含量的主要生物因子,对土壤氮素动态具有双重作用。具体来说,生长季节植物从土壤中吸收氮素,从而降低土壤氮含量。植物群落也可通过凋落物、细根周转和根系分泌物等方式向土壤中输入有机氮,为土壤氮矿化提供有效底物,从而提高土壤氮含量(Peng et al.,2019)。气候变暖将促进高寒灌丛植物群落初级生产,增加植物对土壤有效氮的利用,同时增加根系分泌物、凋落物、死亡细根等形式的氮输入(Butler et al.,2012)。由此可见,植物群落和气候变暖将共同调控高寒灌丛生态系统土壤氮转化过程,进而影响土壤不同形态氮含量,但其具体机制仍需深入探讨。有研究表明,增温显著影响了苔原生态系统土壤不同形态氮含量,其影响程度受植物群落的调控(Alatalo et al.,2017)。在高山树线区域,受不同植物生长的影响,增温使土壤铵态氮和可溶性有机氮含量持续增加(Dawes et al.,2017b),但对其他形态氮含量影响不显著。目前关于高寒灌丛土壤不同形态氮含量动态及其影响因素的研究报道较少,这在一定程度上限制了我们对高寒灌丛土壤氮转化和植物氮素营养过程的认识。
窄叶鲜卑花(Sibiraea angustata)灌丛是青藏高原东部具有代表性的高寒灌丛类型,在该区域具有较大的分布面积(冶民生等,2009)。近年来,人类活动加剧(如过度放牧、砍伐等)使该区域植物群落覆盖度与多样性迅速减少(Zhu et al.,2015),影响根系分泌物和凋落物生产、细根周转等土壤氮输入过程。此外,植物群落结构变化还会导致土壤微环境条件改变,从而间接影响土壤氮转化过程和不同形态氮含量(Liu et al.,2020)。因此,本研究以青藏高原东部窄叶鲜卑花灌丛为对象,开展模拟增温试验,并设置去除/不去除植物处理,研究高寒灌丛土壤不同形态氮含量对增温和植物去除的响应,以更清晰地认识气候变暖背景下人类活动加剧对高寒灌丛土壤氮循环过程的影响。
1 材料与方法
1.1 研究区概况
研究区位于四川省阿坝州松潘县川主寺镇卡卡沟的高寒灌丛生态系统长期定位研究场(32°58′N,103°40′E),海拔 3300 m,距离岷江源3.5 km。气候类型为典型的高原山地气候,昼夜温差大。年均温为4.8 ℃,年均降水量693 mm,降水主要集中在 5—8月。研究区域内灌木层主要有窄叶鲜卑花、山生柳(Salix oritrepha)、金露梅(Potentilla fruticosa)、高山绣线菊(Spiraea alpina)、刺黑珠(Berberis sargentiana)等;草本层主要有矮蒿草(Kobresia humilis)、条纹龙胆(Gentiana striata)、羊茅(Festuca ovina)、蓝玉簪龙胆(G.veitchiorum)、黄花野青茅(Deyeuxia flavens)、红花绿绒蒿(Meconopsis punicea)、钟花报春(Primula sikkimensis)等。研究区域窄叶鲜卑花平均高度为 (1.5±0.1) m,平均丛径为 (0.5±0.2) m,平均盖度为72%,分枝数为15—20。土壤类型为雏形土,土壤pH 5.79,土壤容重为0.67 g·cm-3,土壤有机碳质量分数为89.24 g·kg-1,土壤全氮质量分数为 7.73 g·kg-1。
1.2 试验设计
植物处理方式:于2015年10月初,选择研究区域内物种组成、群落结构和生境相对均匀的窄叶鲜卑花灌丛群落。在去除植物处理中(样方面积 5 m×5 m),首先去除地表凋落物层及所有灌木和草本植物地上生物量。然后沿样方的边缘挖掘一条宽20 cm、深75 cm的壕沟,将2 mm厚的聚乙烯塑料薄膜贴放在壕沟周围。然后用原状土回填壕沟,以避免邻近灌木根系对样方造成干扰。于 2016年 5—10月每月下旬定期去除样方内新生长出的草本植物。在不去除植物处理样方中(样方面积5 m×5 m),选择一丛长势良好、高度大体一致、具有相似生长势的窄叶鲜卑花植株。在样方内不对植物进行任何处理,保持窄叶鲜卑花植株自然生长。
模拟增温试验:采用开顶式生长室(Open top chamber,OTC)对环境进行增温。基于窄叶鲜卑花灌丛群落的平均盖度和高度,OTC材料选用1.6 m×1.6 m×5 mm的有机玻璃板,有机玻璃板(材质:聚甲基丙烯酸甲酯;生产厂家:成都天中广告材料有限公司)的透光率达92%以上。将4块有机玻璃板包围至少1丛窄叶鲜卑花植株,制成正立方体型OTC。OTC下端插入土壤1 cm,地表增温面积为2.56 m2。窄叶鲜卑花植株生长于OTC装置的中央。在之后的土壤样品采集过程中,尽量避免对OTC内窄叶鲜卑花植株生长造成影响。同时在每个OTC的附近随机设置1个对照(不增温)样方。增温和对照样方所选择的窄叶鲜卑花植株均具有一致的高度、盖度和相似的生长势。所有处理均设置4个重复。在整个样地内共建立 8套 OTC,去除/不去除植物处理样地各4套。
1.3 空气温度、土壤温度和水分监测
于2016年5月初开始进行空气温度、土壤温度和水分监测。对照和增温处理地下5 cm土层土壤温度和地上70 cm空气温度分别采用纽扣式温度传感器(DS1921G-F5#,Maxim/Dallas semiconductor,Sunnyvale,California,美国)进行监测。所有的地下温度传感器均埋设在样方的中央。温度传感器设置为每2小时读取1次数据,自动记录试验期间空气温度和土壤温度变化。所得数据用于月平均空气温度和土壤温度的计算。并从2016年生长季节开始,逐月使用便携式土壤水分测定仪(TRIME TDR,IMKO,德国)对地下5 cm土层土壤含水量进行监测。
1.4 土壤样品采集与指标分析
分别于2016年灌丛植物群落生长季初期(5月20日)、中期(7月21日)和末期(9月21日)进行土壤样品采集。土壤样品取样前,首先移除地表凋落物及其他杂物。取样时,在每个取样样方内,采用五点取样法,沿对角线用土钻取0—15 cm土壤,每个样方内取5钻。然后将5个土样混合均匀作为一个混合土壤样品。在不去除植物样方内取土样时,尽量避免对灌丛植物造成干扰。所有土样于4 ℃下保存并带回实验室。去除石块、动植物残体后,过2 mm筛。各处理土壤样品分为两部分,一部分贮存于4 ℃下,用于土壤含水量、硝态氮、铵态氮和微生物生物量氮含量的测定;另一部分风干后用于土壤全氮含量的测定。土壤含水量采用烘干法测定。土壤全氮含量采用 C、N元素分析仪(Multi-N/C 2100,Analytic Jena AG,Jena,Germany)进行测定。土壤硝态氮含量采用紫外比色法测定,土壤铵态氮含量采用靛酚蓝比色法测定。土壤微生物生物量氮采用氯仿熏蒸-K2SO4浸提法进行测定,转换系数为0.45。土壤全氮、硝态氮、铵态氮和微生物生物量氮含量换算成mg·kg-1(以干土质量计)。所有土壤样品硝态氮、铵态氮和微生物生物量氮含量的测定均在采样后1周内完成。土壤无机氮含量为土壤硝态氮与铵态氮含量之和,土壤有机氮含量为土壤全氮与无机氮含量之差。
1.5 数据处理与统计分析
采用重复测量方差分析检验植物、增温和采样时期以及它们之间的交互作用对土壤不同形态氮含量的影响(P=0.05)。采用最小显著差异法(LSD)进行同一采样时期不同处理下不同形态氮含量的多重比较(P=0.05)。采用多元线性回归分析检测土壤氮含量与空气温度、土壤温度和土壤水分之间的关系。应用SPSS 20.0软件进行所有数据的统计分析,应用Origin 8.5软件作图。图表中数据为平均值±标准差。
2 结果
2.1 高寒灌丛生长季温度和水分动态
空气温度和土壤温度均随生长季表现出先升高后降低的趋势;土壤水分除在6月因取样前降雨而出现大幅度提高以外,也表现出随生长季先升高后降低的趋势。整个生长季内,与对照样方相比,增温分别使去除/不去除植物样方空气温度增加了1.6 ℃(P<0.05)和 0.5 ℃(P>0.05),分别使土壤温度平均增加了 0.6 ℃(P>0.05)和 1.3 ℃(P<0.05)(图 1a、b)。同时,增温使去除/不去除植物样方土壤水分含量显著降低了1.4%(P>0.05)和3.2%(P<0.05)(图1c)。
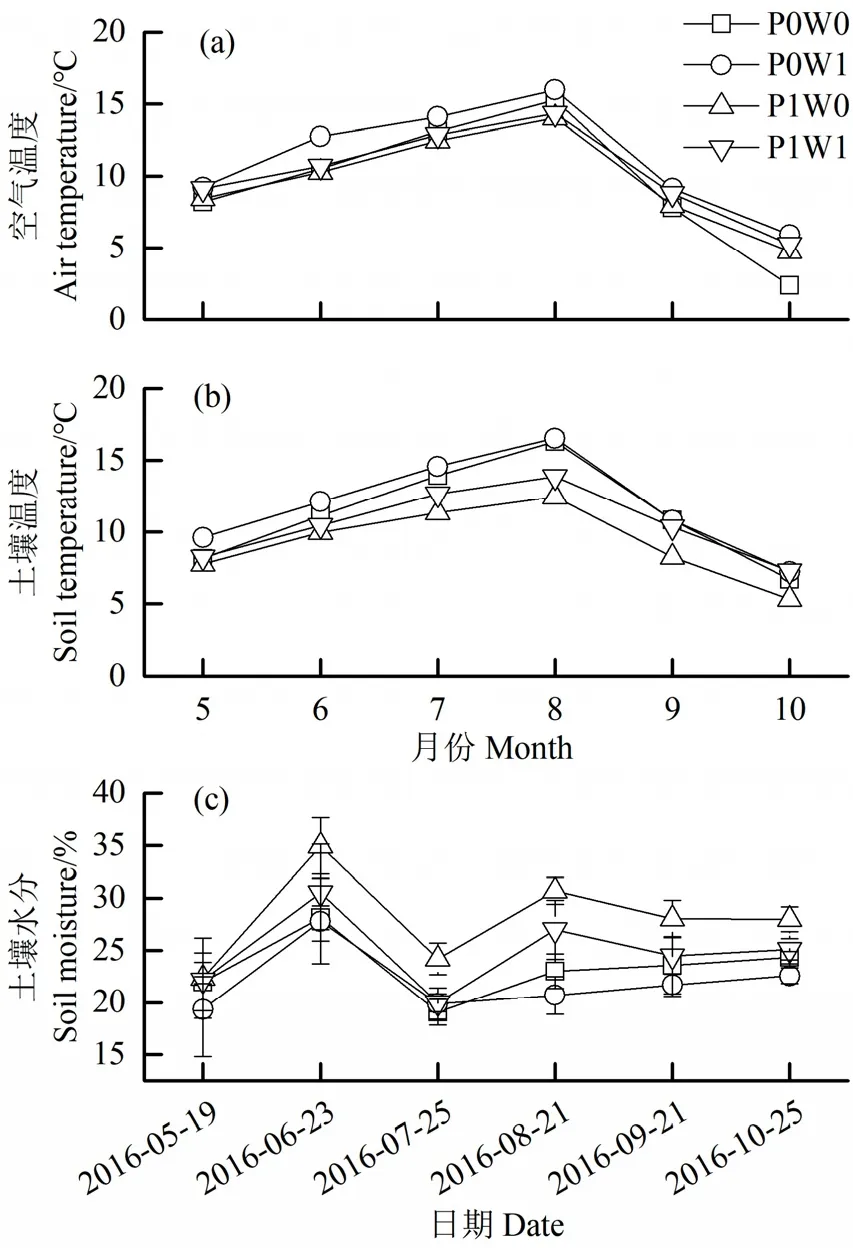
图1 试验期间各处理下70 cm空气日平均温度(a)、5 cm土层土壤日平均温度(b)和5 cm土层土壤含水量(c)的季节动态Figure 1 Dynamics of daily mean air temperature at 70 cm above the ground (a), daily mean soil temperature in 0-5 cm soil layer (b) and soil moisture in 0-5 cm soil layer (c) relative to treatment
2.2 增温与去除植物对土壤全氮的影响
生长季不同时期各处理土壤全氮含量存在显著差异(表 1),且均表现为一直增加的趋势,在生长季末期达到最大值(图 2)。增温对土壤全氮含量的影响因季节和植物处理方式而存在明显差异(图2,表1)。在生长季中期(7月)和末期(9月),增温使去除植物样方土壤全氮含量显著降低了14.2%和13.1%(P<0.05)。在不去除植物样方,增温未显著影响整个生长季土壤全氮含量。植物处理方式对土壤全氮含量的影响仅出现在对照样方(不增温)。与去除植物处理相比,不去除植物处理使生长季末期土壤全氮含量显著降低了 11.6%(P<0.05)(图2,表1)。增温和植物处理的交互作用对土壤全氮含量无显著影响(表 1)。多元线性回归分析结果表明,土壤全氮含量受空气温度和土壤温度的显著影响,二者共同解释了土壤全氮含量变异的20.7%(表2)。
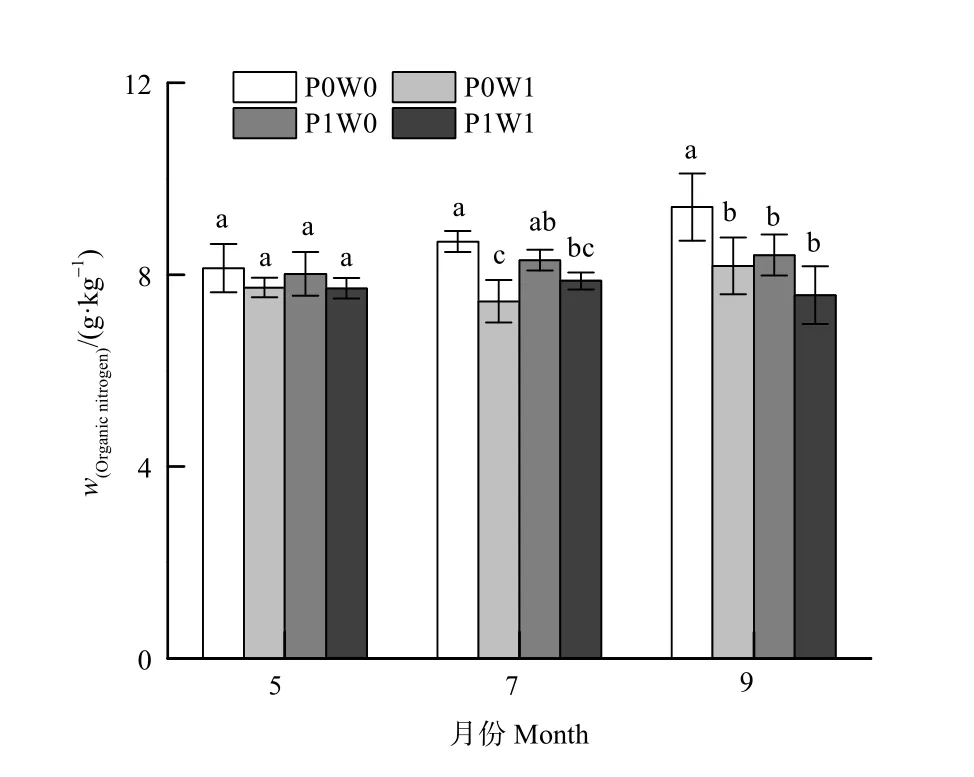
图2 不同处理下土壤全氮含量的季节动态Figure 2 Seasonal dynamics of the soil total nitrogen content relative to treatment

表1 增温、植物处理以及取样时间对土壤不同形态氮含量的重复测量方差分析结果Table 1 ANOVA of the repeated measurements of different forms of soil N relative to treatment and sampling date
2.3 增温与去除植物对土壤有机氮的影响
与土壤全氮含量类似,生长季不同时期各处理土壤有机氮含量存在显著差异(表 1),且均表现为一直增加的趋势,在生长季末期达到最大值(图3)。生长季不同时期增温对土壤有机氮含量的影响不同,且因季节和植物处理方式而存在明显差异(图3,表1)。在生长季中期和末期,增温使去除植物样方土壤有机氮含量显著降低了 14.3%和13.5%(P<0.05)。在不去除植物样方,增温未显著影响整个生长季土壤有机氮含量。植物处理方式对土壤有机氮含量的影响也仅出现在对照样方,与去除植物处理相比,不去除植物处理使生长季末期土壤有机氮含量显著降低了 12.1%(P<0.05)(图3,表1)。增温和植物处理的交互作用对土壤有机氮含量无显著影响(表 1)。空气温度和土壤温度共同影响土壤有机氮含量,二者解释了土壤有机氮含量变异的20.6%(表2)。

表2 土壤氮含量与空气温度、土壤温度和土壤水分之间的回归分析Table 2 Regression analysis of soil nitrogen contents with air temperature, soil temperature and soil moisture content

图3 不同处理下土壤有机氮含量的季节动态Figure 3 Seasonal dynamics of the soil organic nitrogen content relative to treatment
2.4 增温与去除植物对土壤无机氮的影响
生长季不同时期各处理土壤无机氮含量存在显著差异(表 1),各处理土壤无机氮含量随生长季均表现为先增加后降低的变化趋势,在生长季中期达到最大值(图 4)。在去除植物样方,增温仅在生长季末期使土壤无机氮含量显著降低了16.1%(图4,表1)。在不去除植物样方,增温在生长季中期使土壤无机氮含量显著升高了 17.5%(P<0.05),在生长季末期使其显著降低了 21.8%(P<0.05)。植物处理方式在生长季不同时期对土壤无机氮含量的影响不同。在对照样方,不去除植物处理在生长季中期使土壤无机氮含量显著降低了29.3%(P<0.05),在生长季末期使其显著升高了18.7%(P<0.05)。而在增温样方,不去除植物处理仅在生长季末期使土壤无机氮含量显著升高了12.7%(图4,表1)。增温和植物的交互作用对土壤无机氮含量无显著影响(表 1)。土壤无机氮含量受空气温度、土壤温度和土壤水分的显著影响,三者共同解释了土壤无机氮含量变异的35.5%(表2)。

图4 不同处理下土壤无机氮含量的季节动态Figure 4 Seasonal dynamics of the soil inorganic nitrogen content relative to treatment
2.5 增温与去除植物对土壤微生物生物量氮的影响
生长季不同时期各处理土壤微生物生物量氮含量存在显著差异(表 1),且均表现为一直降低的趋势,生长季初期土壤微生物生物量氮含量显著高于生长季中期和末期(图 5)。生长季不同时期增温对土壤微生物生物量氮含量的影响不同,也因季节和植物处理方式存在明显差异(图5,表1)。在去除植物样方,增温在生长季初期(5月)使土壤微生物生物量氮含量显著升高了 37.7%(P<0.05)。在不去除植物样方,增温仅在生长季中期使土壤微生物生物量氮含量显著升高了 122.1%(P<0.05)。植物处理方式对土壤微生物生物量氮含量的影响因增温和季节而不同(图 5,表 1)。在对照样方,不去除植物处理使生长季初期土壤微生物生物量氮含量显著升高了 26.0%(P<0.05),使生长季中期土壤微生物生物量氮含量显著降低了68.9%(P<0.05)。而在增温样方,不去除植物处理仅在生长季中期使土壤微生物生物量氮含量显著升高了 49.2%(P<0.05)。增温和植物的交互作用对土壤微生物生物量氮含量的影响达到显著水平(表 1)。多元线性回归分析结果表明,土壤微生物生物量氮含量受空气温度、土壤温度和土壤水分的显著影响,三者共同解释了土壤微生物生物量氮含量变异的70.7%(表2)。

图5 不同处理下土壤微生物生物量氮含量的季节动态Figure 5 Seasonal dynamics of the soil microbial biomass nitrogen content relative to treatment
3 讨论
3.1 高寒灌丛生长季土壤不同形态氮含量的季节动态
受气候季节波动的影响,高寒灌丛植物生长发育和土壤生物群落结构、生理活性表现出明显的季节变化(Prevé et al.,2017),将对土壤氮转化动态产生一定影响,从而改变土壤不同形态氮含量。本研究结果显示,窄叶鲜卑花高寒灌丛土壤全氮和有机氮含量的季节动态一致,均表现为随生长季一直增加的趋势(图2,图3和表1)。土壤有机氮占土壤全氮的绝大部分,因此土壤有机氮和全氮含量动态往往是协同变化的。本研究中,生长季节窄叶鲜卑花灌丛植物凋落物和根系分泌物持续输入是土壤全氮和有机氮含量一直增加的最主要原因(Liu et al.,2021)。同时,土壤有机质的矿化为土壤微生物提供碳源和氮源,土壤有机碳、氮含量存在耦合关系(Yan et al.,2019)。窄叶鲜卑花高寒灌丛生长季土壤有机碳含量一直维持在较高的水平也可能是土壤有机氮和全氮含量不断增加的一个原因(Liu et al.,2021)。与土壤有机氮含量的季节动态不同,土壤无机氮含量表现出先增加后降低的趋势(图 4)。土壤无机氮含量在生长季中期达到峰值的主要原因是此时期土壤温度升高,含水量下降,土壤透气性好,有利于土壤有机氮矿化(Hu et al.,2018)。较高的土壤无机氮含量将能满足生长季中期植物群落和土壤微生物生长对土壤有效氮的需求。前期研究发现,受高寒灌丛生长季不同时期土壤氨化和硝化作用不同的影响,该区域窄叶鲜卑花灌丛土壤铵态氮与硝态氮含量在生长季不同时期表现出明显不同的动态(马志良等,2018a;刘美等,2021)。这预示着高寒灌丛植物和土壤微生物在生长季不同时期对土壤无机氮的吸收利用格局可能发生明显变化。土壤微生物生物量氮含量则随生长季节一直降低(图 5),这可能与土壤养分资源限制有关。高寒灌丛非生长季土壤表面覆盖一层较厚的雪被,雪被下较好的水热条件有利于土壤微生物繁殖。此时期凋落物分解和土壤有机质矿化使土壤中积累了大量有效氮(Brin et al.,2018),植物群落刚刚萌芽,土壤微生物对土壤有效氮的竞争在此时期占据优势,从而导致了土壤微生物生物量氮含量较高。而在长季中期和末期,植物群落生长消耗了大量的土壤有效氮,植物群落对土壤氮素的竞争占主导地位,土壤微生物生长、繁殖受到严重的养分限制,导致土壤微生物生物量氮含量迅速降低。土壤温度和水分季节性变化导致土壤微生物群落结构向真菌和格兰氏阳性菌方向转变也可能是土壤微生物生物量氮含量降低的一个原因(Ma et al.,2018)。
3.2 增温对土壤不同形态氮含量的影响
土壤温度升高可促进土壤氮矿化,导致土壤有机氮含量降低而无机氮含量升高(Liu et al.,2016)。同时,温度升高也将促进植物和土壤微生物群落生长,增加植物和土壤微生物对土壤无机氮的竞争能力(唐波等,2016)。本研究显示,增温使去除植物样方生长季中期和末期土壤全氮和有机氮含量显著降低(图2和图3)。这是因为增温提高了土壤微生物活性,促进了土壤有机氮矿化过程(Peng et al.,2019)。同时,增温使生长季土壤脲酶活性显著提高10.8%—27.8%(马志良等,2018b),均有利于土壤氮矿化过程。马彩霞等(2019)在川西高山林线区域的研究结果表明,增温显著促进土壤微生物活性,从而使土壤有机层和矿质土壤层可溶性有机氮含量显著降低。本研究还发现,增温在生长季末期使去除植物样方土壤无机氮含量显著降低(图 4)。这主要是因为生长季末期土壤温度较低,土壤含水量一直保持在25%左右(图1),土壤透气性不如生长季中期。增温提高了喜厌氧环境的反硝化细菌的丰富度和活性。反硝化作用的增强使大量的土壤无机氮以N2O和N2的形式释放至大气中(Deng et al.,2016)。增温在生长季初期使去除植物样方土壤微生物生物量氮含量显著升高(图5),表明此时期增温能够促进土壤微生物群落对土壤氮素的利用。此时期植物群落正处于萌芽阶段,增温对土壤微生物群落氮素竞争能力的促进作用明显强于对植物群落,使得生长季初期土壤微生物生物量氮含量显著增加。生长季中期土壤温度升高使土壤养分有效性显著提高,有利于土壤微生物活动,促进土壤有机质的矿化,从而使此时期不去除植物样方土壤无机氮和微生物生物量氮含量显著升高(图4和图5)。而植物和土壤微生物对土壤氮素的吸收利用则是生长季末期土壤无机氮含量显著降低的主要原因。同时,增温使反硝化作用增强加剧了土壤无机氮的消耗,增温导致土壤含水量下降也可能部分抵消了增温对土壤微生物氮矿化的正效应(Deng et al.,2016)。然而,增温未显著影响整个生长季不去除植物样方土壤全氮和有机氮含量(图2和图3),这表明增温导致植物来源土壤有机质增加能弥补矿化作用增强所消耗的土壤有机氮,从而使土壤全氮和有机氮含量维持在一个较为稳定的状态。
3.3 植物对土壤不同形态氮含量的影响
本研究中,与去除植物处理相比,不去除植物处理在生长季末期显著降低了对照样方土壤全氮和有机氮含量(图2和图3)。虽然凋落物分解、细根周转和根系分泌物向土壤中持续输入有机氮,但是灌丛植物和土壤微生物群落在生长季吸收了大量的土壤无机氮。土壤无机氮供应不足诱使土壤有机氮矿化加速,而且新鲜的有机质输入可能通过激发效应刺激土壤有机质矿化(Yin et al.,2018),因而在生长季末期土壤有机氮含量显著降低(图3)。然而,不去除植物处理未显著影响增温样方土壤全氮和有机氮含量(图2和图3),这表明高寒灌丛植物生长能够调节土壤有机氮矿化对增温的响应。而在生长季末期,不去除植物处理使对照样方土壤无机氮含量升高(图 4)。此时期温度大幅降低,植物群落开始进入落叶休眠期,对土壤无机氮的需求降低;土壤微生物活动也受到土壤温度、水分条件的限制,从而缓解了植物和土壤微生物对土壤无机氮的竞争性吸收。在增温样方,不去除植物处理仅在生长季末期使土壤无机氮含量显著升高(图 4),此时期植物细根周转和凋落物输入为土壤微生物提供更多有效底物,加速增温条件下的土壤氮矿化作用。在对照样方,植物群落在生长季初期可能有利于土壤微生物生长、繁殖,因而增加土壤微生物生物量氮含量;而在生长季中期,植物群落与土壤微生物之间发生强烈的养分竞争,因而使土壤微生物生物量氮含量显著降低。在增温样方,植物群落细根周转、分解加快,分泌大量的根系分泌物,为土壤微生物固定土壤氮素提供了有利条件,因此不去除植物处理显著增加土壤微生物生物量氮含量。同时,增温和植物的交互作用仅对土壤微生物生物量氮含量具有显著影响,而对其他3种形态氮含量的影响不显著(表 1),说明高寒灌丛土壤微生物生物量氮含量对增温和去除植物引起的土壤环境变化较其他形态氮更加敏感(Steinauer et al.,2015)。未来气候变暖和植被干扰可显著影响高寒灌丛土壤微生物过程,进而改变土壤氮转化过程。多元回归分析结果也表明,空气温度、土壤温度和土壤水分变化共同影响土壤无机氮和微生物生物量氮含量,而仅有空气温度和土壤温度变化与土壤全氮和有机氮含量有关,土壤水分变化对土壤全氮和有机氮含量的影响不明显。
4 结论
青藏高原东部窄叶鲜卑花高寒灌丛土壤不同形态氮含量受增温和不同植物处理方式的影响,并且植物生长可以调节土壤不同形态氮含量对增温的响应。空气温度、土壤温度、土壤水分和微生物群落等非生物和生物因子是影响高寒灌丛土壤不同形态氮含量的主要因素。未来气候变暖和人类活动对该区域高寒灌丛植被覆盖度的改变将会对土壤不同形态氮含量产生复杂而深刻的影响,进而加速高寒灌丛土壤氮转化和循环过程。
