不同杉木林分类型植物多样性及其土壤碳氮关系的研究
2022-05-24胡靓达周海菊黄永珍姚贤宇叶绍明喻素芳
胡靓达,周海菊,黄永珍,姚贤宇,叶绍明,喻素芳
广西大学林学院,广西 南宁 530004
人工林作为森林资源的重要组分,对于森林的可持续经营意义重大。目前,中国人工林纯林面积大,生态系统结构和功能简单,易造成林产品单一、生物多样性降低、病虫害蔓延、地力衰退和生产力低下等诸多问题(周霆等,2008)。营造混交林在一定程度上可改善土壤理化性质、提升林分涵养水源功能(王青天,2014)、减少林木病虫害发生(叶绍明等,2010),进而提高林分的生物、生境多样性(Chianucci et al.,2016;Kooijman et al.,2019)及林分生产力,从而建立更稳定的森林生态系统。然而,混交树种选择可能会产生不同的经营效果,如杉木(Cunninghamia lanceolate)与木荷(Schima superba)以1∶1进行行间混交可促进杉木生长,混交林中的杉木树高、胸径皆大于杉木纯林(李勇,2016),而与细叶青冈(Cyclobalanopsis gracilis)混交杉木的生长反而受到了抑制(刘雨晖,2018a);桉树(Eucalyptus)与不同树种混交对林下植物物种多样性有显著影响,尤其与望天树(Parashorea chinensis)、格木(Erythrophleum fordii)和红锥(Castanopsis hystrix)混交更具优势(庞圣江等,2020)。混交林经营不仅增加了林下植被多样性,还能提高土壤养分含量,维持地立平衡(刘世荣等,2018),如石亮等(2021)对樟子松(Pinus sylvestris var. mongolica)人工林混交改造后的土壤养分研究发现,38 a生的樟子松与樟子松、银中杨(Populus alba)、色木槭(Acer mono)幼苗混交后,土壤养分均高于沙地樟子松纯林,其中有机质、碱解氮含量在樟子松×色木槭、樟子松×银中杨混交林达到最高,但有效磷含量则低于樟子松×樟子松混交林。
植被和土壤是一个密不可分的整体,植物通过根系下扎、分泌物、凋落物及调节小气候等影响土壤性状(王光州等,2021),同时植物个体之间以及植物种间竞争土壤资源也会对植物群落物种组成和生长产生影响(余轩等,2021)。生物多样性是人工植被恢复重要评价指标,而植物多样性是生物多样性在植物水平上的表现形式(甄倩等,2020),是基于群落的物种数和个体数及其分布均匀程度的统计量(赵耀等,2018),它与土壤环境的协调程度反映出森林生态系统的稳定性。因此,了解植物多样性与土壤养分因子的相互关系对评价人工林可持续经营水平有着重要意义。
杉木是中国南方亚热带地区的主要用材树种之一,因其生长快、适应性强、经济效益高、材质好等特点而被广泛种植(李萌等,2020)。由于杉木纯林经营引发系列生态问题,严重制约林地的可持续利用(明安刚,2017)。目前,已有大量试验证明杉木人工混交林可以有效促进土壤中的养分循环(姜俊等,2019),改善土壤的理化性质(徐海东等,2020),增强土壤肥力,促进林木生长(唐学君等,2019),提升林分的涵养水源功能等(张莉,2019)。相较于杉木纯林,混交林更有利于提高植物多样性,维持生态系统稳定(黄冬梅,2017;刘雨晖,2018)。同时,前人研究发现土壤有机碳含量和有机碳密度在很大程度上依赖于植被群落多样性(刘林馨等,2018),土壤全氮含量与植物多样性指数呈显著正相关关系(刘俊杰等,2021),植物多样性与土壤碳氮含量之间关系密切。然而,不同树种与杉木混交后植物多样性与土壤碳、氮变化的关系鲜见报道。因此,本文以位于广西壮族自治区凭祥市的中国林业科学研究院热带林业研究中心伏波实验场的杉木×红锥混交林、杉木×大叶栎(Quercus griffithii)混交林以及杉木纯林为研究对象,旨在探讨红锥、大叶栎与杉木混交后对植被多样性特征及其土壤碳、氮含量的影响,并分析土壤碳、氮含量及碳氮比变化与植被多样性的相关性,为杉木人工林的可持续经营提供科学依据。
1 研究方法
1.1 研究区概况
试验地位于广西壮族自治区凭祥市中国林业科学研究院热带林业研究中心的伏波实验场,该试验区属南亚热带季风型半湿润-湿润气候,年日照时数1218—1620 h,年平均气温20.5—21.7 ℃,年均降水量 1200—1500 mm。干湿季节明显,4—9月为雨季,10月至翌年3月为旱季,雨热同季的气候特点对植物生长十分有利。土壤为花岗岩发育的山地红壤(卢立华等,2008)。林下主要灌木有三桠苦(Evodia lepta)、水东哥(Saurauia tristyla)、杜茎山(Maesa japonica)、粗糠柴(Mallotus philippensis)、中平树(Macaranga denticulata)、九节(Psychotria rubra)、黄毛榕(Ficus esquiroliana)、潺槁木姜子(Litsea glutinosa)等,草本大多为半边旗(Pteris semipinnata)、乌毛蕨(Blechnum orientale)、扇叶铁线蕨(Adiantum flabellulatum)、芒萁(Dicranopteris dichotoma)、淡竹叶(Lophatherum gracile)等。
1.2 样地设置
试验林均为 1993年营造的杉木纯林,初植密度为2500 plant·hm-2。1999年和2003年分别进行了强度约为30%的透光伐和强度约为40%的抚育间伐,2008年实施近自然化改造,对杉木纯林进行疏伐,近自然改造林杉木保留密度为600 plant·hm-2,疏伐后在林下以 400 plant·hm-2的密度分别均匀套种红锥、大叶栎,构建杉木混交林,并设置疏伐后保留密度为1000 plant·hm-2的杉木纯林作为对照。在杉木×红锥混交林(MCC)、杉木×大叶栎混交林(MCQ)和杉木纯林(PCL)中选取自然环境条件基本一致的典型区域,各设立3块20 m×20 m的固定试验样地,同一林分中各样地间距离20 m以上,即每种林分设3个重复。2018年10—11月在固定样地进行每木调查,样地基本情况见表1。

表1 不同类型林分样地基本情况Table 1 Basic situation of different stand types on the ground
1.3 样地调查
将3种不同类型林分的20 m×20 m样地分成4个10 m×10 m的中样方,分别调查各个中样方里的乔木(DBH≥1 cm)树种种类、数量、胸径、树高和冠幅;在各个中样方(10 m×10 m)的左上角设置1个5 m×5 m的小样方,调查并记录小样方内乔木幼树幼苗(H≥30 cm,DBH<1 cm)及所有灌木种类、数量、平均高度和覆盖度;同时再设置1个1 m×1 m的微型样方,调查并记录微型样方内所有草本植物的种类、数量、平均高度和覆盖度。
1.4 土壤样品采集与分析
在每个20 m×20 m样地内按“S”形设置5个采样点,分别在0—20 cm土层采集原状土壤1 kg。将各个样地内土壤样品分别混合均匀。将每个混合样品沿自然结构轻轻掰开,并过2 mm筛除去动植物残体和小石块等,置于实验室内自然风干。风干后的土壤用磨土机粉碎后过<0.2 mm筛,测定土样的有机碳(SOC)和全氮(TN)含量,其中有机碳采用高温外热重铬酸钾氧化-分光光度法,全氮采用消煮法并使用连续流动分析仪测定(鲁如坤,2000)。
1.5 数据处理与统计分析
1.5.1 植物多样性及重要值测定
统计样方内物种组成并分析树种重要值和多样性。重要值可作为判断群落中优势树种的衡量指标(Naeem et al.,1997)。其计算公式分别为:乔木重要值 (Iv1)=(相对多度+相对显著度+相对频度)/3,灌木 (草本) 重要值 (Iv2)=(相对多度+相对盖度+相对频度)/3。植物多样性利用丰富度指数(Rrichness index,S)、香农维纳指数(Shannon-Wiener index,H′)、辛普森指数(Simpsonindex,D)、均匀度指数(Pielou index,E)进行分析(Magurran,1988)。

式中:
S——总物种数;
Pi——种的个体数占群落中总个体数的比例。
1.5.2 数据统计分析
应用 Excel 2007进行数据计算和制图,SPSS 19.0对数据进行单因素方差(One-way ANOVA)分析和Pearson相关性分析,并采用邓肯法进行多重比较(P<0.05)。
2 结果与分析
2.1 不同林分类型群落的物种组成及重要值
经调查发现(表 2),3种林分类型乔木层物种数表现为PCL>MCQ>MCC。其中,MCC物种数为10种,除杉木和红锥外,重要值超过5%的树种还有香梓楠和中平树;MCQ物种数为14种,除杉木和大叶栎外,格木、三桠苦、黄毛榕、红锥也占据一定的优势;PCL物种数为18种,除主要造林树种杉木外,黄毛榕和毛黄肉楠的重要值也都在5%以上。此外,在3种林分类型中,水东哥均有分布,但重要值均低于5%。
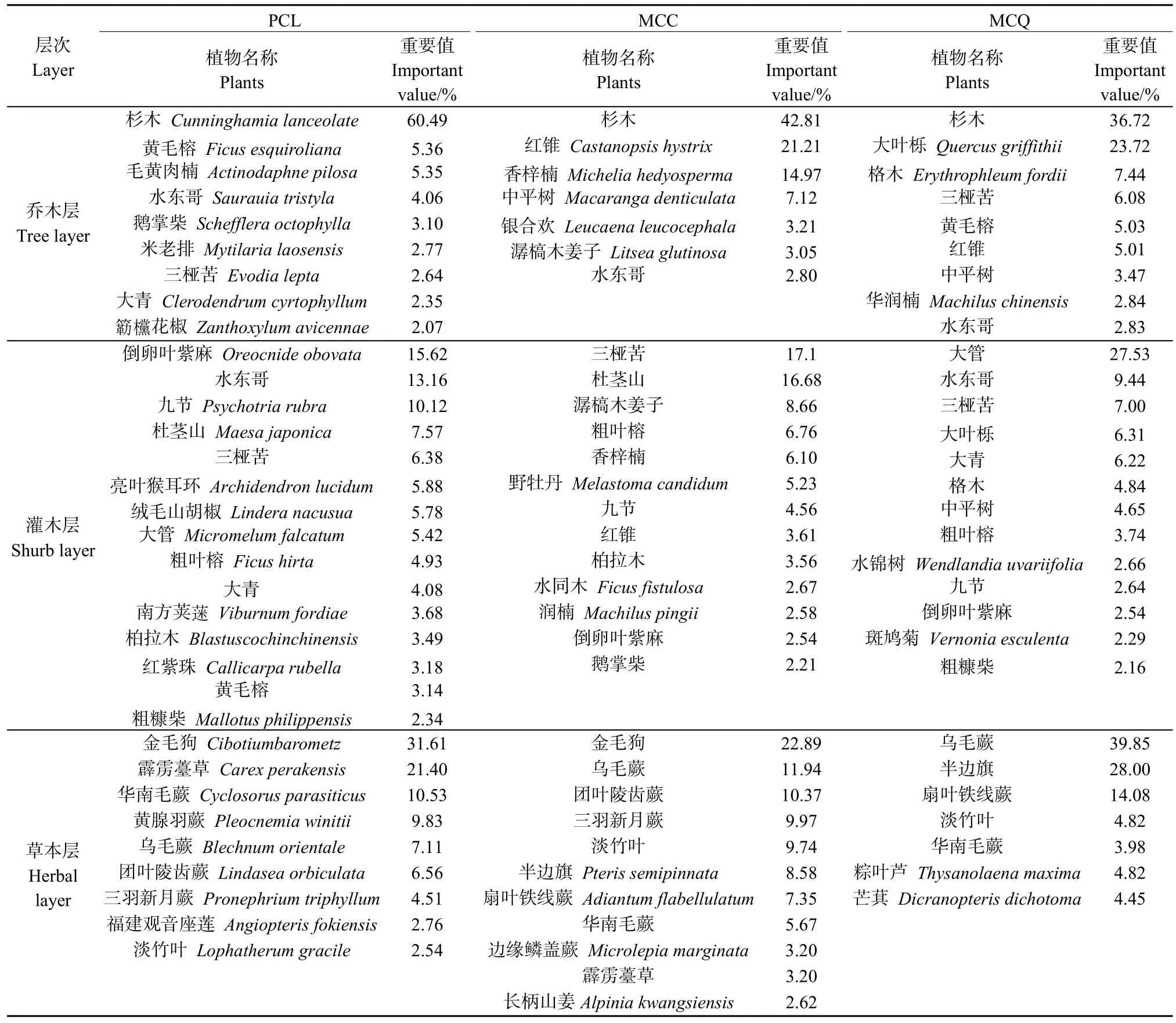
表2 不同林分类型物种组成及重要值Table 2 Species composition and important value of plants in different stand types
灌木层主要由乔木的幼树幼苗(H≥30 cm,DBH<1 cm)及丛生灌木组成。3种林分类型中灌木层物种数为 MCQ>MCC>PCL。不同林分灌木层优势种不同,其中,MCC植物物种数为23种,优势种为三桠苦、杜茎山,重要值分别为 17.10%和16.68 %;MCQ的植物物种数为24种,主要优势种为大管,重要值为27.53%,水东哥、三桠苦、中平树和大青(Clerodendrum cyrtophyllum)的重要值都在5%—10%之间;PCL的植物物种数为18种,优势种为倒卵叶紫麻、水东哥、九节,重要值分别为15.62%、13.16%、10.12%。
草本层的优势种主要为蕨类及禾本科植物。MCC草本层物种数为 13种,优势种为金毛狗(Cibotiumbarometz)、乌毛蕨、团叶陵齿蕨(Lindaseaorbiculata),重要值分别为 22.89%、11.94%、10.37%;MCQ草本层物种数为7种,优势种为乌毛蕨、半边旗、扇叶铁线蕨,重要值为 39.85%、28.00%、14.08%;PCL草本层物种数为11种,优势种为金毛狗、霹雳薹草(Carex perakensis)、华南毛蕨(Cyclosorus parasiticus),重要值为31.61%、21.40%、10.53%。
2.2 不同林分类型植物多样性
对MCC、MCQ、PCL林分乔木层、灌木层、草本层以及群落的植物多样性指数进行分析发现(图1),乔木层Shannon-Wiener指数、Simpson指数和Pielou均匀度指数表现为MCC和MCQ显著高于PCL(P<0.05),MCC和MCQ的丰富度指数与PCL差异不显著(P>0.05)。在3种林分类型中,MCQ的丰富度指数、Shannon-Wiener指数和Simpson指数最高,Pielou均匀度指数最高的是MCC。草本层、灌木层和群落中,MCC、MCQ的植物多样性指数均与PCL无显著差异。MCC的3种群落层次植物多样性指数均无显著差异;在MCQ中,丰富度、Shannon-Wiener和Simpson指数表现为灌木层>乔木层>草本层,且差异显著,Pielou均匀度指数以灌木层最高,且灌木层和草本层显著高于乔木层;PCL中,植物多样性指数均以灌木层最高,灌木和乔木层的丰富度指数显著高于草本层,灌木层Shannon-Wiener和Simpson指数与乔木层、草本层存在显著差异,乔木层Pielou均匀度指数显著低于灌木和草本层。
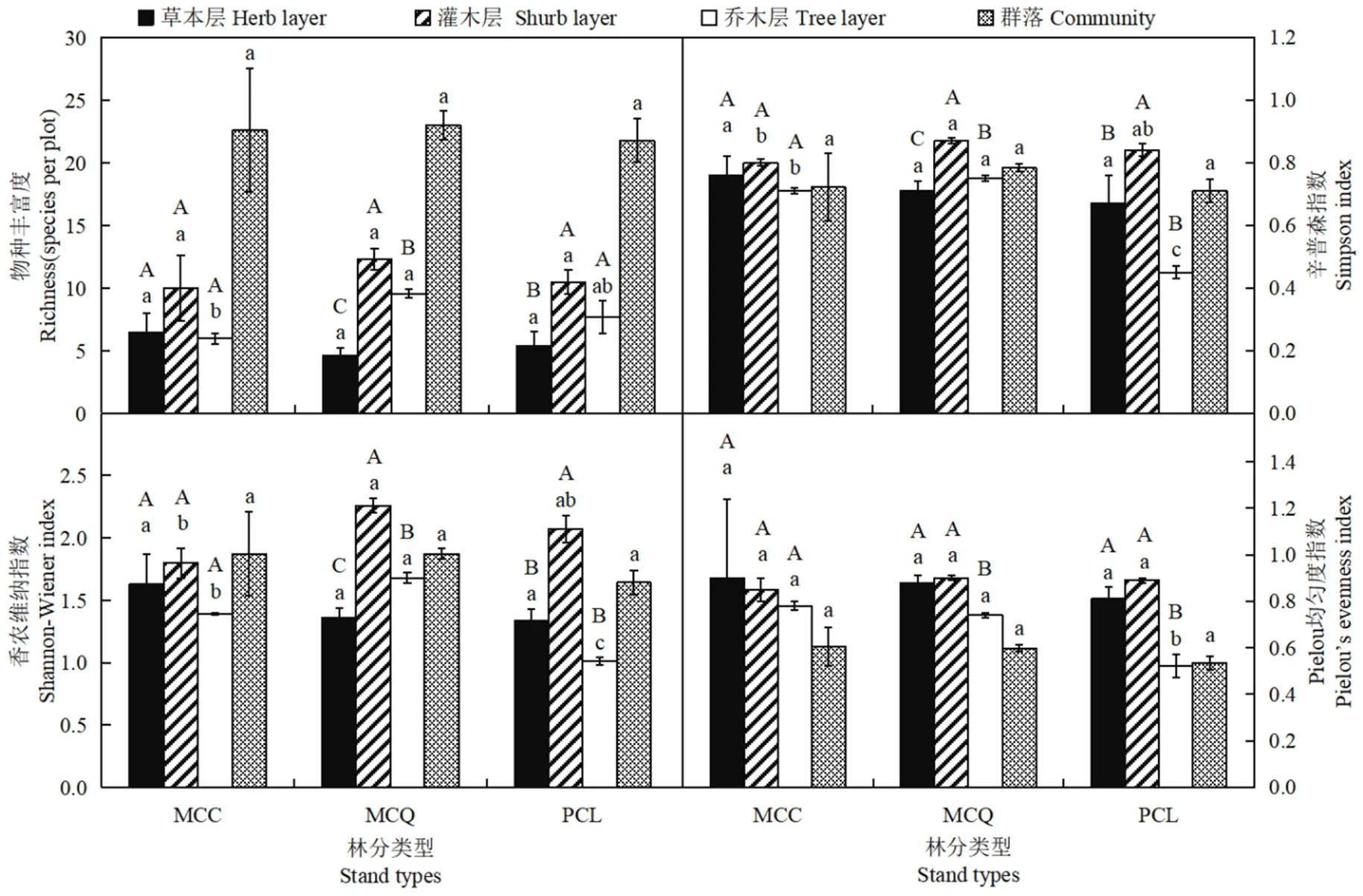
图1 不同林分类型植物多样性指数比较Figure 1 Comparative study on the indexes of plant species diversity in different forest types
2.3 不同林分类型土壤碳氮含量的变化
不同林分土壤有机碳含量以MCQ最高,且显著高于 PCL(P<0.05),MCC土壤有机碳含量高于PCL,但差异不显著(P>0.05);全氮含量由高到低依次为 MCQ、PCL、MCC;碳氮比由高到低依次为MCC、PCL、MCQ;MCC、MCQ的全氮含量及碳氮比均与PCL无显著差异(图2)。

图2 不同林分类型土壤有机碳、全氮含量以及碳氮比Figure 2 Soil organic carbon and total nitrogen content of different stand types
2.4 植物多样性与土壤碳氮含量的相关分析
对3种林分植物多样性指数与土壤碳、氮含量进行相关性分析(表 3),发现乔木层Shannon-Wiener指数与土壤有机碳含量的正相关关系达到显著水平(P<0.05);灌木层Simpson指数与土壤有机碳含量表现为显著正相关关系,Shannon-Wiener指数、Simpson指数与全氮含量呈极显著正相关(P<0.01);丰富度指数及与碳氮比呈显著负相关,Shannon-Wiener指数与碳氮比为极显著负相关,草本层和群落的植物多样性指数均与土壤有机碳、全氮含量及碳氮比无显著相关关系(P>0.05)。

表3 植物多样性指数与土壤碳氮含量相关分析Table 3 Correlation analysis between plant species diversity indices of tree and chemical properties
3 讨论
群落的物种多样性是群落结构和功能复杂性的度量,能较好地反映群落的稳定性(高贤明等,2001)。研究表明,营造针阔混交林,可提升林地空间资源的利用率,增强树种多样性,进而增强林地抗逆性,提高生态系统的稳定性(Jactel et al.,2018)。本研究中,从林分整体群落来看,植物多样性指数以MCQ最高,MCC次之,PCL最低。可见,营造混交林有利于增强植物群落物种多样性,提高抵御不良环境或外部干扰的能力(黄冬梅,2017;康永祥等,2010)。
乔木层两种混交林香浓维纳指数、辛普森指数、Pielou均匀度指数均显著高于PCL(P>0.05),丰富度指数无显著差异(P<0.05),说明与混交林相比,PCL乔木层物种组成较为单一,分布不均。混交林灌木层、草本层的植物多样性指数与PCL的差异不显著,与李海霞等(2010)、冯健等(2021)研究结果相似。灌木层植物多样指数以MCQ最高,原因是混交树种大叶栎树冠浓密,林下透光度较小,更有利于耐荫灌木及乔木幼树幼苗的生长。草本层香浓维纳指数、辛普森指数、Pielou均匀度指数以PCL最低,这是由于杉木纯林地表针叶枯落物较多且难以分解,导致土壤养分下降,使一些草本植物难以在杉木纯林林下定居(王玉杰等,2020)。草本层均以禾本科和蕨类植物为主要优势种,一方面,禾本科草本植物起源于热带森林或林缘开放的生境,对环境适应能力强;另一方面,林下接受光照的面积有限,促进了蕨类等耐荫植物的生长(张涵丹等,2021)。在3种林分类型中,不同群落层次植物多样性指数以灌木层最高,因为灌木层的生态环境分化程度大于乔木层和草本层,不仅具有真正的灌木种类,还包括乔木的幼树幼苗(吴迪等,2017)。此外,杉木纯林及其混交林优势种作用明显(杉木、红锥、大叶栎),其他树种生长处于劣势,均匀度指数较低(吴迪等,2017),致使乔木层的均匀度指数低于灌木层和草本层。
土壤有机碳含量的变化与树种组成、植被覆盖度、凋落物累积与分解程度、土壤微生物、根系在土壤中的分布等因素有关(董云中等,2014)。本研究中PCL有机碳含量低于MCC和MCQ,说明营造杉阔混交林有利于提高土壤有机碳含量,与目前大部分研究结果一致(黄宇等,2005;杨玉盛等,2003;郭家新,2008)。在气候环境基本一致的情况下,植被类型、产量和植物残体的分解过程是有机质的重要控制因子(Ganuza et al.,2003),针叶林凋落物中含有大量难分解的有机质,影响了土壤养分的归还速率,且针叶林林下枯落物层较厚,腐殖质层较薄,既不利于枯落物的分解,又阻挡种子插入土壤和植被发育,导致植被生长较差;而针阔混交林植被组成丰富,腐殖质层较厚,阔叶树种往往具有更大的凋落物产量和分解速率,使其养分归还量和归还速率提高,从而提高土壤中有机碳的含量(孙浩等,2020)。两种混交林土壤全氮含量与PCL差异不显著,与前人的研究结果相似(肖石红等,2018;冯健等,2021)。由于MCQ中含有固氮树种——格木,通过自身固氮减少对土壤的氮吸收,并可能将固定的氮素转移到邻近植物,降低土壤氮消耗(Yao et al.,2019),而且格木具有一定的优势(在乔木层和灌木层的重要值分别为为7.44%和4.84%,其在乔木层的重要值仅次于杉木及大叶栎),故土壤全氮含量最高。3种林分类型碳氮比的变化范围为 14.28—17.51,均高于中国土壤碳氮比均值11.9和世界土壤碳氮比均值13.33(郭冬艳,2013),可能是存在氮受限,影响土壤微生物分解能力(曹小玉等,2019)。
本研究中,土壤有机碳含量与乔木、灌木层的多样性指数呈正相关关系,且与 Shannon-Wiener指数及灌木层Simpson指数的相关性达到显著水平(P<0.05);全氮含量与乔木层表现为正相关关系,与灌木层Shannon-Wiener指数、Simpson指数呈极显著正相关(P<0.01),灌木层丰富度指数和Shannon-Wiener指数与碳氮比呈显著负相关关系,说明全氮、有机碳与杉木林植物多样性相互作用、相互影响。因此,适当施加氮肥,可促进乔木、灌木层植物多样性的提高(舒韦维等,2021)。地上凋落物和地下根系的分解是土壤养分的重要来源(谢君毅等,2021)。乔木、灌木层植物多样性指数与土壤碳、氮含量相关性高于草本层,相较于草本,灌木和乔木的根系更为发达,凋落物更多,且该研究区域相对较开阔,呈现偏阳性的环境,有利于动、植物残体在微生物作用下分解(程瑞梅等,2010),故乔木、灌木层对土壤有机碳、氮含量的影响更大。群落植物多样性指数与土壤有机碳、全氮含量及碳氮比无显著关系(P>0.05),因为植物生长需要消耗、利用土壤中的有机碳和全氮,而植物的调落物分解以及根系分泌小分子物质会改善土壤的有机碳和全氮含量,导致土壤中碳、氮含量变化与植物多样性关系不显著。此外,随着人工林系统的生长发育,及林下植被群落的不断演替更新,植被层对养分的消耗及改良作用之间的关系变得更加复杂,也是土壤碳、氮含量与植被多样性相关性不显著的主要原因之一(王春香,2014)。
4 结论
杉木与红锥、大叶栎混交都不同程度地提高了林分植物多样性,有利于杉木人工林可持续发展,且 MCQ植物多样性最高,抵御不良环境和外部干扰的能力最强(表 1)。两种混交林土壤有机碳均高于PCL,以MCQ最高。乔木、灌木层植物多样性指数与土壤有机碳、氮含量及碳氮比的相关性表明有机碳、氮与杉木林植物多样性相互作用、相互影响。乔木、灌木层植物多样性指数与土壤碳、氮含量相关性高于草本层,群落植物多样性指数与土壤有机碳、全氮含量及碳氮比无显著关系,可见植物多样性及土壤碳、氮含量不仅与混交树种的选择有关,同时也可能受其他因素影响。
