钙蛋白酶系统介导的骨骼肌蛋白降解
2022-05-23杨飞云齐仁立
林 俊,刘 鑫,杨飞云,齐仁立*
(1.重庆市农业学校,重庆九龙坡 401329;2.重庆市畜牧科学院,重庆荣昌 402460;3.西南大学动物科技学院,重庆荣昌 402460)
成年动物的肌肉蛋白长期处于合成与降解的动态平衡。促进肌肉蛋白质合成,减少蛋白降解是维持正常肌肉体积和生理功能的主要途径。钙蛋白酶系统在骨骼肌细胞的蛋白降解和代谢周转中发挥重要调控作用,影响了肌肉发育和生理稳态,也对动物宰后的肉品嫩度有重要影响(王胜军等,2016;Tipton和 Wolfe,2001)。
1 钙蛋白酶系统的组成
钙蛋白酶系统由多种钙蛋白酶(Calpain,CAPN)和钙蛋白酶抑制蛋白(Calpastatin,CAST)组成。CAPN是一类胱氨酸蛋白酶,可催化不同类型的蛋白底物水解(Hyatt等,2020)。CAPN的活化和蛋白水解功能发挥都依赖于细胞内Ca2+浓度。最主要的CAPN可被分为μ-钙蛋白酶(1-钙蛋白酶,CAPN 1)和m-钙蛋白酶(2-钙蛋白酶,CAPN 2),它们分别需要微摩尔和毫摩尔Ca2+的激活,在不同组织都有广泛表达分布(Huang等,1998)。此外,被发现了还有13种CAPN同工酶,它们参与了众多钙离子调节的细胞活动,如信号传导、细胞增殖、细胞分化等(Du等,2019;Leloup等,2011;张勇等,2007)。
CAST是生物内源性专一抑制CAPN活性的蛋白质,位于细胞的胞质溶液中,与CAPN共存(Du等,2019;张勇和邓科,2011)。CAPN与Ca2+结合后会产生蛋白结构的变化,CAST能准确识别这种变化并与之结合,抑制CAPN的催化活性(Wendt等,2004)。
骨骼肌组织中主要存在CAPN 1、CAPN 2、CAPN 3和 CAPN 10(Hata等,2020;许梓荣,2002)。CAPN 3在骨骼肌中特异性表达,在肌原纤维形成方面具有重要作用。CAPN 3基因的突变会引起肌肉发育不健全和多种肌营养不良症,特别是肢带肌营养不良2A症也被称为钙蛋白酶病(Lasa-Elgarresta等,2019)。还有研究指出,CAPN 3可能参与了骨骼肌细胞的生成和细胞凋亡调控(Dargelos等,2002)。
2 钙蛋白酶家族结构和激活机制
2.1 钙蛋白酶的结构 多数CAPN都是异源二聚体结构,由序列不同的80kda催化亚基(大亚基)和序列相同的30kda调节亚基(小亚基)组成。典型大亚基包括4个结构域(Ⅰ-Ⅳ),而小亚基包括2个(Ⅴ、Ⅵ)结构域(图1)(许梓荣等,2002;Huang等,1998)。
位于氨基端的结构域Ⅰ在CAPN活化后会发生自溶,对维持CAPN空间结构及对酶活化所需的空间构象变化发挥重要作用。结构域Ⅱ是蛋白酶水解活性的关键部位,由2个子域组成(Ⅱa和Ⅱb)。构象发生变化后,结构子域Ⅱa上的氨基酸残基Cys-105和结构子域Ⅱb上的His-262和Asn-286会接近,并形成催化活性中心(催化三聚体),是蛋白酶的主要水解活性位点。结构域Ⅲ上有1个由8~10个酸性氨基酸残基组成的酸性环,是结构域Ⅳ和Ⅵ的调节中心。酸性氨基酸残基的数量越少,Ca2+结合能力越弱。位于大亚基羧基端的结构域Ⅳ和小亚基上同样位于羧基端的结构域Ⅵ是钙离子结合域,都含有5个EF-手型钙离子连接序列。小亚基的另1个结构域Ⅴ位于氨基末端且富含甘氨酸(约60%左右),具有强疏水作用,对维持大亚基的正确空间构型不可或缺(Reverter等,2001)。
CAPN 3是由单体蛋白组成,增加了3个独立序列结构单元:NS、IS1和IS2,分子量达到 94KDa(图 1)(Hata等,2020;Hiroyuki和Yasuko,2012)。CAST 并不能有效结合和抑制CAPN 3,相反还是CAPN 3的水解底物(齐曾鑫等,2013)。
2.2 钙蛋白酶抑制蛋白的结构 CAST含有5个结构区域,包括有4个重复结构单位(I、Ⅱ、Ⅲ、Ⅳ)和1个L结构区。位于氨基末端的L区是Ca2+的通道调节区,而每个重复结构单位又包含3个序列保守区(A区、B区、C区),是CAPN的抑制区(图 1);杨雪梅等,2007;Wendt等,2004)。
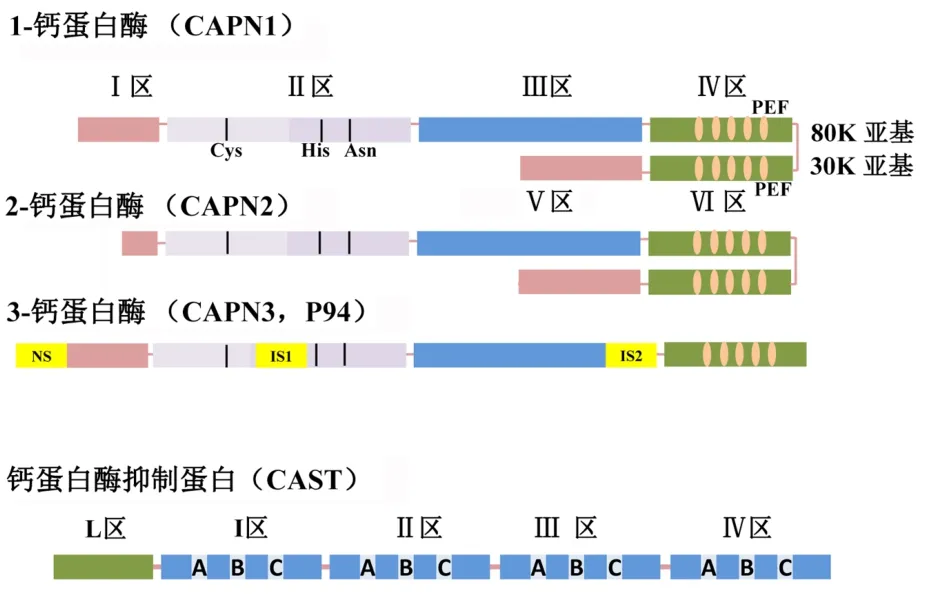
图1 钙蛋白酶和钙蛋白酶抑制蛋白的结构示意图
2.3 钙蛋白酶的激活机制 Moldveanu等(2002)研究发现,细胞中游离的Ca2+浓度不足时,CAPN的子结构域IIa和IIb之间的裂缝由于结构域间的相互吸引而被封闭。结构域IIa上和IIb上构成酶活性中心的3个氨基酸残基Cys105、His262和Asn286彼此分离。当细胞中的Ca2+浓度快速上升到一定程度后会结合并诱发CAPN的结构域构象发生改变,大亚基上的结构域I发生自溶,导致结构域IIa和IIb之间的裂缝封闭移除,氨基酸残基相互接近并形成催化三聚体(酶活中心),CAPN开始发挥蛋白水解催化活性(Nakagawa等,2001)。
3 钙蛋白酶调控的骨骼肌蛋白降解
3.1 骨骼肌蛋白降解的不同途径 人和动物的骨骼肌细胞中肌原纤维蛋白占肌肉蛋白总量的55%~60%,其中肌球蛋白和肌动蛋白是最主要的两种(Goll等,2008)。在肌细胞内存在3种主要的蛋白降解途径:溶酶体途径、钙蛋白酶途径和泛素-蛋白酶体途径(Neti等,2009)。由于哺乳动物的肌原纤维尺寸较大(粗1~3 μm,长 1~ 40 μm),无法进入溶酶体(直径通常0.2~0.8 μm)进行分解。而泛素-蛋白酶体只能降解单个肌原纤维蛋白,当肌原纤维交联时就不能被降解(Solomon,1996)。CAPN可能是骨骼肌蛋白降解的发起者和主要限速步骤,它首先催化肌细胞中大量肌原纤维解联、降解并释放出粗肌丝和细肌丝,然后肌丝及其组分蛋白又被其他蛋白水解途径单独或协同分解。
3.2 钙蛋白酶系统介导的骨骼肌蛋白降解 CAPN在骨骼肌细胞中催化的蛋白降解通常只针对肌原纤维。CAPN参与并调控了肌原纤维降解并释放出肌丝的过程(Neti等,2009;赵红艳,2008)。肌细胞中的CAPN主要存在于肌原纤维Z线(z-line)附近。CAPN活化后,首先会迅速分解Z线附近的肌联蛋白和伴肌动蛋白,然后继续分解中间丝蛋白及肌纤维膜上的索蛋白,随后α-辅肌动蛋白等被释放出来,直至Z线消失(Sorichter等,1999)。分解下来的肌联蛋白、肌球蛋白等进一步降解还需要泛素蛋白酶体、溶酶体、线粒体蛋白酶等其他水解酶的参与。
3.3 钙蛋白酶影响动物肉品嫩度 CAPN催化的肌原纤维降解也是动物肌肉宰后嫩度提高和肉品成熟的主要原因(Huang等,2019;Huff 等,2010;王媛媛等,2006)。正常状态下,动物肌细胞中Ca2+浓度较低,不足以激活钙蛋白酶,而在动物处死后,肌浆网上的Ca2+大量外泄,CAPN(主要是CAPN1)被激活,启动肌纤维蛋白降解。动物死后肌肉中CAPN 1的活性大概在24h后基本消失。
4 营养对钙蛋白酶系统的影响
4.1 营养水平对钙蛋白酶系统的影响 日粮营养素或营养水平变化会导致动物肌肉生理特性的改变与钙蛋白酶系统存在紧密的联系。一些研究认为,肌肉中的CAST更容易被日粮营养所影响。Helman等(2003)犬类研究表明,日粮营养水平仅对CAST产生明显的影响,但是对CAPN的催化活性影响不显著。相似地,Du等(2004)在牛上的试验结果表明限饲并不影响肌肉中CAPN的活性,但显著降低了CAST的调控影响,导致肌肉块减小。但还有一些研究指出,CAPN也会受到营养水平的影响。Ilian等(1992)研究发现,长期禁食使兔的肌肉蛋白降解程度增加2倍,CAPN 1,CAPN 2和CAST的mRNA水平分别提高4.2倍、4.5倍和3.2倍,这使肌肉体积减小并导致骨骼肌萎缩。Tang等(2010)在猪上的研究指出,适度降低营养水平会增加育肥猪肉品嫩度,增加肌肉中CAPN的mRNA水平。
4.2 功能性营养素对钙蛋白酶系统的影响 一些功能性营养物质也可影响和调控钙蛋白酶系统。使用富含亮氨酸(3%)的食物饲喂带有癌症肿瘤的的妊娠母鼠,能强化其子代小鼠肌肉的蛋白周转,保持肌肉组织中CAPN的活性(Miyaguti等,2019)。Choi等(2012)给小鼠补充维生素D3三周发现,背最长肌中的脂肪含量明显增加,并提高肌肉中CAPN 1含量,减少CAST含量。给大鼠补充大豆提取蛋白可显著提高其运动能力,对抗运动导致腿部肌肉损伤,抑制CAPN 1和CAPN 2的激活,提高CAST的蛋白活性(Nikawa等,2002)。Tang等(2008)发现,在屠宰前 5 d给商品猪补充天冬氨酸镁(2000 mg/kg)可显著提高宰后猪肉中CAST含量,减轻运输和屠宰应激对肉嫩度的负面影响。显然,研究并建立以钙蛋白酶系统为靶点的精准营养调控技术可提高动物产肉性状和改善肉品质。
5 小结
钙蛋白酶系统在肌细胞蛋白降解过程中扮演了重要的调控角色,影响肌肉生长与生理稳态,且对动物肉品质具有重要影响。研究并利用精准营养调控技术靶向调控CAPN和CAST的表达与活性,可能是未来提高动物产肉量和改善肉品质的一种新的调控措施。
