小麦TaKLU基因调控植物株型的研究
2022-05-23周梦蝶李梦瑶吴林楠郭利建
周梦蝶,李梦瑶,吴林楠,郭利建,王 倩,2,马 猛,2
(1.西北农林科技大学生命科学学院,陕西杨凌 712100;2.旱区作物逆境分子生物学国家重点实验室,陕西杨凌712100)
作物株型受株高、分枝数目、分枝角度等因素的影响,是决定作物产量的关键因素之一。其中,分枝数目是影响株型的主要因素,一般与株高呈负相关。在表现形式上,作物通常有两种分枝状态,分别为分枝少而株高高与分枝多而株高矮,表现为顶端优势的加强与弱化。因此,作物的分枝数目及株型在很大程度上受到顶端优势的影响,进而决定作物产量形成。不同作物对顶端优势的利用存在很大差异。如玉米通过加强顶端优势、减少分枝来增加产量,而水稻则通过增加分枝数目、弱化顶端优势来提高产量。因此,改良作物株型是成功培育高产作物新品种的关键途径之一,但调控小麦株型的分子机制尚不明了。
基因家族是一类只存在于植物体内的P450亚家族,对作物的性状改良育种具有重要的意义。前人对家族基因的研究多集中在单个器官发育上,尤其是对籽粒大小的调控。拟南芥中,5()基因正调控生殖器官大小,过表达该基因会导致花序和种子增大;和双突变体表现为育性降低和籽粒变小,导致产量降低。水稻中,()基因通过调节胚和胚乳的比例,影响籽粒大小和产量;()基因是拟南芥基因的直系同源基因,通过调节叶原基细胞的增殖影响叶片发育,突变体表现为叶片起始速率加快、叶片变小。玉米中,()基因调控器官大小、株高和产量,并且涉及生长素代谢,暗示基因的调控作用可能不仅仅局限于单个器官,甚至于影响整个植株的发育。目前研究者普遍认为,家族基因以非细胞自主的方式,通过调控下游的促生因子来促进植株发育,即类似于植物激素的作用模式。
本课题组前期研究发现,小麦基因家族包含四个成员,分别为、(以下用表示)、和,其中,和基因对籽粒大小和生殖器官发育具有显著调控作用。然而植物中关于家族基因调控株型的研究鲜有报道。本研究通过综合调查小麦对拟南芥分枝数目、顶端优势的影响,初步明确基因与株型的关系,以期为进一步揭示基因调控植物株型的功能和机制奠定基础,为小麦株型改良育种提供参考。
1 材料与方法
1.1 试验材料
本研究所用小麦材料为小偃6号,拟南芥材料为哥伦比亚野生型(Columbia-0)。拟南芥功能缺失突变体(Salk_024697C)购自Biological Resource Center(https://abrc.osu.edu/)。
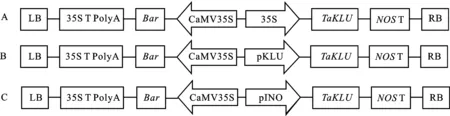
LB:左边界;35S T PolyA:35S(CaMV35S)终止子;Bar:抗除草剂基因;CaMV35S/35S:花椰菜花叶病毒35S启动子;pKLU:拟南芥KLU基因启动子;pINO:拟南芥胚珠特异启动子;NOS T:NOS基因终止子;RB:右边界。
1.2 试验方法
1.2.1 阳性植株的筛选
利用RNA提取试剂盒(百泰克,中国)提取小麦苗期叶片总RNA,并用反转录试剂盒(艾科瑞,中国)反转录为cDNA。利用特异性引物TaKLU-cDNA-F/R对基因编码区进行扩增。选用pCAMBIA3301表达载体(含有35S启动子和Basta除草剂抗性)进行载体构建,通过I和II限制性酶切位点将基因的编码区序列连接到pCAMBIA3301载体的35S启动子,获得35S::过表达载体(图1A),并将其导入野生型拟南芥中,获得3个转基因拟南芥株系(35S::-1、35S::-3和35S::-5)。利用特异性引物pKLU-F/R扩增拟南芥基因(AT1G13710)上游2 229 bp的启动子序列,通过R I和I限制性酶切位点将扩增得到的pKLU启动子序列替换表达载体p35S::中的35S启动子序列,获得表达载体pKLU::(图1B);利用特异性引物pINO-F/R扩增拟南芥基因 (AT1G23420)启动子序列,通过R I和I限制性酶切位点将扩增得到的pINO启动子序列替换35S::中的35S启动子序列,获得表达载体pINO::(图1C)。具体引物序列见表1。上述构建的表达载体分别转入农杆菌EHA105中,并通过花序侵染法导入拟南芥突变体中进行回补试验,获得pKLU::;、pINO::;和35S::;回补转基因株系,以验证基因在调控株型和器官发育方面的功能。

表1 本研究使用的引物信息Table 1 Primers used in the study
将上述T代转基因拟南芥株系、野生型以及突变体材料的种子种植在蛭石和营养土的混合 (1∶2)土壤中,在温室(16 h光照/8 h黑暗)中培养,培养温度为22 ℃,直至全部角果成熟。
基于阳性转基因拟南芥能够获得除草剂Basta(含20%草铵膦)的抗性,本研究通过对拟南芥叶片(2片真叶时)喷施200 mg·L的Basta溶液对阳性植株进行初步筛选;用CTAB法提取转基因阳性植株、野生型和突变体的叶片DNA,以野生型和突变体的叶片DNA为对照,进一步利用TaKLU-F/R引物(表1)对Basta筛选的阳性植株进行PCR鉴定;最后综合Basta筛选和PCR鉴定结果,确定下一步试验的阳性植株。PCR反应体系为20 μL,包括2×Rapid Tap Master Mix(诺唯赞,中国)10 μL,模板DNA(60 ng·μL)1 μL,上下游引物(10 μmol·μL)各0.5 μL,HO 8 μL。PCR反应程序:95 ℃ 3 min;95 ℃ 15 s,68 ℃ 15 s,72 ℃ 20 s,29 个循环;72 ℃ 5 min。
为了进一步检测回补转基因拟南芥中的基因是否正常表达,收集转基因拟南芥和对照植株组织的样品[其中pINO启动子驱动基因表达的转基因植株以及野生型和突变体在生殖生长期(发芽后6周)取花序作为检测样品,其他启动子驱动基因表达的转基因拟南芥在发芽后3周取叶片作为检测样品],利用RNA提取试剂盒(百泰克,中国)提取上述样品总RNA,并用反转录试剂盒(艾科瑞,中国)反转录为cDNA;以TaKLU-RT-F/R为引物,利用qRT-PCR检测基因的转录水平。以cDNA为模板,PCR反应体系同上。PCR反应程序:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,35个循环。
1.2.2 转基因拟南芥的形态学观察
以野生拟南芥为对照,对35S::、pKLU::;、35S::;和pINO::;转基因株系的分枝数目和顶端优势进行观察,并在萌发后10周统计不同株系的总分枝数目,用数码相机记录表型,试验重复3次。
1.2.3 转基因拟南芥的产量性状统计
为了测量准确,一个小盆种一株拟南芥。待野生型、35S::转基因植株和突变体植株开始有角果成熟时,以单株为单位,每隔1 d收集一次成熟且干黄的角果,直至所有角果成熟。最后将所有种子置于37 ℃烘箱中放置一周,完全干燥后用于测定单株产量。
1.3 数据处理
用Excel 2013对数据进行平均值、检验和标准误的分析。
2 结果与分析
2.1 转基因拟南芥的阳性鉴定结果
T代转基因拟南芥的种子在混合营养土中萌发后,经过Basta溶液喷施筛选后,挑选阳性幼苗移栽到小盆中进行下一步的检测和观察。待发芽后3周,提取35S::、pINO::;、pKLU::;、35S::;转基因拟南芥以及野生型和突变体的叶片DNA,并用特异性引物TaKLU-F/R进行进一步的阳性鉴定。结果(图2A)显示,具有Basta抗性的转基因拟南芥均含有外源目的基因。
为更进一步检测外源目的基因在拟南芥中的表达水平,在发芽后3周收集35S::、pKLU::;和35S::;转基因拟南芥以及野生型和突变体的叶片,在发芽后6周收集pINO::;转基因拟南芥以及野生型和突变体的花序,提取样品的总RNA,并利用qRT-PCR检测目的基因的表达模式。结果(图2B和2C)显示,35S::、pKLU::;、35S::;和pINO::;转基因拟南芥中均能检测到外源目的基因的表达,而野生型和突变体中未检测到基因的表达,各植株中内参基因的表达模式相拟,表明上述转基因植株均为阳性植株,可用于后续的表型观察。
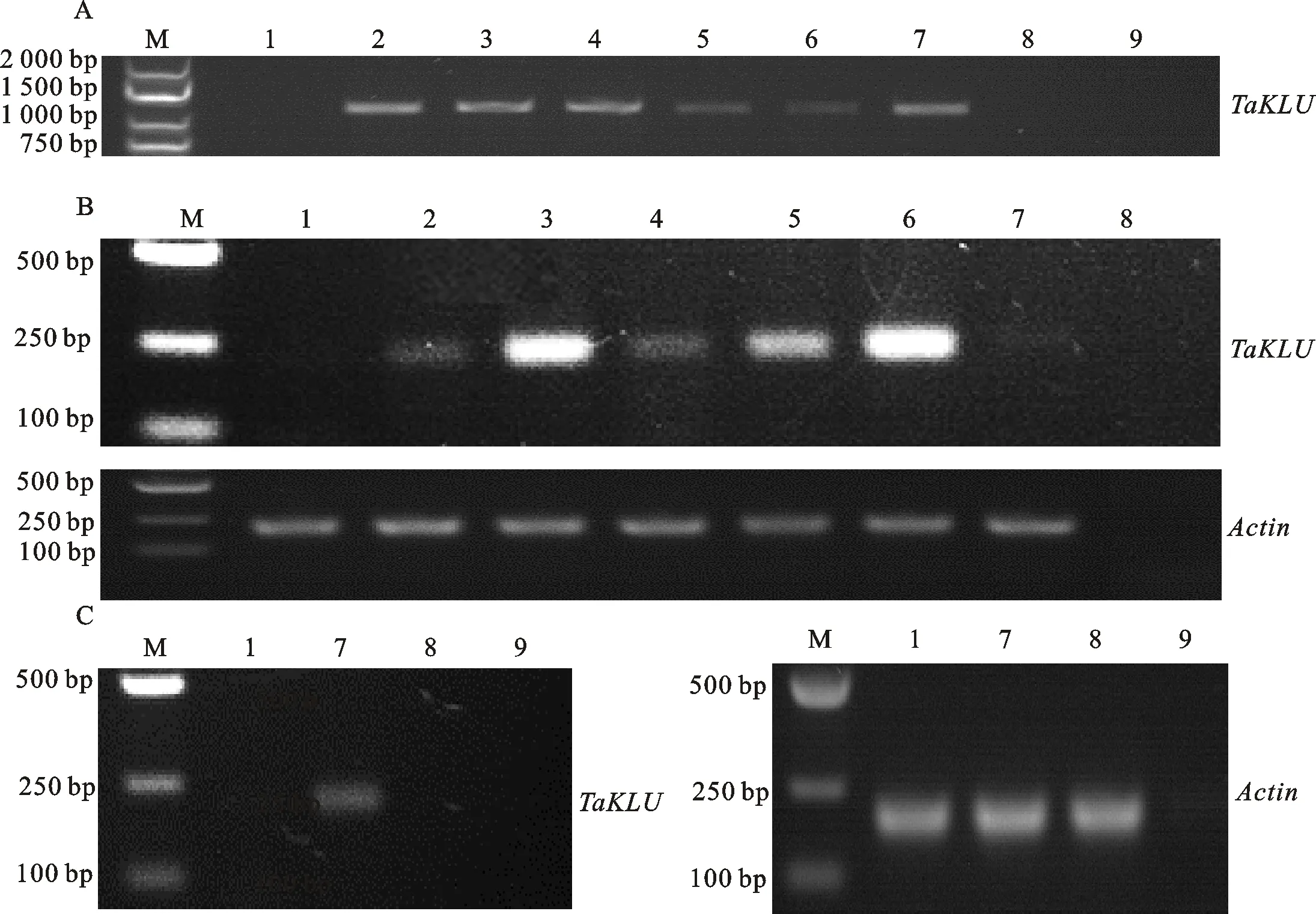
A:不同株系中外源目的基因TaKLU的PCR检测;B:qRT-PCR检测不同株系叶片中TaKLU的表达;C:qRT-PCR检测不同株系花序中TaKLU的表达;M:250 bp DNA Ladder Marker;1:野生型拟南芥;2:35S::TaKLU-1;3:35S::TaKLU-3;4:35S::TaKLU-5;5:pKLU::TaKLU;klu;6:35S::TaKLU;klu;7:pINO::TaKLU;klu;8:klu突变体;9:水;Actin:内参基因。
2.2 KLU基因缺失对拟南芥形态的影响
结果(图3)发现,与野生型拟南芥相比,突变体株高降低,花序和叶片变小,但分枝数目显著增加,增幅约为70%。此外,突变体主茎与分枝的高度几乎无明显差异,表现为顶端优势的弱化。因此,推测基因能够调控株型,影响植株的顶端优势。
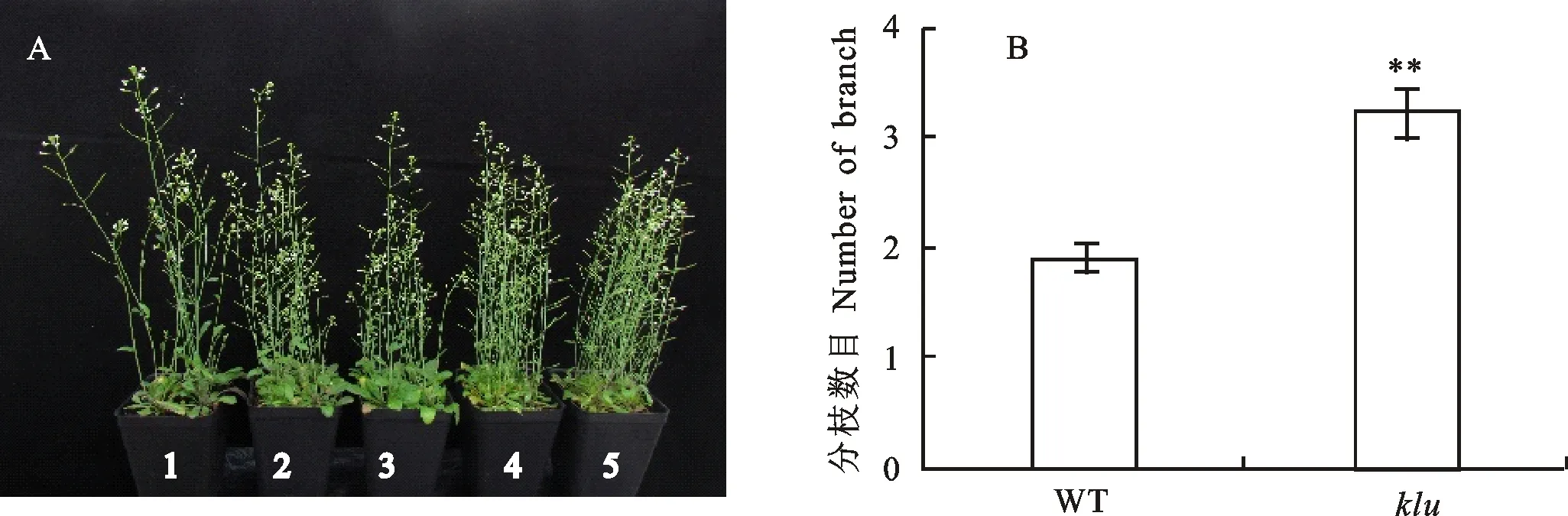
A:野生型拟南芥与klu突变体的植株形态比较;1:野生型;2~5:klu突变体。B:野生型拟南芥与klu突变体的分枝数目统计(n>18);WT:野生型;klu:klu突变体。图4和图5同。**表示野生型拟南芥与klu突变体的分枝数目在0.01水平上差异显著。
2.3 TaKLU基因过表达对拟南芥顶端优势的 影响
对野生型拟南芥和35S::转基因拟南芥(以株系35S::-1为研究对象)的株型进行比较,结果(图4A)发现,35S::转基因拟南芥的株高明显高于野生型拟南芥,且主茎远远高于分枝,表现为顶端优势的加强。此外,基因过表达还会抑制分枝的产生,即分枝数目明显减少(图4B)。进一步比较野生型拟南芥、35S::转基因拟南芥和突变体的总分枝数目,结果显示,与野生型拟南芥相比,35S::转基因拟南芥的分枝数目显著降低(除株系35S::-3与野生型差异不显著),降幅约为14%~34%,进一步说明了基因过表达可使顶端优势加强(图4B和4C)。综合以上结果表明,过表达小麦基因能够增强拟南芥的顶端优势,减少拟南芥的分枝数目。
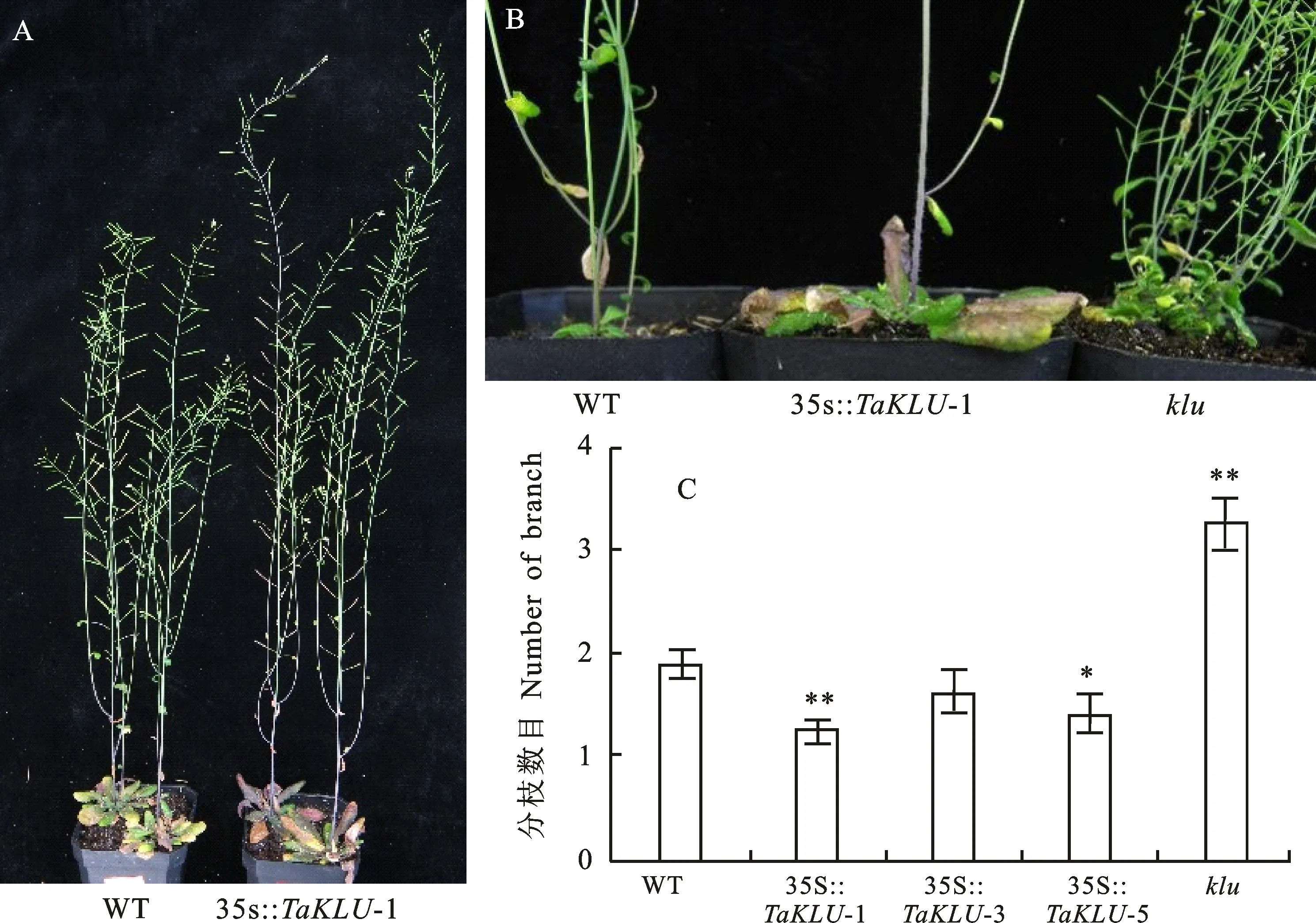
A:TaKLU基因过表达对拟南芥顶端优势的影响;B和C:不同株系的分枝情况(B)以及分枝数目的统计(C,n>17)。*和**分别表示该株系与野生型拟南芥的分枝数目在0.05和0.01水平上差异显著。
2.4 不同启动子驱动小麦TaKLU回补klu突变体后拟南芥的形态
鉴于小麦基因过表达和拟南芥基因功能缺失后,拟南芥的株型呈相反的效应,即分别表现为顶端优势的加强和减弱,推测基因可能在调控植物株型方面功能保守。为了验证这个推测,利用不同启动模式的启动子回补突变体,获得拟南芥pKLU::;、35S::;、pINO::;回补株系,并以野生型拟南芥和突变体为对照进行表型观察。结果(图5)显示,由拟南芥基因自身启动子pKLU驱动小麦基因在突变体中表达,能够恢复突变体的表型,表现为pKLU::;回补株系的株高、分枝数目、叶片和花序大小与野生型拟南芥无明显差异。与突变体相比,由强启动子35S和特异启动子pINO驱动基因在突变体中表达的35S::;和pINO::;回补株系均表现为株高明显升高,与野生型拟南芥相比,分枝数目有所增加,分别为野生型的1.4和2.6倍,其中pINO::;回补株系与野生型拟南芥之间的分枝数目差异达到极显著水平。所有基因回补株系的顶端优势都得到了加强,其中pINO::;回补株系的顶端优势加强的效应最为显著,其主分枝较其他分枝明显伸长(图5A),这也进一步支持了过表达基因能够加强顶端优势的结论。以上结果表明,和基因在调控株型上功能保守,均具有正调控植株顶端优势的功能。
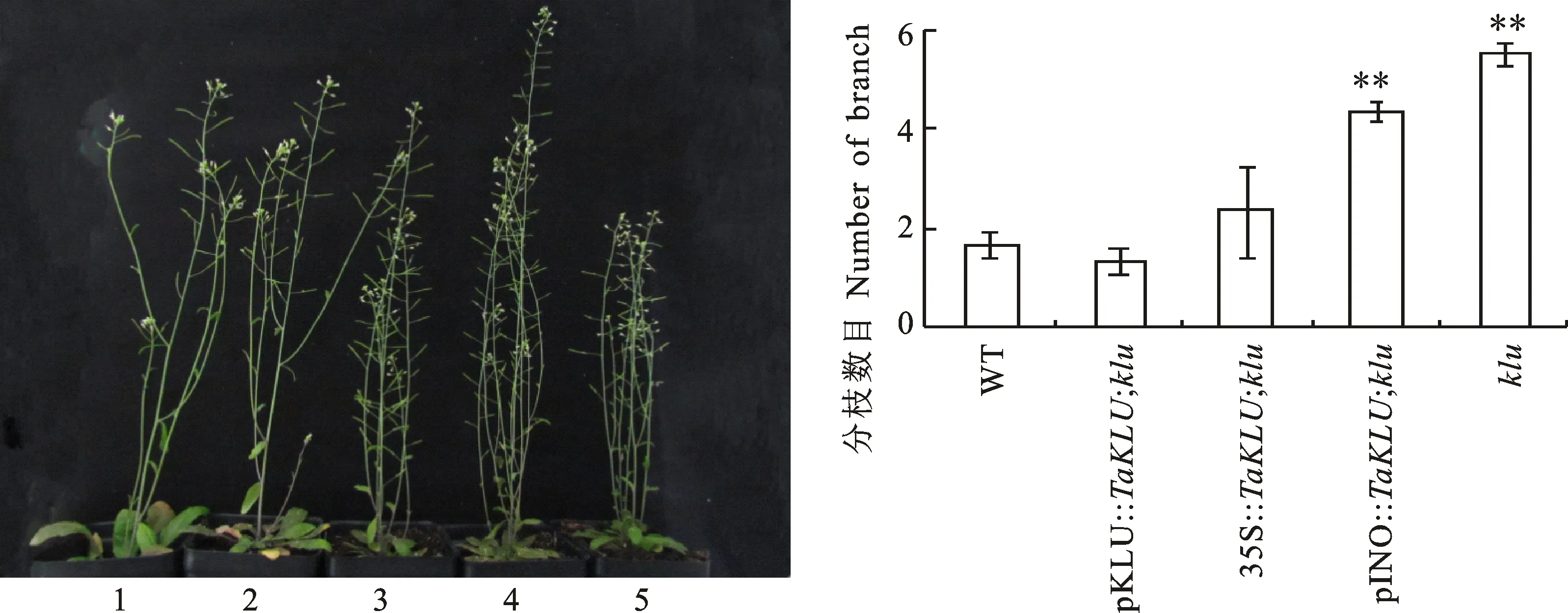
A:不同启动模式下小麦TaKLU回补klu突变体的表型;1:野生型;2:pKLU::TaKLU;klu;3:35S::TaKLU;klu;4:pINO::TaKLU;klu;5:klu突变体。B:不同株系的分枝数目(n=6)。**表示该株系与野生型拟南芥的分枝数目在0.01水平上差异显著。
2.5 KLU基因对拟南芥产量的影响
通过对野生型拟南芥、35S::转基因拟南芥以及突变体的单株产量进行调查,结果(表2)发现,与野生型拟南芥相比,35S::转基因拟南芥的单株产量与野生型无显著差异,而突变体的单株产量极显著下降,降幅为 50.8%。这说明基因不仅影响植物株型,还影响拟南芥的单株产量。
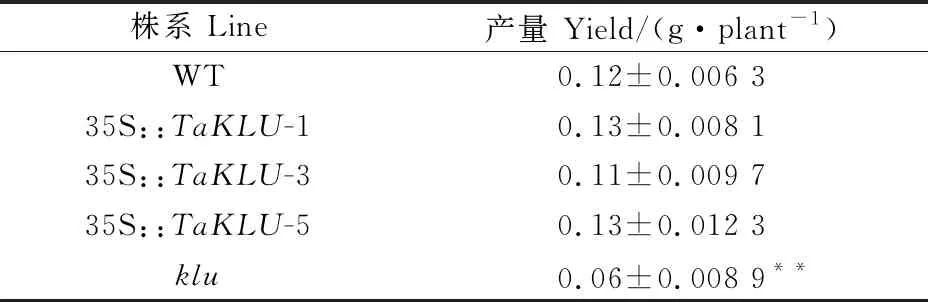
表2 TaKLU基因过表达对拟南芥产量的影响Table 2 Effect of TaKLU gene overexpression on Arabidopsis yield
3 讨 论
在被子植物中,顶端优势和分枝数目在很大程度上决定了植物的株型,并影响植物的营养分配、株高、产量等。作物的驯化也常常涉及株型和顶端优势的变化,通过选择抑制或促进芽的形成来减少或增加分枝数目,进而改变作物的株型和顶端优势。拟南芥基因具有调控叶片和花序生长与发育的功能,其突变体表现为叶片和花序变小,本研究发现,拟南芥基因的功能缺失导致植株株高降低和顶端优势弱化、分枝数目增加、器官变小;相反地,采用组成型强启动子35S驱动小麦基因在拟南芥中过表达,可使拟南芥株高增加和顶端优势增强,分枝数目减少、器官增大(数据未展示),表明基因对植物株型具有调控作用。本研究通过回补试验,观察分析了pKLU::;、35S::;、pINO::;回补株系与野生型拟南芥和突变体的表型差异,发现所有基因回补株系的顶端优势都得到了加强,其中pINO::;回补株系的顶端优势加强的效应最为显著,其主分枝较其他分枝明显伸长,进一步证实了基因在调控植株株型和顶端优势方面功能保守,为基因在作物株型遗传改良方面奠定基础。
本课题组前期研究发现,基因具有正调控种子大小的功能,而拟南芥基因缺失导致种子大小和结实率显著降低,暗示尽管过表达基因会引起种子增大,但过表达基因还会导致顶端优势加剧、分枝数目降低,很可能抵消种子增大对产量增加的正效应,本研究中35S::转基因拟南芥的单株产量较野生型无明显差异,可能就是以上原因造成的。相反,尽管突变体表现为分枝数目增加,但籽粒变小、结实率和顶端优势降低对产量降低具有更强的负效应,本研究中突变体的单株产量较野生型降低了50.8%,推测可能也是这个原因造成的。Adamski等研究发现,对拟南芥中基因进行过表达和敲除,均会导致产量显著降低。因此,适当的顶端优势和合适的分枝数目才有利于作物产量的提高。鉴于此,Guo等对小麦基因的利用提出了新的策略,即在小麦籽粒发育阶段通过局部过表达基因,达到在不影响小麦株型的前提下增加粒重,实现了单株产量显著增加的目的。但该研究并未实现基因在株型改良上的应用。综上,基因作为一种在单子叶和双子叶植物中功能保守的株型调控因子,在作物株型遗传改良育种中仍需要进一步的探索。
目前,研究者普遍认为,顶端优势和分枝数目受激素水平调控。例如,基因编码一种色氨酸转氨酶,水稻突变体由于内源生长素分泌减少,导致顶端优势减弱。基因编码脱落酸生物合成酶,过表达水稻基因可通过参与独脚金内酯对腋芽的调控,从而减少水稻分蘖,调控植株形态。本课题组前期研究也发现,小麦基因在调控器官发育过程中可影响生长素代谢通路,改变生长素的积累,推测基因调控植物顶端优势和分枝数目与激素调控也有关系,但相关机制仍需进一步深入研究。
