小麦铭贤169休眠解除期间种子结构与主要成分的变化
2022-05-20张明婷姚雅鑫王京宏孙风丽奚亚军
张明婷,姚雅鑫,王京宏,孙风丽,张 超,奚亚军
(1.西北农林科技大学农学院,陕西杨凌 712100;2.农业农村部西北地区小麦生物学与遗传育种重点实验室,陕西杨凌 7102100;3.陕西省杂交油菜研究中心, 陕西杨陵 712100)
小麦(L.)是全世界广泛栽种的一种作物,也是历史上第一个被驯化的主食作物。中国是世界上较早种植小麦的国家之一,而黄淮麦区是我国小麦种植面积最大的地区之一,栽培面积占全国小麦总面积的40%,育成和推广的品种数量较多。铭贤169是在黄淮麦区育种试验中广泛使用的条锈病诱发材料,其种子具有休眠特性。休眠的种子可以抵挡不良环境的影响,即使长期贮藏仍能保持较高生活力。植物种子通过自身休眠可以避开外界不利因素,对保持其自身的繁衍具有重要意义。然而在驯化小麦的历史中,为了追求更好的经济效益和快速均匀的发芽,休眠特性被人为弱化,导致现在生产上的骨干品种大多不具有休眠性,加剧了穗发芽现象的发生,穗发芽会导致籽粒品质劣化,食用和种用安全性降低。解决穗发芽问题最经济有效的途径是培育具有强休眠性的品种。
种子的休眠性主要与基因型有关,也受植物激素、光照、温度、营养物质和后熟作用等外界因素的影响。干燥后熟通常是解除新鲜收获的成熟小麦种子休眠的常用方法。小麦种子的后熟期一般为一至两个月左右,后熟初期的种子呼吸强度、耐储藏性、酶活性、生理代谢、品质和发芽率等都会随着后熟时间的延长而发生变化。孙果忠等的研究表明,随着后熟时间的延长,冀麦1号、西农6028等9个小麦品种的种子萌发力提高的程度不同。冯攀屹等研究表明,后熟对小麦中的碳水化合物、含氮化合物、脂类等有很大影响。后熟过程中,小麦发芽率、淀粉、糖类、蛋白质、酶类以及mRNA等均会发生变化。通过转录组分析发现,细胞壁修饰相关程序在种子后熟过程中被激活,激素的转录和淀粉、蔗糖代谢等途径在小麦铭贤169种子休眠至萌发的过渡中显著上调表达。研究表明,后熟诱导小麦种子打破休眠与细胞信号转导和能量代谢相关蛋白质的差异表达有关,如编码α-淀粉酶和淀粉合酶的基因的表达发生了显著变化。但是,有关休眠解除期间小麦种子结构和主要成分变化的报道甚少。
本研究以休眠性明显的铭贤169为试材,探究其种子在发育和后熟阶段的休眠解除过程中淀粉和种皮显微结构以及主要成分的变化,以期为揭示小麦穗发芽抗性机制提供借鉴。
1 材料与方法
1.1 供试材料及种植条件
本试验所用材料铭贤169,2019-2020年度种植于陕西杨凌西北农林科技大学北校区试验田(34°17′24.5″N,108°04′05.2″E),生长条件同当地大田。成熟时取移除穗上端和下端的种子,取穗子中部的种子脱粒、晾晒后在室温下储存备用。
1.2 种子和胚的发芽率测定
在小麦花后10、20、30、40 d分别取穗中部种子500粒,收获后后熟15、30、45、60、75、90 d种子分别取500粒,用于发芽率测定。
种子发芽率测定:取花后或后熟不同时间种子250粒,用5%次氯酸钠进行表面消毒15 min,用无菌水冲洗5次;在直径为9 cm的无菌培养皿中放双层无菌滤纸并加入8 mL的无菌水,将灭菌种子腹沟朝下放置于培养皿,每皿50个种子。在培养箱中进行发芽,发芽温度设置为20/15 ℃(光照/黑暗),光周期为10/14 h(光照/黑暗)。每12 h记录发芽种子数,直到7 d后发芽率不再发生变化(胚根突破种子外皮>1 cm即为发芽)。
胚发芽率测定:参考孙果忠等的方法稍作改动,取开花后或后熟不同时间种子各250粒,用5%次氯酸钠消毒3 min(后熟种子需浸泡4~6 h),再用ddHO冲洗5次;无菌操作下剥离胚,将胚盾片朝下摆放在配制好的MS培养基上,每皿放胚50粒,用parafilm封口,放于培养箱中 20 ℃暗培养。每12 h记录发芽数,直到发芽率不再发生变化,以根尖变白膨大(直径>1 mm)为发芽。
1.3 种子吸水率和活力测定
吸水率的测定:随机选取50粒收获备用种子,称重后置于烧杯中,加入ddHO在室温下浸种。每隔6 h取出种子,用滤纸吸干表面水分后称重,直至连续两次种子质量恒定。
种子活力(TTC)检测参考高俊凤方法。5次重复。
1.4 种皮结构和淀粉粒的显微结构观察
每20 d取收获备用种子5粒,共取6次,用解剖刀切成2 mm厚的横切片,挑选切面平整的样品用双面导电胶粘贴于扫描电镜样品台上,横断面朝上,吹去表面的灰尘后,用真空喷涂仪对样品进行喷金,并在S-3400N型电子显微镜(加速电压5~15 KV)下观察特定部位并拍照。
1.5 籽粒品质测定
采用perten公司的DA7200近红外古物分析仪测定样品水分、吸水率、粗蛋白、面筋和淀粉含量、沉降值,重复5次。
1.6 面粉RVA粘度参数测定
参考Jiang的方法略做改动,准确称取含水量为12%的铭贤169面粉3.0 g置于小铝罐中,加入蒸馏水25 g;用快速粘度分析仪(RVA 4500, Perten, SWISS)测定黏度相关指标。程序设置为在50 ℃保持1 min,以12 ℃·min的速率升温至95 ℃,保持2.5 min,以同样的速率降温至50 ℃,保持2 min;前10 s以960 r·min的速率进行搅拌,之后以160 r·min速率搅拌。重复3次。
1.7 淀粉热稳定性测定
淀粉提取参考peng等的方法并作修改。取5 g铭贤169籽粒在0.5 mol·L的NaCl溶液中浸泡16 h,在研钵中研成匀浆并用500目的尼龙网过滤,直至滤出液用碘染色不变蓝为止。将淀粉匀浆在6 000 r·min下离心5 min,去上清,沉淀依次分别用2 mol·L的NaCl、2% SDS和0.2%的NaOH溶液洗涤,直到沉淀变为白色为止;用丙酮清洗3次,风干后过筛。于-20 ℃贮存备用。每个样品重复2次。
取10 mg淀粉于坩埚中,放入热稳定分析仪中(EXSTAR TG/DTA 6000日本),在氮气氛围下以10 ℃·min的升温速率从30 ℃升至 550 ℃,得到热重-热重微分(TG-DTG)和差热(DTA)谱图。每个样品重复3次。
1.8 淀粉粒径测定
淀粉提取同1.7方法,粒径采用MASTERSIZER-2000激光粒度仪(Malvern UK公司)测定。称取0.5 g铭贤169淀粉悬浮于5 mL蒸馏水中;超声波分散20 s,统计的总粒数达到10 000时停止。数据用激光衍射法进行自动分析。每个样品重复3次。
1.9 淀粉酶活性和可溶性糖含量的测定
淀粉酶活性和可溶性糖的测定参照高俊凤方法。重复3次。
1.10 数据处理
用SPSS进行数据分析,用Excel作图。
2 结果与分析
2.1 铭贤169种子发芽率变化
从图1A可以发现,收获前的胚在花后25 d已经具有发芽能力,开花后35 d胚发芽率增加,胚伸长率增加,胚两端都有明显的芽伸出。而种子在花后35 d没有发芽。开花后40 d,胚发芽率超过40%,种子发芽率只有3%(图1B)。说明在收获前,种子休眠不是由胚引起的,推测胚以外的结构对种子休眠有着正向的作用。收获后的种胚和种子萌发率都随后熟时间延长而升高。在收获后15 d,胚发芽率达到80%,在收获后90 d达到100%(图1C)。在收获后15~30 d,种子发芽率从3%快速增长到40%,在收获30 d天后增长率变慢。以上结果说明,种胚萌发能力始终强于种子的萌发能力;铭贤169的休眠由胚以外的母体成分引起,其可能的原因是胚之外的某些母体成分抑制了胚向外延伸,也可能是胚乳和种皮的机械结构阻碍了胚芽的伸长。

A:开花后25 d、35 d种胚和种子发芽情况(上排:种胚发芽;下排:种子发芽)。B:发育期种胚和种子萌发率。C:后熟期种胚和种子萌发率。
2.2 铭贤169种子吸水率和活力变化
图2结果表明,在后熟期间,干燥的铭贤169种子吸水的初始阶段(0~12 h)吸水速率最快,此后吸水速率逐渐变慢,48 h后吸水率不再发生变化。S5的吸水率最高,48 h的吸水率达到66%,S1的吸水率最低,为61%(图2A)。结果说明,吸水率随着后熟时间的延长而逐渐升高,与发芽率趋势相同。种子活力随后熟时间延长整体呈上升的趋势,从后熟初期的80%增加到100%(图2B),在S3时期种子活力明显下降,可能是后熟期高温潮湿的环境抑制了部分种子的活性,但与后熟发芽率的相关分析表明,种子活力与发芽率关系不大。
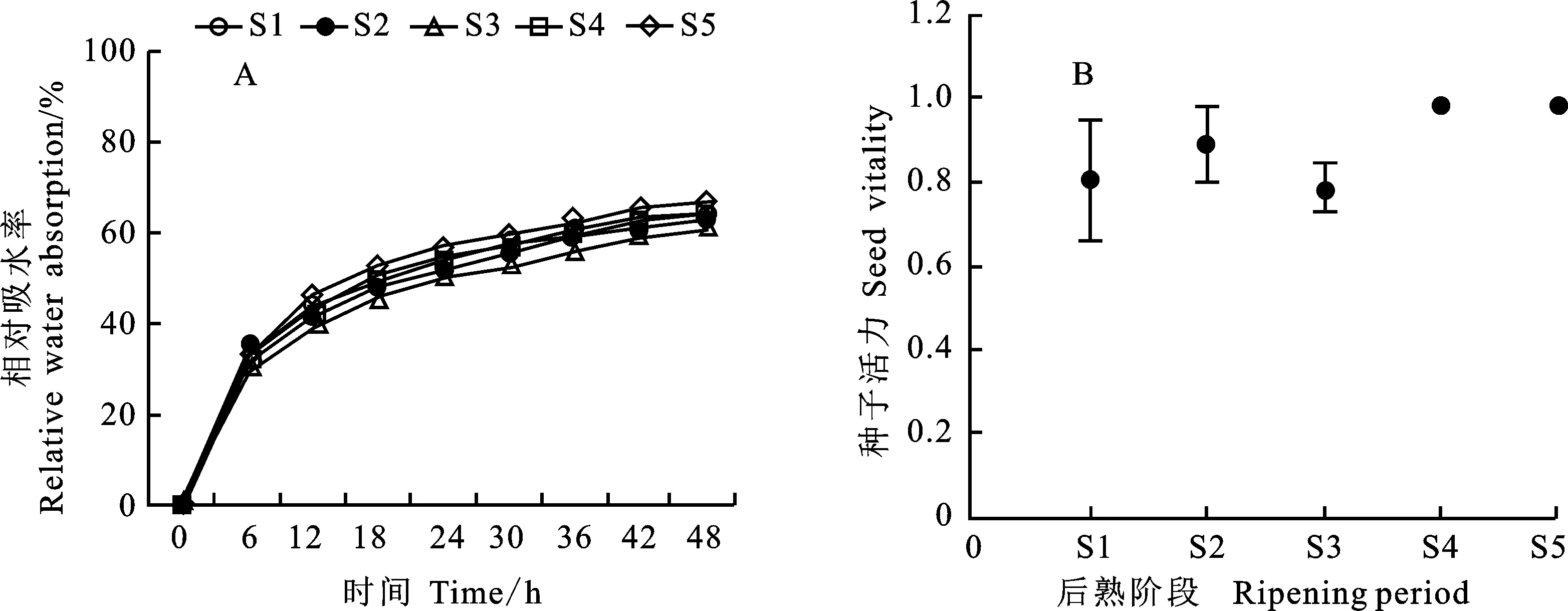
S1~S5分别表示后熟20、40、60、80、100 d。下同。
2.3 铭贤169种皮结构和淀粉粒显微结构变化
铭贤169种子的淀粉粒呈圆盘或透镜状,以饱满的A型淀粉粒为主,周围分布着呈球形的B型淀粉粒。S1时期,A型淀粉粒形状饱满,表面光滑无褶皱无破损,有少量B型淀粉粒分布在周围,且各淀粉粒之间的界限分明(图3)。S2和S3时期,A型淀粉粒表面不光滑,有凸起和小孔出现,偶有破损残缺的淀粉粒,并且形状也不够饱满,略为凹陷和扁平;A型淀粉数量较多,界限比较清晰,周围偶有出现蛋白基质的结构。S4时期,A型淀粉粒表面出现明显的斑点凹陷,类似月球坑的形状,淀粉粒之间的空隙缩小,连接变得紧密;A型淀粉粒数量变少,B型淀粉粒增多,且A型淀粉粒被大量蛋白基质包围。S5时期,淀粉粒破损情况加剧,几乎没有表面完整光滑的A型淀粉粒,且数量减少;B型淀粉粒表面也有破损现象,并且粘连现象严重。S6时期,A型淀粉粒的破损程度加剧,数量少;大部分结构嵌入B型淀粉粒和蛋白基质之中,形成团状结构,A型淀粉粒的界限模糊已经无法与其他物质分离。

图3 休眠解除期间铭贤169淀粉粒的显微结构
铭贤169小麦种皮中,表皮细胞呈条形,细胞中间凹四周高,表面十分不光滑,有褶皱,细胞与细胞间以明显的条状突起隔开,表面分布着灰尘或蛋白基质的碎片(图4)。S1时期,表皮细胞纹路紧凑清晰,排列得很有规律,其中间隔的起伏较强。后期的表皮纹路不如S1时期清晰,排列逐渐分散,不平整,以S5和S6时期这种变化更为明显。S5时期的细胞间隔模糊,褶皱模糊不清晰,宽窄不一致,整体看上去种子表面起伏较大;S6时期褶皱不明显,细胞排列分散无规律。结果表明,随着后熟时间的延长,种子表面细胞从排列紧密到疏松无规律,其可能导致更多的空气和水分进入种子内部从而解除休眠。

图4 休眠解除期间铭贤169种皮表面的显微结构
从图5可以看出,种籽实皮由多层细胞组成,在S1和S2时期,每层组织排列紧密,中间的缝隙较小。S3-S6时期,随着后熟时间的延长,每层细胞之间的距离增加,排列变得疏松,厚度增加。S1时期,胚乳外部的栅栏细胞形态明显,细胞成规则的四边形,表面光滑,且排列紧密,与胚乳结合紧密,胚乳细胞的表面光滑完整。S2时期,栅栏细胞开始变形,出现椭圆形细胞,排列疏松。S3和S4时期,栅栏细胞表面粗糙,细胞周围出现明显的断裂结构,与下层胚乳的结合不如S1时期紧密,胚乳细胞的表面破损,露出内部的淀粉粒。S5时期,栅栏细胞的表面不完整,形状接近圆形;S6时期,栅栏细胞变为扁平的四边形,与种皮和胚乳之间出现明显缝隙,分层明显。

图5 休眠解除期间铭贤169种皮横断面的显微结构
2.4 铭贤169籽粒品质变化
铭贤169种子水分含量和吸水率随着储藏时间的延长而显著增加,对比S1时期和S5时期,二者水分含量和吸水率差异均显著。后熟初期,淀粉含量的变化不明显,在S4和S5时期,淀粉含量显著增加,增幅接近2%。推测S1~S3时期淀粉含量也在增加,由于淀粉含量基数较大,所以表现不明显,直到后熟末期,淀粉累加使得淀粉含量的上升变得明显。沉降值在后熟期间的变化可分为三个阶段,第一阶段沉降值快速降低,从44 mL快速下降到41 mL;在第二阶段(S2~S4时期)沉降值保持稳定,约为41.5 mL;第三阶段是后熟末期,沉降值显著下降(接近2 mL)。推测铭贤169的蛋白质可能在后熟初期和末期发生变化,导致沉降值改变。湿面筋含量和粗蛋白含量在后熟期内整体变化不大。

图6 休眠解除期间籽粒品质变化
2.5 铭贤169种子淀粉糊化特性变化
随着后熟时间的延长,铭贤169的各被测糊化指标均呈上升的趋势,峰值粘度、破损值、终粘度和回生值各后熟时期间差异不显著;S5时期的谷值粘度值显著高于S1时期(图7A)。峰值时间变化显著,S1时期显著低于S5时期(图7B)。糊化温度呈下降趋势,但差异均未达到显著水平(图7C)。整体来看,后熟期间铭贤169糊化特性并未发生明显的变化。
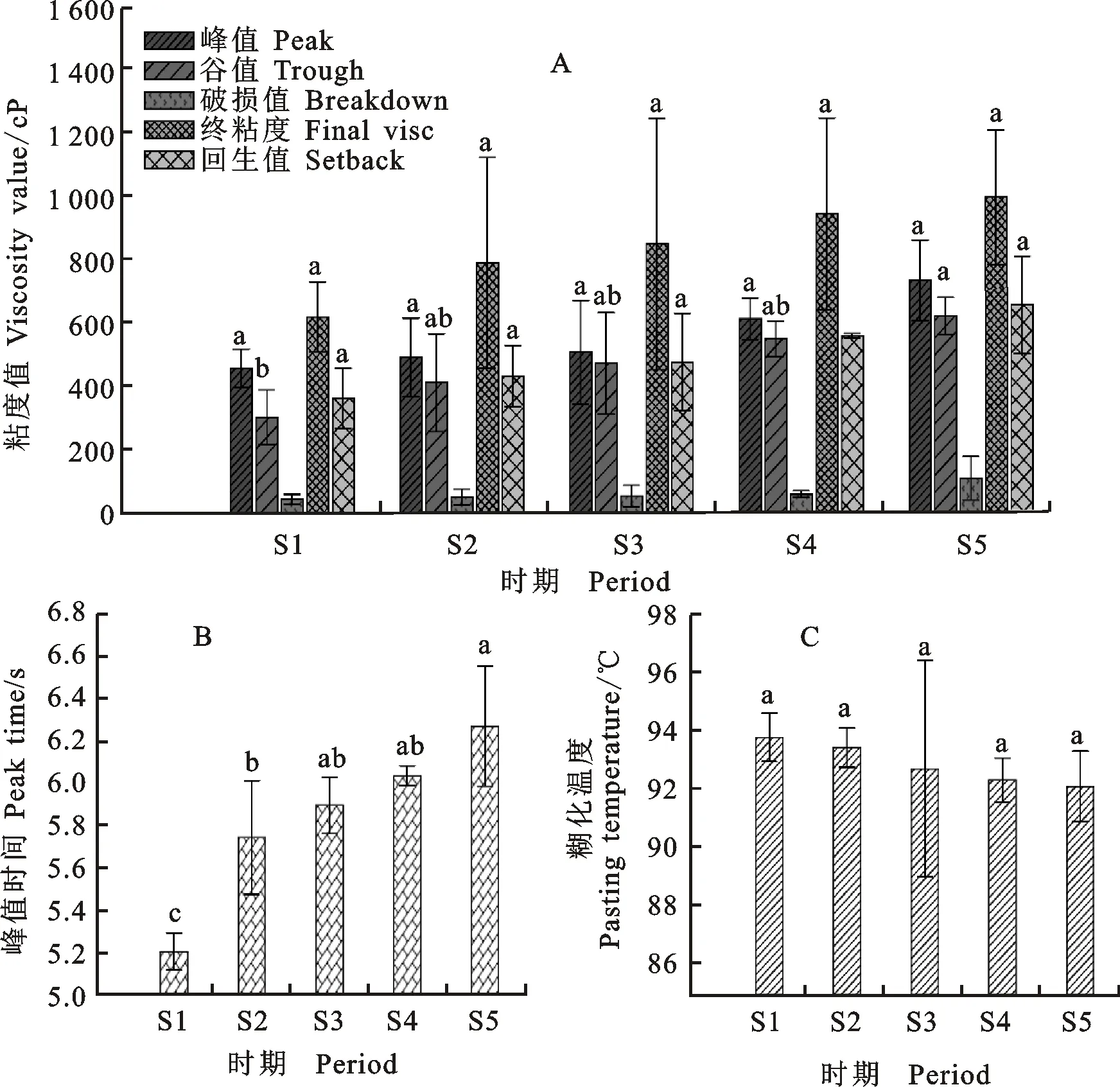
图7 后熟期间糊化特性的变化
2.6 休眠解除期间铭贤169淀粉热特性变化
铭贤169的淀粉有三个明显的吸热阶段(图8A):第一阶段为<140 ℃,对应的DTA曲线为吸热峰,推测主要由水分蒸发引起;各后熟时期种子的吸热反应温度大致相同,S1吸收的能量最多,S3最少;第二阶段(250~330 ℃)对应的DTA曲线为吸热峰,S3~S5吸热峰发生温度大致在300 ℃,而S1和S2的吸热峰出现在温度为 275 ℃,其中S5吸收的热量最多,S2最少,可能是由C-CH、C-O和C-C键发生断裂而引起的;第三阶段为温度>330 ℃,吸热峰的出现温度有较大差异,吸热峰温度最低的是S2,最高的是S5,而反应所吸收的能量是S5最多,S2最少。在S1和S2的差热曲线中,220~250 ℃出现了一个小的放热峰,而其他时期并没有这一阶段的放热(图8A)。推测在小麦后熟期间,初期和末期的淀粉性质可能发生一些变化。差热分析表明,铭贤169在后熟期内淀粉的热稳定性增强,初期阶段的热量变化与末期阶段有所不同。
由TG曲线分析可知,淀粉降解分为两个阶段(图8B)。第一阶段发生在30~120 ℃,由于水分的蒸发而出现小幅度的失重,各后熟时期淀粉的失重速率相近。第二阶段的降解温度为220~350 ℃,淀粉颗粒迅速失重,形成明显失重台阶,各时期的下降拐点不相同。S1发生失重的温度最低,大约为215 ℃,S3温度达到250 ℃左右才开始分解失重。S1和S2的第二阶段曲线的拐点比较接近,而S3~S5的拐点形状相似,说明后熟前后的性质不同。350 ℃后重量缓慢下降,直到550 ℃反应结束时,S1的剩余质量最大,而S5最小(图8B)。从DTG的试验结果可以看出,铭贤169淀粉的失重分为两步,与TG曲线对应。在失重第一阶段,S2的峰值是最低的,峰面积也最小,说明失重速率最慢,失重量最小,热稳定性较差。S5的峰值最高(图8C),失重速率最快,失重最大,热稳定性较强。第二阶段失重的差异明显,S1峰值峰面积最小,而S5最大。表明在淀粉迅速分解的阶段,S1分解温度最低,分解速率最慢;S5分解温度最高,速率最快(图8D)。这些结果说明后熟作用可增加淀粉热稳定性。

图8 休眠解除期间淀粉热差热重变化
2.7 休眠解除时期铭贤169淀粉粒径变化
随着后熟时间的延长,淀粉粒径分布有所差异(表1)。
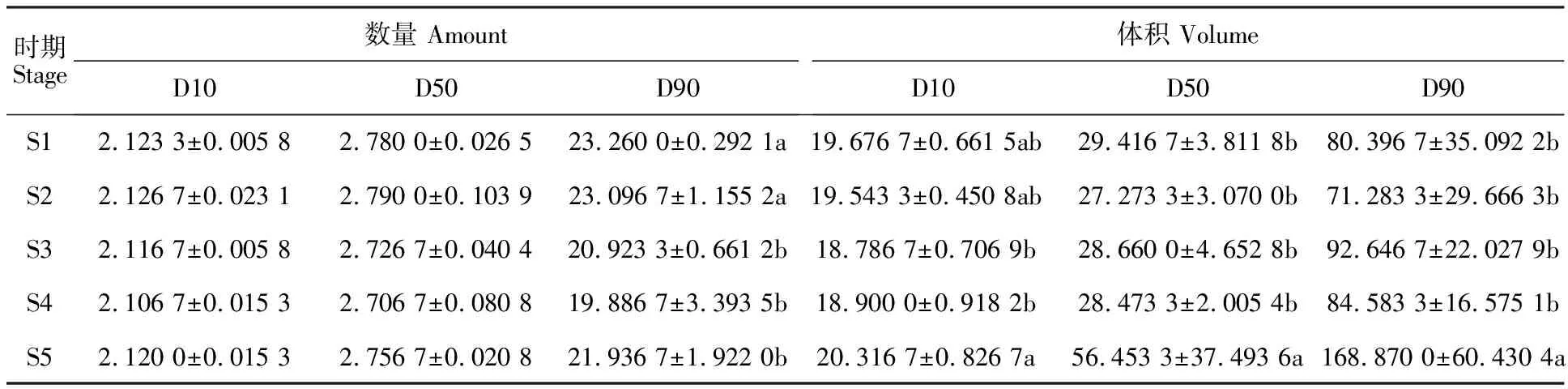
表1 淀粉粒径分布
从数量百分比而言,各后熟时期淀粉的D10值在2.106 7(S4)~2.126 7(S2)μm之间,随着后熟时间延长,粒径逐渐减小(除S5时期),各时期差异不显著。D50中粒径的分布差异不大,散布在2.706 7(S4)~2.790 0(S2)μm之间。D90粒径分布在19.886 7~23.260 0 μm之间,S1和S2与S3~S5之间差异显著,说明淀粉的大粒径随着后熟时间的增加而减小。
体积百分数,D10在S5与S3和S4之间出现显著差异;S5的D50粒径值几乎为其他时期的2倍,差异显著;D90的粒径分布范围为71.283 3~168.870 0,最大值是S5时期。总体来看,粒径体积百分数逐渐减小,但S5时期增大明显,具体的原因还有待探讨。
2.8 休眠解除期间铭贤169种子的可溶性糖含量、淀粉酶活性变化
可溶性糖在铭贤169后熟过程的变化如图9 A。S1时期,铭贤169小麦可溶性糖含量较高,到S2时期明显下降,随后进入一个稳定的时间段(S2~S4),S4~S5期间快速下降。α淀粉酶和β淀粉酶活性随后熟时间延长逐渐下降,但β淀粉酶活性始终高于α淀粉酶,均以S1时期的活性最高。表明后熟作用会降低种子可溶性糖含量,减弱淀粉酶活性。
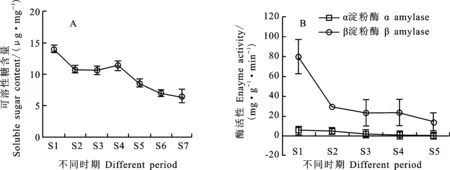
图9 休眠解除期间淀粉可溶性糖含量(A)和淀粉酶活性(B)
3 讨 论
目前,有关小麦休眠机制还没有统一的定论,之前的研究普遍认为,母本的种皮和胚乳对子代的休眠有着重要作用。毛伯仁和吴兆苏将不同休眠期的品种进行正反杂交,结果表明,子一代的休眠性倾向母本。本研究比较铭贤169籽粒建成时期和收获后种胚和种子的休眠解除过程,发现灌浆期种胚在花后25 d即具有发芽能力,花后35 d胚萌发率可达35%,但完整种子仍处于休眠状态。推测铭贤169的休眠性受胚乳和种皮等母体结构的影响。早期研究指出,麦类作物的种皮是制约种子萌发的重要因素,本研究结果也表明,离体胚的萌发率更高,而具有完整胚种皮的籽粒休眠性更强。可能的原因是母体通过进化出一些机制来控制种子的休眠行为,如母本硬化的外包组织(种皮)和大量的胚乳等成分。说明种皮和胚乳对小麦种子休眠和萌发具有一定的 影响。
种皮是小麦籽粒吸水和透气的最大障碍,也是母株组织控制休眠的重要机制。本研究分析了自然条件下后熟的铭贤169种皮显微结构的变化,发现后熟作用会导致表皮细胞排列疏松无规律,组织结构变薄,增加种皮吸水率。这与蒋国梁等的研究一致,籽粒吸水速率与穗发芽呈显著正相关,吸水速率快的品种穗发芽严重,反之则轻。所以种皮结构变化导致吸水速率变化可能是穗发芽的原因之一。本研究表明,刚成熟时期的铭贤169由于种皮结构吸水性不强,因此具有较强的穗发芽抗性,种皮细胞壁的纤维含量和厚度会影响水分的吸收,后熟期间的铭贤169籽粒种皮可能因干燥变得结构疏松,不仅提高了水分的透性,更重要的是有利于氧气的供应和弱化了机械物理阻碍,从而利于发芽。
淀粉是小麦籽粒内含量最多的储藏性碳水化合物,约占总量的70%,主要由直链淀粉和支链淀粉组成。本研究发现,后熟作用使铭贤169种子B型淀粉粒的比例增加,A型淀粉粒破损严重且与蛋白质结合紧密。王 增等的研究结果表明,弱休眠性的小麦淀粉粒分解程度更大,淀粉粒与蛋白基质紧密包裹;而强休眠性的种子内部淀粉粒分解程度较轻,与蛋白基质结合松散。这与本研究结果相同,推测铭贤169小麦种子休眠性与淀粉粒的分解程度和与蛋白质结合方式有关。此外,本研究表明,在后熟期间铭贤169的大淀粉粒径减小(表1),淀粉的失重量减小,热稳定性增强,后熟期间总淀粉含量逐渐增加。这些结果与徐 瑞等,吴俊男等的研究一致,暗示在后熟期间淀粉性质的变化在铭贤169休眠解除机制方面发挥重要作用。
小麦籽粒中蛋白质含量约占12%~14%。在本研究中,由醇溶蛋白和麦谷蛋白组成的面筋含量在后熟期间上下波动,未出现明显变化,说明后熟作用对铭贤169的蛋白质含量的影响不大。有研究表明,新收获小麦在室温条件储存,总蛋白含量基本不变,但蛋白质中各组分会发生变化,部分蛋白质在蛋白酶的水解下变成小分子肽类和氨基酸,同时形成新的氨基酸和蛋白质。推测铭贤169蛋白质含量的波动可能是由于内部组分的变化导致的。
在本研究中,可溶性糖含量在后熟初期剧烈下降,中期平稳后又显著下降,这与徐 瑞的研究结果一致。其原因可能是,小麦在后熟初期还未达到生理成熟,籽粒内部进行着复杂的分解合成作用,导致可溶性糖大量消耗。经过一段时间的稳定后,可溶性糖含量又因为淀粉的合成而逐渐减少。说明铭贤169后熟期间休眠解除与淀粉和可溶性糖含量密切相关。酶活性也是影响小麦后熟期间碳水化合物含量变化的重要因素,其中最主要的是淀粉酶。胚乳中的淀粉需要在淀粉酶的作用下转化为糖,然后运往种胚的生长部位,为构建组织提供底物和能量。本研究结果表明,淀粉酶活性在小麦收获初期较高,在后熟期间逐渐下降。这与孙 辉等和王晓曦等研究结果一致,说明后熟作用降低铭贤169淀粉酶活性是导致休眠解除的重要原因。
