基于Oncomine等数据库分析CDCA基因家族在胃癌中的表达①
2022-05-18陈晓萍吴美珠刘丽雅沈阿灵陈友琴
陈晓萍 吴美珠 刘丽雅 沈阿灵 陈友琴③ 彭 军
(福建中医药大学中西医结合研究院,福州 350122)
胃癌是常见的恶性肿瘤之一,据2018年全球癌症数据分析显示:胃癌在全球发病率居常见肿瘤的第5 位,病死率居第3 位,严重威胁人们的生命和健康[1-3]。虽然目前手术配合放疗、化疗的治疗方法得到有效改善,然而这些治疗策略在进展期胃癌患者中的治疗效果并不理想。分子靶向治疗因其可通过调节肿瘤中异常表达的分子来影响肿瘤细胞的生物学行为,从而达到治疗效果而备受关注。因此,通过寻找新的药物靶点进行治疗以延长进展期胃癌患者生存期是目前该领域研究的热点与难点[4]。
Oncomine 数据库是目前世界上最大的癌基因芯片数据库和整合数据挖掘平台[5]。利用该数据库可挖掘靶基因在大多数主要肿瘤类型及其相应正常组织的表达差异分析数据,同时也可获得靶基因表达与肿瘤重要临床特征的关系,并发现新的潜在生物分子标志和治疗靶点[6]。
细胞分裂周期相关(cell division cycle-associated,CDCA)基因家族由8 个成员组成(CDCA1~8),是细胞增殖的重要调控因子。其不仅是正常细胞所必需的,而且对肿瘤细胞增殖具有重要调控作用。其中,CDCA1 也被称为NUF2,其编码的蛋白质在核分裂和微管稳定方面至关重要[7]。CDCA2 可与蛋白磷酸酶1γ(PP1γ)结合,调节细胞周期中的DNA损伤反 应[8]。 CDCA3 是 SKP1-Cullin-RING-F-box(SCF)泛素连接酶(E3)复合物的一部分,在调节细胞周期中尤为重要[9]。CDCA4也称为HEPP/SEI-3/TRIP-Br3,2001 年由 ABDULLAH 等[10]发现其为参与造血干细胞和祖细胞生长分化内在调节的新基因;同时,其还具有调节肿瘤细胞G1/S 周期转换的功能[11]。CDCA5 在细胞分裂过程中对于姐妹染色单体凝聚和分离方面发挥关键调节作用[12]。CDCA6 又称CBX2,该基因编码的多梳蛋白复合物需要通过染色质重塑和组蛋白修饰在整个发育过程中维持许多基因的转录抑制状态[13]。CDCA7即JPO1,参与c-Myc介导的细胞转化[14]。CDCA8也称为Borealin/DasraB,是脊椎动物染色体乘客复合体的一个组成部分,该复合体是有丝分裂和细胞分裂的重要调节因子,并受细胞周期调节[15-16]。现有的研究已经表明,由细胞分裂周期相关蛋白异常表达引起的细胞周期失调在许多肿瘤的发生发展中扮演重要角色[17-18]。然而,在胃癌的发生发展中鲜有报道。因此,研究细胞周期调控相关基因对于揭示胃癌的发生发展具有重要意义。
本研究利用Oncomine 数据库及其他公共数据库,分析CDCA 基因家族在胃癌中的表达及其与预后的关系,以期初步明确CDCA 基因家族在胃癌发生发展中的作用,为进一步研究其对胃癌的作用机制提供新的理论依据。
1 材料与方法
1.1 基于Oncomine 数据库筛选目标基因并提取数据信息 通过注册Oncomine(https://www.oncomine.org)数据库,获得使用数据库的权限,然后根据研究需求在数据库中设定筛选和提取数据的条件。本研究中筛选条件为:①“Cancer type:Gastric cancer”;②“Gene:CDCA1”或“Gene:CDCA2”或“Gene:CDCA3”或“Gene:CDCA4”或“Gene:CDCA5”或“Gene:CDCA6”或“Gene:CDCA7”或“Gene:CDCA8”;③“Analysis Type:Cancer VS Normal Analysis”;④“Data Type mRNA”;⑤临界值设定条件(Pvalue<0.01,fold change>2,gene rank=top 10%)。P<0.05 为差异有统计学意义。
1.2 基于GEPIA数据库提取数据信息 利用GEPIA(http://gepia.cancer-pku.cn/)数据库分析TCGA(The Cancer Genome Atlas)(https://www. cancer. gov/aboutnci/organization/ccg/research/structural-genomics/tcga)数据库中CDCA 基因家族在肺癌中的mRNA 差异表达。在“Expression DIY”中选择“Boxplot”选项,在设置的数据提取条件为:①“Gene:CDCA1”或“Gene:CDCA2”或“Gene:CDCA3”或“Gene:CDCA4”或“Gene:CDCA5”或“Gene:CDCA6”或“Gene:CDCA7”或“Gene:CDCA8”;②“|Log2FC|Cutoff:1”;③“p-value Cutoff:0.05”;④“Datasets Selection:STAD”;⑤“Matched Normal data:Match TCGA normal and GTEx data”。
2 结果
2.1 CDCA 基因家族在不同类型肿瘤中的表达结果 Oncomine 数据库中共纳入了2 664 项关于CDCA 基因家族在不同类型肿瘤中的表达,其具体结果如图1所示:Oncomine数据库中共收集了301项不同类型的关于CDCA1 的研究结果,CDCA1 表达差异有统计学意义的研究结果共有62 项,其中CDCA1表达增高的研究有56项,表达降低的研究有6 项;Oncomine 数据库中共收集了282 项不同类型的关于CDCA2 的研究结果,CDCA2 表达差异有统计学意义的研究结果共有37 项,其中CDCA2 表达增高的研究有33 项、表达降低的研究有4 项;Oncomine 数据库中共收集了367 项不同类型的关于CDCA3的研究结果,CDCA3表达差异有统计学意义的研究结果共有70 项,其中CDCA3 表达增高的研究有65 项、表达降低的研究有5 项;Oncomine 数据库中共收集了361 项不同类型的关于CDCA4 的研究结果,CDCA4 表达差异有统计学意义的研究结果共有39项,其中CDCA4表达增高的研究有35项、表达降低的研究有4 项;Oncomine 数据库中共收集了305 项不同类型的关于CDCA5 的研究结果,CDCA5表达差异有统计学意义的研究结果共有73项,其中CDCA3表达增高的研究有68项、表达降低的研究有5 项;Oncomine 数据库中共收集了367 项不同类型的关于CDCA6 的研究结果,CDCA6 表达差异有统计学意义的研究结果共有28 项,其中CDCA6 中表达增高的研究有27 项、表达降低的研究有1 项;Oncomine数据库中共收集了295项不同类型的关于CDCA7的研究结果,CDCA7表达差异有统计学意义的研究结果共有61 项,其中CDCA7 表达增高的研究有57 项、表达降低的研究有4 项;Oncomine 数据库中共收集了386 项不同类型的关于CDCA8 的研究结果,CDCA8 表达差异有统计学意义的研究共有57 项,其中CDCA8 表达增高的研究有51 项、表达降低的研究有6项。
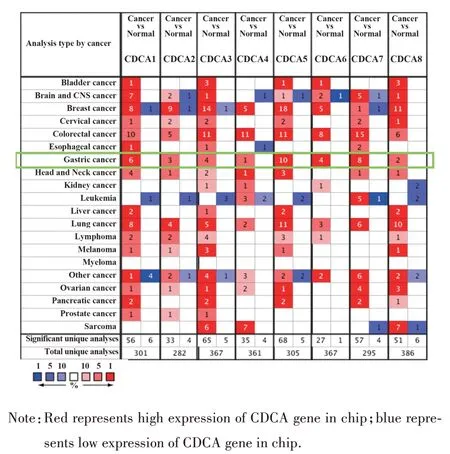
图1 CDCA 基因家族在Oncomine 数据库中不同类型肿瘤研究中的表达Fig.1 Expressions of CDCA gene family in different types of tumors studies in Oncomine database
2.2 CDCA 基因家族在胃癌中的表达 CDCA 基因家族各基因在胃癌中的表达结果如表1所示:CDCA1高表达研究有6项,低表达研究有0项。在这6项不同的胃癌研究中,CDCA1 在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05)。荟萃这6项数据研究发现,与癌旁组织相比,胃癌组织中CDCA1 的表达显著增加(P=3.36E-4),见图2A;CDCA2 高表达研究有3 项,低表达研究有0 项。在这3 项不同的胃癌研究中,CDCA2 在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05)。荟萃这3 项数据研究发现,与癌旁组织相比,胃癌组织中CDCA2表达显著增加(P=3.02E-5),见图2B;CDCA3 高表达研究有4 项,低表达研究有0 项。在这4 项不同的胃癌研究中,CDCA3 在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05);荟萃这4项数据研究发现,与癌旁组织相比,胃癌组织中CDCA3 表达显著增加(P=6.60E-4),见图2C;CDCA4 高表达研究有1 项,低表达研究有0 项。在这1 项胃癌研究中,CDCA4 在胃癌组织中的表达水平高于癌旁组织,差异有统计学意义(P<0.05);CDCA5 高表达研究有10 项,低表达有0 项。在这10 项不同的胃癌研究中,CDCA5 在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05)。荟萃这10项数据研究发现,与癌旁组织相比,胃癌组织中CDCA5 的表达显著增加(P=6.04E-6),见图2D;CDCA6 高表达研究有 4 项,低表达有0项。在这4项不同的胃癌研究中,CDCA6在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05)。荟萃这4 项数据研究发现,与癌旁组织相比,胃癌组织中CDCA6的表达显著增加(P=1.88E-4),见图2E;CDCA7高表达研究有8项,低表达研究有0 项。在这8 项不同的胃癌研究中,CDCA7 在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05)。荟萃这8项数据研究发现,与癌旁组织相比,胃癌组织中CDCA7 的表达显著增加(P=1.86E-4),见图 2F;CDCA8 高表达研究有2项,低表达研究有0项。在这2项不同的胃癌研究中,CDCA8 在胃癌组织中的表达水平均高于癌旁组织,差异有统计学意义(P<0.05)。荟萃这2 项数据研究发现,与癌旁组织相比,胃癌组织中CDCA8的表达显著增加(P=7.73E-9),见图2G。由此可见,CDCA 基因家族中的每个成员在胃癌组织中的表达均显著高于癌旁组织。

图2 CDCA 基因家族在Oncomine 数据库中各项胃癌研究中的表达Fig.2 Expressions of CDCA gene family in gastric cancer from different studies in Oncomine database
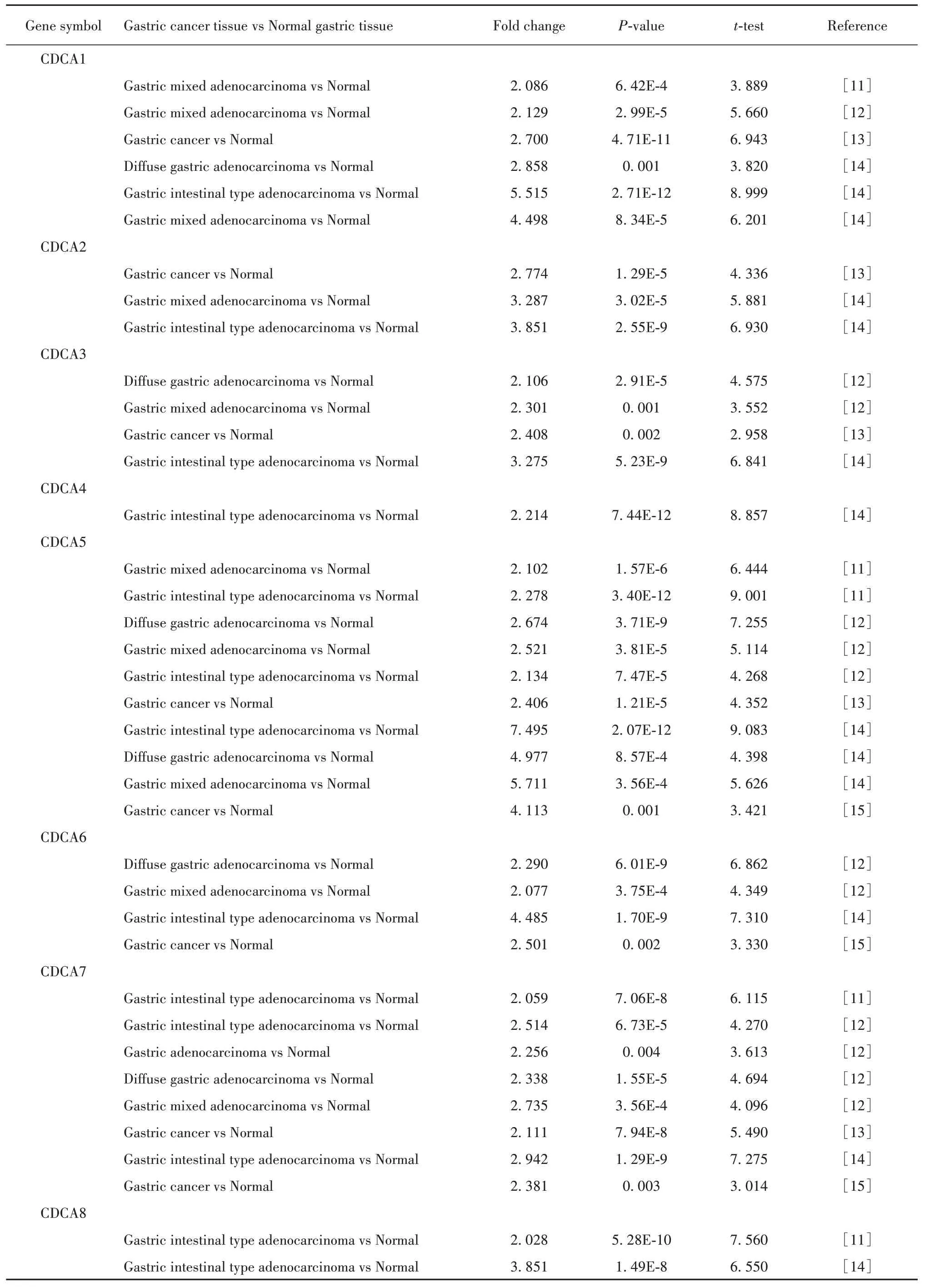
表1 CDCA基因家族在不同研究者数据集的表达情况Tab.1 Expressions of CDCA gene family in datasets of different investigators
2.3 CDCA 基因家族在胃腺癌中的表达 采用GEPIA 平台进一步分析了TCGA 数据库中CDCA 基因家族在胃腺癌中的表达情况,在TCGA 数据库中涉及胃癌旁组织共211 例,胃腺癌组织408 例。CDCA1、CDCA2、CDCA3、CDCA4、CDCA5、CDCA6、CDCA7 和CDCA8 在胃腺癌组织中的表达水平均显著高于癌旁组织(P均<0.001,图3),其差异倍数Log2FC 均>1,见表2。这一分析结果进一步佐证了CDCA基因家族在胃癌组织中的表达显著增加。
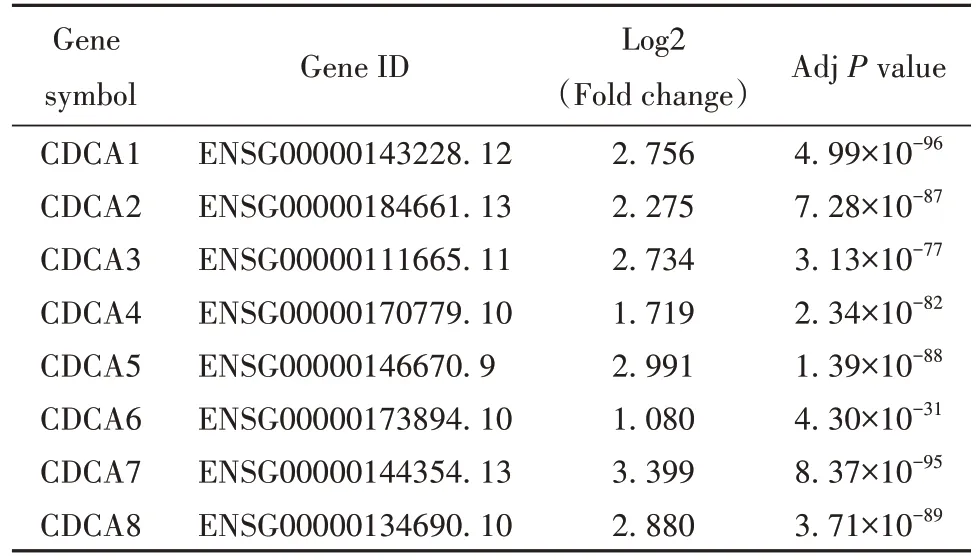
表2 CDCA 基因家族在胃腺癌组织和癌旁组织间的表达差异Tab.2 Significant changes of CDCA gene family expressions between stomach adenocarcinoma and normal gastric tissues

图3 GEPIA 数据库CDCA 基因家族在胃腺癌组织和癌旁组织之间的表达Fig.3 Expressions of CDCA gene family between gastric adenocarcinoma tissue and adjacent tissue in GEPIA database
3 讨论
胃癌是危害人类身体健康的常见恶性肿瘤,我国是世界上胃癌高发的国家之一,因此对胃癌的预防及治疗显得尤为重要[19-20]。目前,胃癌的主要治疗手段仍是手术和化疗。虽然在此基础上,许多新的方法已开始应用于胃癌治疗,然而这些治疗策略在进展期胃癌患者中的治疗效果并不理想。分子靶向治疗的兴起使其成为对进展期胃癌患者进行治疗的极有价值的手段之一。因此通过寻找新的药物靶点进行治疗以延长进展期胃癌患者生存期是目前转化医学研究的热点与难点[4]。
快速的细胞生长和细胞分裂几乎是所有恶性肿瘤的特征,CDCA 基因家族的异常表达可导致肿瘤的发生[21]。其中 CDCA1 也称为 NUF2,该基因编码的蛋白质对核分裂和微管起具有重要作用[16]。CDCA1 敲除会引起动粒-微管附着的异常和纺锤体检查点的激活,最终导致有丝分裂细胞的死亡[7]。有研究表明,通过siRNA 下调CDCA1 基因的表达可抑制肝癌细胞的迁移和侵袭以及结直肠癌细胞增殖[22-23]。本研究表明CDCA1 在胃癌组织的表达显著高于癌旁组织(P<0.001),其差异倍数为2.756倍。
CDCA2 编码的蛋白质参与细胞有丝分裂、核膜重建和损伤修复[24-27]。近年已有研究证实,CDCA2在肺腺癌、胰腺导管腺癌和乳腺癌等肿瘤中高表达,且 CDCA2 的表达与癌症恶性程度相关[8,28-31]。本研究表明CDCA2 在胃癌组织中的表达水平显著高于癌旁组织(P<0.001),其差异倍数为2.275倍。
CDCA3 是细胞进行有丝分裂的必需胞质蛋白,其通过控制有丝分裂入口来调节细胞周期[32]。CDCA3 在包括肺癌、前列腺癌和口腔鳞癌等多种肿瘤中表达增加[33-34],通过调控肿瘤细胞的增殖、耐药、早衰等参与其生物学行为[32]。在本研究中,CDCA3 在胃癌组织中的表达水平显著高于癌旁组织(P<0.001),其差异倍数为2.734 倍。CDCA4 主要定位于细胞核仁中,可能作为转录的抑制因子与细胞周期的有丝分裂相关[35]。现有的研究认为,CDCA4 通过E2F 转录调节途径参与调节宫颈癌细胞和肺腺癌细胞的增殖过程[11];敲低CDCA4的表达能够显著抑制裸鼠体内及体外三阴性乳腺癌细胞的生长增殖能力[36]。在本研究中,CDCA4 在胃癌组织中的表达水平显著高于癌旁组织(P<0.001),其差异倍数为1.719倍。
CDCA5 定位于人染色体11q12.1,其功能是在有丝分裂后期确保姐妹染色单体准确分离[37]。陈墅圳等[38]研究发现,以 siRNA 靶向抑制 CDCA5 基因在肝癌细胞中的表达可抑制肝癌细胞的增殖和促进凋亡。NGUYEN 等[39]的研究也发现,CDCA5 在非小细胞肺癌中高表达与患者预后差相关。在本研究中,CDCA5 在胃癌组织中的表达水平显著高于癌旁组织(P<0.001),其差异倍数为2.991倍。
CDCA6 也称为CBX2,许多基因拷贝数正常的恶性肿瘤表现出周期性的CDCA6 过表达[40]。CLERMONT 等[41]发现 CDCA6 在乳腺癌和前列腺癌中的表达显著上调,且上调程度与肿瘤转移和患者生存期密切相关;而CDCA6 的缺失可促进细胞凋亡,提示CDCA6可能是驱动细胞增殖和转移的关键调节因子。还有研究表明,CDCA6 的表达与乳腺癌的临床特征相关,包括淋巴结转移阳性、肿瘤体积大、HER-2 阳性[42]。在本研究中,CDCA6 在胃癌组织中的表达水平显著高于癌旁组织(P<0.001),其差异倍数为1.080。
CDCA7 作为转录因子Myc 和E2F1 的下游靶基因,参与细胞周期过程,是很多基因的表达转录调控因子[43-44],现有的报道显示,CDCA7 在包括淋巴瘤、卵巢癌和结直肠癌等肿瘤中表达增高[14,45-49]。在本研究中,CDCA7 在胃癌组织的表达水平显著高于癌旁组织(P<0.001),其差异倍数为3.399倍。
CDCA8 是一个细胞周期调节基因,其在定位CPC 到着丝粒、纠正动粒结合错误、稳定双极纺锤体等方面发挥重要作用[37]。现有的研究结果显示,CDCA8 在肿瘤细胞和未分化的人类胚胎干细胞中高表达,而在正常细胞中无表达或有微量表达[50-52]。本研究中,CDCA8 在胃癌组织的表达水平显著高于癌旁组织(P<0.001),其差异倍数为3.880倍。
综上所述,本研究通过分析CDCA 基因家族在不同类型肿瘤中的表达,进一步分析其在胃癌中的表达。分析结果显示,CDCA 基因家族中的各成员在胃癌组织中的表达均显著高于癌旁组织,由此可见,CDCA 基因家族与胃癌的发生发展密切相关,有望成为评估患者病程进展的辅助检测指标和治疗胃癌的潜在新靶点。
