水牛基因组及重要经济性状关联基因解析研究进展
2022-05-16吴思雯RehmanSaifur刘庆友
吴思雯,冯 彤,Rehman Saifur,刘庆友
(中国科学院亚热带农业生物资源保护与利用国家重点实验室,广西大学动物科学技术学院,广西南宁 530005)
水牛()约在6 000 年前被人类驯化,为约20 亿人提供了乳、肉等重要初产品,以及黄油、酸奶、奶酪等优质加工品,带动了至少67 个国家的经济发展,被联合国粮农组织(FAO)誉为养活世界人口最多的大家畜。作为世界重要的经济型家畜,水牛的产奶性状、生长性状、繁殖性状和毛色等性状越来越被重视并纳入育种规划的重要指标。1999—2019 年全球水牛数量增长约25.9%,水牛奶产量增长约106%,水牛肉产量增长约45%,水牛生产快速发展。
虽然水牛整体数量和产业发展较快,但水牛在选择培育中依然面临着生殖效率低、生产潜力差和产奶量低等诸多问题,亦有少数水牛品系濒临灭绝。因此,为了更明确地鉴定现存水牛品种以保护遗传完整性,选择培育优良水牛品种,通过对水牛进行参考基因组分析,研究其基因型和核型,挖掘出一批与其重要经济性状高度关联的基因,对水牛基因组育种具有重要的指导意义。全基因组单核苷酸多态性(Single Nucleotide Polymorphisms,SNPs)选育方式使用全基因组SNP育种芯片作为遗传标记,可同时分析大量SNP 位点的遗传效应,减少世代间隔,加快遗传进展,奶牛全基因组育种技术使其遗传改良育种的速度提高两倍,目前基因组分析在畜禽育种生产中已经得到了广泛应用。本文将对水牛基因组和功能基因解析研究进展进行综述,并对优化建立水牛全基因组选择育种和基因组设计育种进行展望。
1 水牛的经济价值
近年来,水牛基因组及重要经济性状关联基因的研究受到越来越多的关注,这是因为水牛营养丰富,且可以在高温高湿的环境下靠粗饲较好生存。水牛是第二大乳用动物,产奶量约占全球牛奶产量的15%。水牛奶含有100 多种营养成分(表1),与奶牛奶相比,水牛奶含有更高的总干物质,更多的蛋白质(酪蛋白含量极高)和脂肪酸。水牛奶富含钙(191 mg/100 g)和磷(185 mg/100 g),两者比率几乎为1:1,其维生素B含量(0.33 mg/100 g)也比奶牛奶的高出近1 个数量级,同时含有维生素C、维生素A 和维生素H 等。水牛奶还是丰富的必需氨基酸来源,其赖氨酸含量是奶牛奶的两倍。此外水牛奶可以加工成丰富多样口感极佳的奶制品。除了供奶,水牛肉的胆固醇和脂肪含量较低,是优质动物蛋白。Giordano 等研究发现食用水牛肉可以带来明显的心血管益处,适用于有患心血管和脑血管疾病风险的人。水牛肉还含有丰富的锌和复合维生素B,可满足Ordovas 等推荐给女性的营养需求,从而实现个性化营养的目标。本草纲目记载:“水牛肉:安中益气,养脾办,健强筋骨,消水肿,除湿气”。此外,水牛是最重要的稻田劳动力,与水稻产生了独特的稻作文化和水田中天然的循环农业。水牛皮是各种皮革产品的重要来源,水牛粪是天然有机肥料。水牛对维持生态系统也起至关重要的作用,例如Chilika 水牛濒临灭绝,导致其放牧和粪便量减少,一定程度破坏了印度东部奇利卡湖作为野生动物栖息地和渔场的生态系统。水牛除了上述优势特性外,比其他牛更能有效地将劣质粗饲料转化为高营养产品(奶和肉)。

表1 每100g 水牛奶和奶牛奶的营养成分比较
2 水牛基因组研究进展
水牛在分类学上属于哺乳纲(),偶蹄目(),牛科(),水牛属()。野生水牛主要有亚洲水牛亚属()和非洲水牛亚属()2 种类型。亚洲水牛属为目前主要的研究对象,可以分为河流型水牛(Bubalus bubalis)(2n=50)和沼泽型水牛(Bubalus carabanesis)(2n=48)2 个亚种,并在驯化和分化历程中,逐渐表现为河流型水牛种群的系统地理结构较弱,表型多样性较高且品种较多;沼泽型水牛种群缺乏基因流,具有较强的系统地理结构,表型一致性较高。
基因组学主要研究基因组的结构、功能和进化及其对生物体的影响。在进化过程中,基因组会发生许多大大小小的变化,因此研究水牛基因组,首先要探讨水牛基因组结构,同时要解析水牛进化历程以及驯化有关的重要基因座。
2.1 水牛的染色体共线性分析 河流型水牛染色体有25对,而沼泽型水牛有24 对,河流型水牛的4 号染色体和9 号染色体融合为沼泽型水牛的1 号染色体(图1),这2 个亚种除了前5 对是中间着丝粒染色体外,其他所有的染色体(包括性染色体在内的)都是端着丝粒(图1-A)。黄牛有29 对近端着丝粒染色体和1 对性染色体(ΧY),而水牛有19 对近端着丝粒染色体、5 对双臂染色体和1 对性染色体(ΧY)组成(图 1-B)。
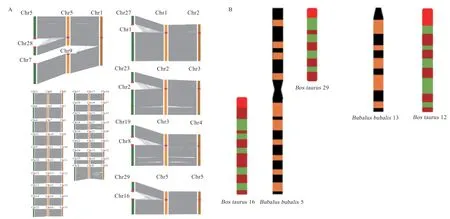
图1 水牛(河流型和沼泽型)和黄牛的细胞遗传学
2.2 水牛的分化与驯化 目前全世界共记录水牛品种有136 个,约2.04 亿头(2019 年),其中约2 亿头(97%)分布在亚洲,其余散布在其他四大洲,这种地理分布是水牛分化与驯化的结果。分子数据表明沼泽型水牛和河流型水牛的分化发生在它们被驯化之前,这种分化可能是由于种群大小、样本、mtDNA 序列、突变率和基因差异造成的。最近的全基因组研究使用Ks 分布和MCMC 树方法评估了沼泽型水牛和河流型水牛之间的分化时间,估计沼泽型水牛和河流型水牛分化在350 万年~110 万年前,并由此推测这两类水牛发生了独立的人工驯化选择。然而Mishra 等认为分化时间在170 万年~1 万年前,而Wang 等通过线粒体基因组分析估算沼泽型水牛分化时间为90 万年~86 万年前。分化时间点仍存争议,这可能需要更多的水牛基因组数据进一步解析。沼泽型水牛在3 000~7 000 年前驯化于印度支那边界地区,河流型水牛约6 300 年前驯化于印度次大陆西部。通常连续的水牛迁移事件发生在不同时间和不同地理尺度上,依据SNPs、mtDNA 和考古证据,可提出2 个独立的迁移事件:河流型水牛早期驯化中心最有可能在印度-巴基斯坦地区,然后向西迁移远至埃及、巴尔干半岛和意大利南部,而沼泽型水牛早期驯化中心在靠近中国/印度支那的边界,从印度支那北部通过东-南-东的路线迁移,在中国、中南半岛、菲律宾和印度尼西亚等推广。更新世冰川事件影响了沼泽型水牛祖先的种群数量,导致种群规模下降,形成偏远的聚集地。在全新世后冰期(1.1 万年~0.6万年前),由于气候改善,沼泽型水牛驯化后得到快速的种群扩张和广泛的地理分布。
2.3 水牛全基因组测序进展 水牛全基因组测序在过去5年里取得了较大进展。截至2020 年12 月31 日,已有36个水牛基因组注释集,29 个在NCBI(https://www.ncbi.nlm.nih.gov),5 个在EBI(https://www.ebi.ac.uk),2 个在中国科学院北京基因组研究所数据中心(https://bigd.big.ac.cn)。水牛基因组总大小、注释到的基因数、测序样本量等都达到新的高度,极大地丰富了水牛基因组数据信息集。例如,Mintoo 等对孟加拉的河流型水牛基因组进行测序组装,拼接基因组大小为2.77 Gb,Contig N50 为25 kb,注释到24 613 个功能基因,确定了159 个基因家族,并发现与其他哺乳动物相比,这些基因家族扩张在水牛中大大加强。与非洲野生水牛基因组(37.21%)相比,家养化水牛基因组中重复DNA占51.19%(1 418 Mbp),高出约13%,且家养水牛49.06% 的基因组为转座元件,其中41.50% 为长散在重复序列。Dutta 等对印度6 个不同水牛品种(Bhadawari、Banni、Murrah、Jaffarabadi、Surti 和Pandharpuri)进行重测序分析,不仅发现了26 247 559个双等位SNVs,其中25 513 085 个位于常染色体,而且其结果与其他驯化物种的特定品种对比突出了河流型水牛的遗传多样性,并提供了跨物种趋同驯化的证据。Luo 等在国际上首次组装获得中国沼泽型水牛和印度河流型摩拉水牛的高质量染色体级别参考基因组(2.631 Gb 和2.646 Gb),Scaffolds N50 分别达到117.3 Mb 和116.1 Mb,分别注释到19 279 个和20 202个基因,其BUSCO 评估率分别为 96.8% 和 96.0%。此外,Luo 等对中国、印度和巴基斯坦等15 个国家共30 个地方品种水牛230 头个体进行了全基因组重测序分析(其中沼泽型水牛共132 头,河流型水牛98 头),共获得了沼泽型水牛1 873 万个、河流型水牛2 372 万个和水牛整体群体3 352 万个SNPs 位点,构建成迄今最完善的水牛基因组遗传多样性数据集。
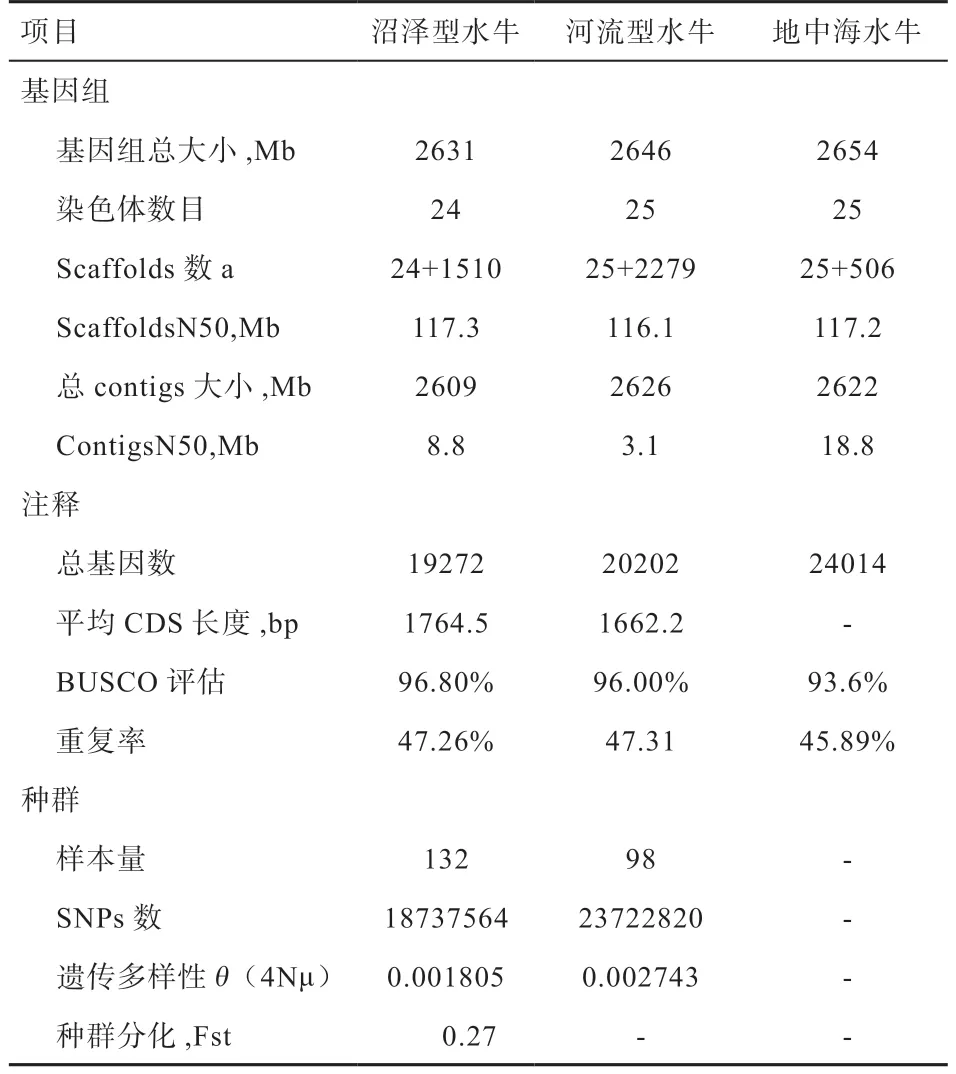
表2 基因组序列统计、注释和种群参数[13]
3 水牛重要经济性状关联基因解析
3.1 水牛耐热性状的相关基因 水牛主要栖息在热带和亚热带地区,能在炎热的夏天在水稻田耕作,抗热应激能力极强,这与热休克蛋白(Heat Shock Proteins,HSPs)基因调控十分相关。HSPs 是包括水牛在内的多种生物中广泛存在的一类蛋白质,最关键的功能是通过抵抗细胞蛋白质的变性来保护细胞免受热应激侵袭,被认为是量化不同牲畜物种中抗热应激强度的潜在生物标记。目前水牛基因组中共发现64 个基因,其中39 个属于基因家族,4 个属于基因家族,8 个属于HSPB 家族,10 个属于基因家族,家族、家族和家族各1 个。研究发现水牛HSP 家族基因的SNPs、外转录子和启动子区域与水牛的产奶量、耐热性、应激恢复力和疾病易感性呈显著关联。Luo 等研究发现水牛基因组中HSPs 基因家族显著扩张,HSPs 的拷贝数显著多于其他物种,是胁迫诱导的分子伴侣,与水牛的环境胁迫适应相关。其中家族的研究较为成熟。与家族的其他基因相比,基因的非同义SNPs 与同义SNPs 的比例较高(0.59),这表明基因由于与细胞耐热性密切相关而处于选择状态,这在Murrah 和Tarai 水牛中有所体现,即在夏季的表达更高。另外基因与繁殖性能有关,其SNPs 多态性影响公水牛精液质量,且mRNA 在水牛卵母细胞热环境和冷环境处理中具有显著差异,该基因表达上调后具有促进细胞凋亡的效果,在牛科动物胚胎中常用其表达量作为热应激程度的标识指标。
3.2 水牛繁殖性状的相关基因 水牛繁殖效率低,筛选繁殖性状关联基因以提高水牛的生产效益备受关注。Lei 等报道,可能通过调节颗粒细胞的增殖和凋亡来促进水牛卵泡的选择和优势化,添加重组蛋白会引起颗粒细胞关键细胞周期调节因子(如cyclin D1 和cyclin D2)的表达上调,而且还上调了在生长滤泡的选择和控制中起重要作用的系统和等的表达。黄体生成素多肽基因和促性腺激素释放激素受体基因的SNPs 与中国水牛精子质量性状之间存在显著关联。某些特定基因型与精子性状有关。Cheng 等通过连锁不平衡分析鉴定出4 个单倍型(H1:ACG,H2:CCG,H3:CTA 和H4:CCA),其中基因型为H2H2 的水牛射精量较多,单体型为H1H4 的水牛精子密度较高。Wang 等通过同种分析方法鉴定出3 个单倍型(H1:TAI,H2:CT-和H3:TT-),合并基因型H1H2 和H1H3 的水牛射精量较多,且中18 953 bp 处的GGCAAAGTAA 缺失与较多射精量显著相关。基因编辑的细胞色素P450 芳香酶调节雌激素的生物合成,并且基因表达降低可能会提高卵巢氧化性损害的发生率,影响雌水牛的繁殖效率。
3.3 水牛产奶性状的相关基因 产奶性状的遗传标记选择是提高水牛产奶量的重要途径,在不同的水牛品种中,目前已报道了19 个与产奶性状有关的候选基因,包含47 个突变,分别发生在启动子区(4 个)、内含子区(24 个)和编码区(19 个),而这19 个编码区中有13 个突变是非同义的,会导致氨基酸发生替代。如表3 所示将这19 个与产奶性状相关的基因分为4 大类。Iamartino 等研究发现了与牛奶生产相关的9 种重要SNPs,其中重要的SNPs 位于基因的下游~51 kb 处,其内含子区域的SNP 与牛的乳脂产量相关。生长激素受体(Growth Hormone Receptor,)基因多态性与埃及水牛产奶量、牛奶质量以及分子变化有关,在GHR 突变型AA 单倍型水牛的体细胞中GH 和IGF1 表达显著增加,表明c.380G>A(该SNP 位于GHR 胞外结构域中)和c.836T>A(该SNP位于GHR 跨膜结构域中)SNPs 可能直接或间接(通过与其他致病性SNP 连接)改变GH 表达,进而诱导IGF1 表达,造成这些基因型水牛的产奶量更高。Fan 等在河流型和沼泽型水牛的基因编码序列中发现13 个SNPs,其中8 个SNPs 是非同义的,其中c.580T>C 和c.642C>G 引起的氨基酸变化可能影响水牛S2-酪蛋白(S2-casein,S2-CN)合成,最终影响产奶量。
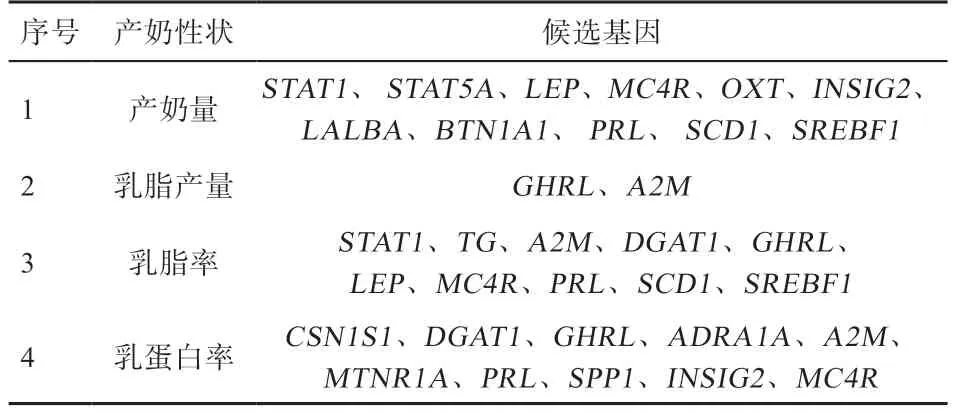
表3 具有不同产奶性状的候选基因[35]
3.4 水牛役用性状的相关基因 中国水牛几乎都是沼泽型水牛,是稻作文明的主要劳动力,温顺耐劳。研究在沼泽型水牛基因组中发现了与运动相关的一些受选择的基因(和),这些基因参与信号通路和黏着连接通路,与水牛的稻田耕作耐力强和肌肉低氧代谢息息相关。沼泽型水牛的催产素受体基因呈现多态性明显降低的信号,基因多态性与人类多种社会行为(包括亲情、社会认知、共情和利他行为等)相关,由此推测是水牛温顺听话性格形成的关键调控基因。此外,信号通路中的关键基因在沼泽型水牛基因组中表现出强烈的选择信号(π 值低,Fst 值高,Tajima's D 值为负值),改变基因的表达可实现骨骼肌纤维由快肌纤维向慢肌纤维的定向分化,充分表明水牛基因受到选择与稻作役用性能息息相关。
3.5 水牛的抗病相关基因 水牛与其他同类动物相比,具有较强的抵抗感染的先天免疫力,对肺结核、乳腺炎、蜱虫病或巴贝斯虫病的抵抗力更好。Sun 等报道,在水牛自身免疫性疾病中起重要作用,在淋巴细胞与脾基质成分的正确结合中起着重要作用,与子宫炎有关,和与结核病有关,这些基因表达增强了水牛环境适应能力和抗病性。此外,(干扰素-)基因中的g.4467G>A 多态性影响miR-125b 的靶序列并赋予其对水牛中牛结核病的敏感性。在埃及水牛群体中追踪到补体C3 基因第27 外显子C>A 的替换与乳腺炎的潜在征兆显著相关。
3.6 水牛毛色调控基因 毛色是水牛重要的体貌特征,为显性遗传性状,尽管目前已有一些报道其分子调控机制,但尚未被系统解析。沼泽型水牛通常是深灰色,但纯白色的是一个常见变种,发生频率高达10%。白色斑点水牛在印度尼西亚被高度重视,其基因中含有两种导致功能丧失的独立突变,包括供体剪接位点的突变和过早终止密码子突变,这可能是造成白斑皮肤的重要原因。张毅等研究发现基因的高表达阻碍了黑色素细胞发育,导致白水牛皮肤基底层缺乏成熟黑色素细胞及色素颗粒,首次系统地揭示了水牛白毛色性状形成的分子机制。因在白水牛Agouti 信号蛋白()基因上游存在一个LINE-1 转座子的插入,长2 809 个核苷酸,只含有完整LINE-1 转座子的5'-UTR区域,但仍行使基因新的强启动子功能,使白水牛皮肤基因的表达量提高了近10 倍,且LINE-1转录序列与基因编码外显子形成白水牛特有的融合转录本。
4 小 结
近年来随着众多与畜禽重要经济性状相关位点的挖掘及其功能基因的定位,由此产生的标记辅助选择育种技术逐步完善,因此有效推动了包括水牛在内的多种动物遗传改良进展。水牛基因组研究与重要经济性状功能基因的挖掘得到加速,基因组选择育种技术受到全球关注。目前已有一些关于水牛基因组的研究被报道,但水牛基因组数据仍然缺乏,特别是我国经过近50 余年杂交改良,存养了一批优秀的杂交水牛群体,亟需建立现代基因组选择育种技术加快遗传进展和水牛新品种选育。水牛基因组和功能基因解析相对滞后成为制约我国水牛产业发展的瓶颈问题,亟待加速解决。水牛基因组、基因组变异和重要经济性状关联基因的进一步解析将加速建立水牛基因组水牛血统溯源、基因组选配和优化的基因组育种芯片技术,以及针对重要功能基因的基因编辑技术,最终为人类培育出产奶量高、个体大、性格温顺、抗热应激且耐粗饲的理想水牛品种。
