基于转录组测序技术分析愈创木酚对禾谷镰刀菌的抑菌机制
2022-05-16张瑶高弢马桂珍史建荣
张瑶 高弢 马桂珍 史建荣


摘要:植物源酚类化合物愈创木酚可以显著抑制禾谷镰刀菌的生长,本研究利用转录组测序分析愈创木酚对禾谷镰刀菌的作用机制,采用RNA-Seq技术从转录组水平分析禾谷镰刀菌在0.4 μl/ml愈创木酚胁迫下的响应机制。结果显示,共筛选到905个差异表达基因表达量发生了变化,其中表达量上调的基因为464個,表达量下调的基因为441个。差异表达基因的COG、GO及Pathway功能分析发现,愈创木酚影响禾谷镰刀菌氧化应激反应、膜组分及离子运输途径。表明,愈创木酚处理后,禾谷镰刀菌的细胞膜完整性受到了破坏,Ca2+运输途径受到破坏,同时愈创木酚作为抗氧化剂也能显著影响禾谷镰刀菌的氧化应激反应。
关键词:禾谷镰刀菌;愈创木酚;转录组;抑菌机制
中图分类号:Q946.887文献标识码:A文章编号:1000-4440(2022)02-0343-09
Analysis on the antifungal mechanism of Fusarium graminearum treated by guaiacol based on transcriptome sequencing
ZHANG Yao GAO Tao MA Gui-zhen SHI Jian-rong
[1.Jiangsu Ocean University, Lianyungang 222000, China;2.Institute of Food Safety and Nutrition, Jiangsu Academy of Agricultural Sciences/Key Laboratory of Control Technology and Standard for Agro-Products Safety and Quality, Ministry of Agriculture and Rural Affairs/Key Lab of Agro-product Safety Risk Evaluation(Nanjing), Ministry of Agriculture and Rural Affairs/Key Laboratory of Food Quality and Safety, Nanjing 210014, China]
Abstract:The plant derived phenolic compound guaiacol can significantly inhibit the growth of Fusarium graminearum. In this study, the action mechanism of guaiacol on F. graminearum was analyzed by transcriptome sequencing, and the response mechanism of F. graminearum under 0.4 μl/ml guaiacol stress was analyzed by RNA-Seq technique at transcriptome level. The results showed that, 905 differentially expressed genes (DEGs) were screened to be varied in expression quantity, of which 464 were up-regulated and 441 were down-regulated. Analysis on the function of COG, GO and pathway of DEGs showed that, guaiacol affected oxidative stress response, membrane components and ion transport pathways of F. graminearum. In conclusion, the integrity of the cell membrane and Ca2+ transport pathway of F. graminearum were damaged after treated with guaiacol, and guaiacol could also significantly affect the oxidative stress response of F. graminearum as an antioxidant.
Key words:Fusarium graminearum;guaiacol;transcriptome;antifungal mechanism
江苏农业学报2022年第38卷第2期
张瑶等:基于转录组测序技术分析愈创木酚对禾谷镰刀菌的抑菌机制
主要由禾谷镰刀菌复合种(Fusarium graminearum species complex)引起的小麦赤霉病是世界范围内广泛发生的一种主要病害[1]。小麦赤霉病不仅会造成小麦产量降低,还会产生多种镰刀菌毒素,严重影响小麦质量安全,威胁人畜健康。筛选、培育和种植具有稳定抗赤霉病和抗毒素积累的小麦品种是防控镰刀菌危害最安全有效的措施之一,但目前尚未开发出可以商业化种植的抗病品种。利用化学药剂进行防控是控制镰刀菌毒素产生和累积的最有效的措施之一[2]。自20世纪70年代以来,中国使用的多菌灵等苯并咪唑类和戊唑醇等三唑类杀菌剂,在控制病害流行、稳定小麦生产上发挥了重要作用[3]。但是,长期单一使用化学农药导致中国长江中下游部分小麦产区病原菌产生明显的抗药性[4],严重威胁了环境生态健康。因此,探寻针对禾谷镰刀菌的新型高效杀菌剂,已成为目前有效防治小麦赤霉病的关键突破口之一。
愈创木酚(2-甲氧基酚),别名甲基儿茶酚,是一种白色或微黄色结晶或无色至淡黄色透明油状液体,在自然界中主要存在于愈创树脂或松油中,也是木材干馏所得杂酚油中的主要成分[5]。愈创木酚在工业上用途广泛,常用愈创木酚来生产各种香料,如丁香酚、香兰素和人造麝香[6]。愈创木酚在医药上也有大量应用,它可被用于合成苯磺酸愈创木酚(愈创木酚磺酸钾)、用于制作局部麻醉剂或防腐剂[7],还可用于制备祛痰口服液和治疗消化不良[8]。目前,利用愈创木酚抑制植物病原菌方面的研究较少。有研究发现,一些含有取代酚(丁香酚、百里香酚、卡瓦克酚和愈创木酚)的精油表现出很强的抗菌和抗氧化作用[9],而多种具有抑菌作用的植物提取物及木醋液中均含有大量的愈创木酚[10-12]。前期研究发现愈创木酚能够抑制禾谷镰刀菌的生长和产毒,但是抑菌机制尚不明确。本研究借助转录组测序手段初步分析愈创木酚对禾谷镰刀菌的杀菌作用及其抑菌机制,为禾谷镰刀菌的防治提供新思路,为开发新的、具有优良防效的植物源农药提供应用基础。
转录组,是指特定生长阶段某组织或细胞内所有转录产物的集合,主要包括mRNA和非编码RNA(ncRNA)[13]。利用新一代转录组测序RNA-Seq (RNA-sequencing)分析愈创木酚处理后的禾谷镰刀菌转录组的变化,筛选鉴定差异基因,并通过GO功能和KEGG途径注释分析,探究禾谷镰刀菌响应愈创木酚胁迫的关键基因,可为后续阐明愈创木酚作用机制的基因组学研究提供参考。
1材料与方法
1.1试验材料
供试菌株为禾谷镰刀菌标准菌株PH-1;供试药剂为99%愈创木酚(Macklin),4 ℃保存备用。PDA培养基:马铃薯200 g/L,葡萄糖20 g/L,瓊脂20 g/L。YEPD培养基:葡萄糖 20 g/L,酵母粉 3 g/L,胰蛋白胨10 g/L,调pH至7.0,高温高压灭菌后常温保存。PrimeScript TM RT反转试剂盒 (宝日医生物技术有限公司产品,货号:DRR036A)用于RNA反转成cDNA;TB Green TM Premix Ex TaqTM Ⅱ(宝日医生物技术有限公司产品,货号:DRR820A)用于实时荧光定量PCR。
1.2试验方法
1.2.1禾谷镰刀菌菌丝处理与RNA提取收集禾谷镰刀菌分生孢子(1 ml 105个)接种至YEPD培养基中,25 ℃下180 r/min振荡培养24 h,加入愈创木酚提取液使其终浓度为0.4 μl/ml,以不加愈创木酚为对照组,继续振荡培养24 h后收集菌丝并冻干研磨成粉末。使用TRIzol 试剂从组织中提取总RNA,并使用DNA酶(宝日医生物技术有限公司产品,货号:2270A)去除基因组DNA。
1.2.2构建文库及转录组测序提取样品的总RNA送至上海美吉生物医药科技有限公司进行转录组测序。首先使用fastx_toolkit_0.0.14软件对原始数据的相关质量进行评估,包括碱基质量分布统计、碱基错误率分布统计和碱基(A、T、G、C)含量分布统计;随后,使用SeqPrep和Sickle软件对测序接头序列、低质量读段、不确定碱基信息率较高序列及长度过短序列进行去除,获得质控后的原始数据,即clean data(reads),利用TopHat2和HISAT2将clean reads与参考基因组Fusarium_graminearum (GCF_000240135.3)进行比对,获得用于后续转录本组装、表达量计算等的 mapped data(reads),同时对该转录组测序的比对结果进行质量评估。使用StringTie或Cufflinks软件对Mapped Reads进行拼接和组装。
1.2.3差异表达基因分析及功能分析将转录组组装获得的所有基因和转录本在六大数据库(NR、Swiss-Prot、Pfam、EggNOG、GO和KEGG)中进行比对分析,获得基因和转录本的功能信息。通过与NR数据库比对,可以查看转录本序列与相近物种的相似情况,以及同源序列的功能信息;通过与Swiss-Prot数据库比对,可以了解转录本注释中蛋白质的功能、转录后修饰、特殊位点和区域、二级结构、四级结构。通过Pfam数据库比对可以对组装出来的转录本进行蛋白质家族的注释;EggNOG提供了更细致的OG分析,可以根据物种所属的进化分支选择参考数据集;利用GO数据库可以获得转录本的功能分类情况;通过与KEGG数据库比对,获得基因或转录本可能参与的具体生物学通路情况。使用RSEM、Kallisto和Salmon软件对基因的表达量进行定量分析,随后根据基因/转录本在不同样本间的表达情况,对样本间共有与特有表达基因/转录本(venn分析)、相关性和主成分(PC)进行分析。
在获得基因/转录本的Read Counts数后,采用DESeq2、DEGseq或edgeR 软件,对多样本项目进行样本间或组间基因/转录本差异表达分析,鉴定出差异表达的基因/转录本。在EggNOG、GO和KEGG 3个数据库中对差异表达基因进行功能分类分析,随后利用Goatools软件进行GO富集分析,从而分析获得的显著差异表达基因主要具有哪些GO功能,分析方法为Fisher精确检验,当经过校正的P值<0.05时,认为此GO功能存在显著富集情况;采用R脚本对差异富集基因/转录本进行KEGG Pathway富集分析,计算原理同GO功能富集分析,当经过校正的P值<0.05时,认为此KEGG Pathway功能存在显著富集情况。
1.2.4荧光定量PCR验证分别提取愈创木酚处理和对照组的禾谷镰刀菌RNA,并反转为 cDNA,反应程序:37 ℃ 15 min,85 ℃ 5 s,4 ℃保存。总体系10.0 μl:2.0 μl 5×PrimeScriptTM Buffer,0.5 μl PrimeScriptTM RT Enzyme Mix I,0.5 μl Oligo dT Primer (50 μmol/L),0.5 μl Random 6 mers (100 μmol/L),2.0 μl total RNA,4.5 μl RNase Free dH2O。以cDNA为模板进行实时定量PCR反应,反应程序:96 ℃预变性120 s;94 ℃变性5 s,56 ℃退火20 s,72 ℃延伸20 s,45个循环。融合阶段:94 ℃变性20 s; 56 ℃退火20 s,72 ℃延伸20 s。总体系25.0 μl:12.5 μl TB Green Premix Ex Taq,1.0 μl PCR Forward Primer,1.0 μl PCR Reverse Primer,2.0 μl cDNA;8.5 μl dH2O。反应所用引物见表1。获得的所有数据参照GAPDH基因(编号:FGSG_06257.1)进行校准,目的基因的相对表达水平利用LightCycler 96 SW 1.1分析软件生成。每个处理设置3个重复。
2结果与分析
2.1转录组测序数据质量控制
试验设置对照(CK)和愈创木酚处理(Gua),各有3个生物学重复,建立了6个cDNA文库。经过测序后,禾谷镰刀菌在对照、愈创木酚处理下的平均Clean reads分别为42 887 328和45 133 555,Q20(测序质量在99.0%以上的碱基占总碱基的比例)分别平均达到98.86%和98.59%,Q30(测序质量在99.9%以上的碱基占总碱基的比例)分别平均达到94.18%和93.94%,G+C平均含量分别为52.63%、52.76%。以上结果说明,本次测序所得cDNA文库质量高,可以进行后续生物信息学的进一步研究。具体数据见表2。
2.2转录组表达量差异分析
共检测到表达基因12 611个,其中已知基因12 000个,新基因611个。分析对照组的样品和处理组的基因表达量,默认参数为:P<0.05 & |log2FC|≥1(FC表示2个样品间基因表达量的比值)。愈创木酚处理禾谷镰刀菌后,共有905个基因表达量发生了变化(图1),其中表达量上调的基因为464个,表达量下调的基因为441个(图1A)。表达差异倍数大于5的基因有65个,占比7.18%,其中表达量上调的基因23个,表达量下调的基因42个;表达差异倍数小于5的基因占比92.82%,其中表达差异倍数在1~2倍的基因有378个,表达差异倍数在2~5倍的基因有462个,表达量上调的基因210个,表达量下调的基因252个。
2.3转录组的COG注释
对愈创木酚处理后的差异表达基因进行COG功能分析,差异表达基因共参与了25个代谢过程。如图2所示,除去652个功能不明的差异表达基因,其余差异表达基因中39个与翻译后修饰、蛋白质翻转、伴侣相关,35个与碳水化合物的代谢运输相关,30个与RNA的转录相关。
2.4差异表达基因的GO功能富集分析
对愈创木酚处理后的差异表达基因进行GO功能注释分析,将差异表达基因分为参与的生物学过程、构成细胞的组分、实现的分子功能三大类。结果如图3所示,共有706个差异表达基因属于生物学功能类,其中有285个差异表达基因为代谢过程相关基因;共有681个差异表达基因属于构成细胞的组分,其中有252个差异表达基因属于膜组分相关基因;共有609个差异表达基因属于实现的分子功能类,其中有294个差异表达基因为催化活性相关基因。
对注释完的差异表达基因进行GO功能富集分析,在差异显著(P<0.05) 的前提下总结基因差异富集度排名前20位的基因功能,发现差异表达基因主要富集在氧化还原相关生物进程、膜整体组成成分以及离子运输途径等(图4)。表明愈创木酚能够影响禾谷镰刀菌代谢过程,使细胞膜组分发生变化。
2.5差异表达基因的KEGG Pathway富集分析
将愈创木酚处理后的差异表达基因按照參与的通路或行使的功能进行分类,结果如图5所示。上调表达基因的功能主要为新陈代谢途径中的氨基酸代谢途径、碳水化合物代谢途径及能量代谢途径,遗传信息处理途径中的折叠、分类和降解途径,细胞过程途径中的运输和分解代谢途径;下调表达基因的功能主要为新陈代谢途径中的碳水化合物代谢途径和脂质代谢途径。
采用R脚本对获得的差异表达基因/转录本进行KEGG Pathway富集分析,按P<0.05的前提下显示显著的富集结果,P值越小表示越显著。主要富集途径如表3所示,差异表达基因主要富集在新陈代谢途径的氮代谢通路和谷胱甘肽代谢通路中。
2.6差异表达基因的qRT-PCR验证
对转录组测序结果中表达量差异倍数为5倍以上的差异表达基因进行筛选分析,除去功能未知的蛋白质后,选取另外14个基因进行qRT-PCR验证,基因编号及表达量上调、下调结果见表4和图6。这14个基因在愈创木酚处理后发生不同程度的表达。与转录组测序结果相比,除FGSG_05882表达量由下调变为上调外,其余基因的表达量变化趋势与测序结果相符,表明转录组测序的分析结果是可靠的。
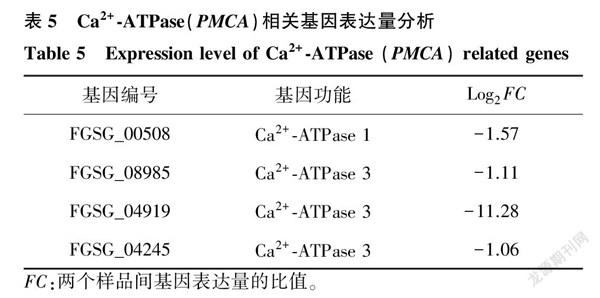
2.7Ca2+转运相关基因表达量分析
有研究发现,作为当归的有效活性成分,愈创木酚已被证实可以通过抑制破骨细胞中NF-κB、MAPK和AKT信号通路,抑制Ca2+外排,引起胞内Ca2+紊乱,影响破骨细胞的形成,最终达到治疗骨质疏松等疾病的目的[14]。在本研究中亦发现愈创木酚处理后Ca2+-ATPase 3(PMCA3)基因表达量显著下调,对测序结果中PMCA相关基因进行分析,发现
0.4 μl/ml愈创木酚处理禾谷镰刀菌后4个PMCA基因均表现为下调(表5)。利用qRT-PCR验证不同浓度愈创木酚处理后4个PMCA基因的表达情况,结果发现愈创木酚不同程度地抑制了这4个基因的表达,其中PMCA1基因(FGSG_00508)和2个PMCA3基因(FGSG_08985、FGSG_04919)的表达量整体上随着药剂处理浓度的升高而逐渐降低;另一个 PMCA3基因(FGSG_04245)在低浓度处理下表达量显著提高,随后又逐渐降低(图7)。
3讨论
近年来,通过比较药剂处理后基因表达差异情况可以分析药剂的潜在作用途径。已有研究结果表明,以愈创木酚为主要成分的各种木醋液对多种病原真菌均表现出一定的抑菌作用[15],但愈创木酚的作用机制尚不明确。本研究利用RNA-seq技术研究愈创木酚处理后禾谷镰刀菌基因表达的差异,旨在探寻愈创木酚的有效作用途径,为今后开发以愈创木酚为主要成分的禾谷镰刀菌杀菌剂提供理论依据。
测序结果表明,经过药剂处理后有905个基因表达量发生变化,其中表达差异倍数大于5的基因有65个,上调表达基因23个,下调表达基因42个,表明药剂处理后表达量变化较大的基因变化趋势以下调表达为主。已有研究结果表明,作为木质素生物油主要成分的愈创木酚具有很好的还原能力和自由基清除能力[16]。同时在调节植物生长方面,愈创木酚作为植物营养素的主要成分,可以清除活性氧,调节植物体氧化应激反应[17],愈创木酚亦作为牙科常用药物被验证具有较强的抗氧化作用[18]。在生物体中,谷胱甘肽的代谢途径与氧化应激反应相关,可以还原细胞内的活性氧自由基,生成氧化型谷胱甘肽来行使抗氧化的功能[19-22]。本研究发现,愈创木酚处理禾谷镰刀菌后,差异表达基因的GO功能主要富集在氧化还原相关生物进程上,Pathway 途径主要富集在谷胱甘肽代谢途径上,表明愈创木酚能够显著影响禾谷镰刀菌的氧化应激反应途径。
一般的植物源酚类化合物均具备较强的疏水性,可以通过破坏细胞膜的完整性进而影响细胞的生长[23],而当真菌受到外来药物作用时,细胞内的转运蛋白又可以将有毒有害物质运输至细胞外,保护真菌细胞的生长[24]。因此,用愈创木酚处理禾谷镰刀菌后,细胞膜组分及细胞运输相关的差异表达基因数量较多,且膜本体基因表达量显著下调,推测愈创木酚破坏了禾谷镰刀菌细胞膜的完整性,影响病原菌的生长。差异表达基因GO功能富集分析结果表明,愈创木酚处理禾谷镰刀菌后离子运输相关的差异表达基因较多,同时对表达差异基因进行验证时亦发现Ca2+-ATPase3基因(FGSG_04919)在药剂处理后表达量显著下调。在真菌细胞中,Ca2+作为第二信使影响了真菌的生长、细胞器的定位等一系列过程[25]。细胞内Ca2+浓度不平衡会导致细胞器内环境紊乱,内质网会出现蛋白质合成问题,而囊泡则会出现蛋白质降解问题,这些问题都会导致细胞凋亡[26]。真菌细胞中影响钙离子运輸的主要元件为膜上的钙离子运输ATP酶(PMCA)相关基因[27],对不同浓度愈创木酚处理后的禾谷镰刀菌的4个PMCA基因进行qRT-PCR分析验证,结果发现高浓度愈创木酚显著抑制此类基因的表达,由此推测愈创酚处理后PMCA基因表达量的下调破坏了细胞内Ca2+的平衡,进而引起细胞凋亡,影响病原菌的生长。
本研究中,我们在转录组水平上研究了愈创木酚对禾谷镰刀菌的作用机制,结果表明,愈创木酚能够通过影响禾谷镰刀菌的氧化应激反应途径、破坏细胞膜的完整性以及破坏细胞内Ca2+的稳态等方式来影响病原菌的生长,研究结果为进一步揭示愈创木酚作用的分子机制、挖掘作用靶标提供了参考。
参考文献:
[1]STARKEY D E, WARD T J, AOKI T,et al. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity[J]. Fungal Genetics and Biology,2007, 44(11):1191-1204.
[2]GULLINO M L, DAUGHTREY M L, GARIBALDI A, et al. Fusarium wilts of ornamental crops and their management[J].Crop Protection, 2015,73:50-59.
[3]CHEN Y,ZHOU M G. Characterization of Fusarium graminearum isolates resistant to both carbendazim and a new fungicide JS399-19[J]. Phytopathology,2009,99(4):441-446.
[4]CHEN C J,WANG J X,LUO Q Q, et al. Characterization and fitness of carbendazim-resistant strains of Fusarium graminearum (wheat scab)[J]. Pest Management Science, 2007, 63(12): 1201-1207.
[5]张琬琳,李艳,蔡祖善. 木材热解工艺的发展[J].林产工业, 2015, 42(12): 8-11.
[6]郭林林,张党权,谷振军,等. 樟树根材苯/醇提取物的Py—GC/MS分析[J]. 中南林业科技大学学报, 2011, 31(1): 142-147.
[7]杨宇,李妞,王爽. 愈创木酚的制备研究进展[J].工业催化, 2017, 25(4): 1-11.
[8]DE SOUZA ARAJO E, PIMENTA A S, FEIJ F M C, et al.Antibacterial and antifungal activities of pyroligneous acid from wood of Eucalyptus urograndis and Mimosa tenuiflora[J]. Journal of Applied Microbiology, 2018, 124(1): 85-96.
[9]NICULESCU O, GAIDAU C, SIMION D, et al. The Study on the possibility of using ecological materials with antifungal properties for treating natural leathers[J].Revista de Chimie,2020,71(1):445-449.
[10]TEMIZ A, AKBAS S, PANOV D, et al. Chemical composition and efficiency of bio-oil obtained from giant cane (Arundo donax L.) as a wood preservative[J]. Bio Resources, 2013, 8(2): 2084-2098.
[11]YANG J F, YANG C H , LIANG M T, et al. Chemical composition, antioxidant, and antibacterial activity of wood vinegar from Litchi chinensis[J]. Molecules, 2016, 21(9):1150.
[12]ROMERO-CORTES T, HUGO PREZ ESPAA V, ANTONIO LPEZ PREZ P,et al. Antifungal activity of vanilla juice and vanillin against Alternaria alternata[J]. CYTA: Journal of Food, 2019, 17(1): 375-383.
[13]张文云,张建诚,姚景珍. 氮胁迫下小麦叶片转录组分析[J].中国农业科技导报,2020,22(11):26-34.
[14]ZHI X, FANG C, GU Y, et al. Guaiacol suppresses osteoclastogenesis by blocking interactions of RANK with TRAF6 and C-Src and inhibiting NF-κB, MAPK and AKT pathways[J]. Journal of Cellular and Molecular Medicine, 2020,24(9): 5122-5134.
[15]刘润宇,叶乐夫,王雪,等. 木醋液在农业病虫草防控中的应用研究进展[J].中国农学通报,2020,36(35):113-118.
[16]CESARI L, MUTELET F, CANABADY-ROCHELLE L. Antioxidant properties of phenolic surrogates of lignin depolymerisation[J]. Industrial Crops and Products, 2019, 129: 480-487.
[17]JAYARAJ P, PARTHASARATHY S, RAJAGOPALAN S, et al. Discovery of novel synthetic hydroxyanisole derivatives as promising myeloperoxidase inhibitors (MPOIs) targeting atherosclerotic CVD[J]. Chem Med Chem, 2020,15(13): 1187-1199.
[18]吳补领,史俊南,王多宁. 几种牙科常用药物抗超氧化阴离子自由基的研究[J].牙体牙髓牙周病学杂志,1993(4):200-201,249.
[19]BURT S. Essential oils: their antibacterial properties and potential applications in foods—a review[J]. International Journal of Food Microbiology, 2004, 94(3): 223-253.
[20]张庆,魏树和,代惠萍,等. 硒对茶树镉毒害的缓解作用研究[J].南京林业大学学报(自然科学版),2020,44(1):200-204.
[21]陈玉胜,陈全战.一种功能性酸奶的研制及其抗氧化活性[J].江苏农业科学,2020,48(14):221-226.
[22]郑佩华,汪蕾,张秀霞,等. 凡纳滨对虾微粒体谷胱甘肽硫转移酶3基因克隆及其功能分析[J].南方农业学报,2020,51(10):2311-2320.
[23]REES D C, JOHNSON E, LEWINSON O. ABC transporters: the power to change[J]. Nature Reviews Molecular Cell Biology, 2009, 10(3): 218-227.
[24]MAILLOUX R J, MCBRIDE S L, HARPER M E. Unearthing the secrets of mitochondrial ROS and glutathione in bioenergetics[J]. Trends in Biochemical Sciences, 2013, 38(12): 592-602.
[25]HARRIS S D. Branching of fungal hyphae: regulation, mechanisms and comparison with other branching systems[J]. Mycologia, 2008, 100(6): 823-832.
[26]SHANAHAN C M, CROUTHAMEL M H, KAPUSTIN A, et al. Arterial calcification in chronic kidney disease: key roles for calcium and phosphate[J]. Circulation Research, 2011, 109(6): 697-711.
[27]MACHACA K. Ca2+ signaling and lipid transfer ‘pas a deuxat ER-PM contact sites orchestrate cell migration[J]. Cell Calcium, 2020, 89: 102226.
(责任编辑:陈海霞)
收稿日期:2021-05-26
基金项目:国家重点研发计划项目(2018YFE0206000);国家自然科学基金项目(31901936)
作者简介:张瑶 (1997-),女,江苏连云港人,硕士研究生,主要从事食品加工与安全方面的研究。(E-mail)1912682939@qq.com
通讯作者:马桂珍, (E-mail)guizhenma@sohu.com;史建荣, (Tel)025-84392001,(E-mail)shiji@ jaas.ac.cn
