施氮对杉木-浙江楠(Cunninghamia lanceolata× Phoebe chekiangensis)混栽苗木生长与植株元素含量的影响
2022-05-14杜佳奇王曼丽童佳丽王镜如朱国亮伊力塔
杜佳奇, 王曼丽, 童佳丽, 王镜如, 朱国亮, 伊力塔,*
施氮对杉木-浙江楠(Cunninghamia lanceolata× Phoebe chekiangensis)混栽苗木生长与植株元素含量的影响
杜佳奇1, 王曼丽1, 童佳丽1, 王镜如1, 朱国亮2, 伊力塔1,*
1. 浙江农林大学林业与生物技术学院, 浙江杭州 311300 2. 缙云县林业局, 浙江缙云 321400
为了探究氮沉降背景下杉木和浙江楠混栽苗木的生长特征, 于2019年5月模拟两种氮水平(对照0 kg·hm-2·yr-1·N、施氮45 kg·hm-2·yr-1·N)和三种混栽方式(每盆种植1颗杉木苗木、1颗浙江楠苗木, 地上竞争被隔板完全隔绝, 地下竞争由三种根系分隔方式控制竞争强度: 塑料膜分隔-根系无竞争、尼龙网分隔-根系干扰性竞争、无分隔-根系完全竞争), 2019年生长季(5月~12月)施氮处理并对杉木和浙江楠苗木生物量、根系形态和植株元素含量进行分析。结果表明: 根系竞争对杉木和浙江楠苗木的根、冠生物量无显著影响(>0.05)影响, 但两种苗木的根系生长随着竞争的加剧而受到抑制作用, 种间竞争特别是干扰性竞争能够促进植物对磷的吸收; 施氮后, 根系竞争促进了根、冠生物量的增加, 并对冠生物量的增加更有利, 同时, 根系干扰性竞争的促进作用最明显。另外, 两种苗木的根系生长随着竞争的加剧而受到促进作用。施氮和种间竞争均能促进植物对营养元素的吸收。
氮沉降; 生长; 植株元素含量; 杉木; 浙江楠
0 前言
杉木()是我国南方栽培最广的用材树种之一, 目前, 杉木纯林在生产经营中饱受地力衰退、生产力下降以及生态环境恶化等一系列问题[1]。采用针阔混交的方式是改善其人工林生态系统功能和服务的重要手段和途径[2]。浙江楠()作为我国特有珍稀树种, 不仅是常绿阔叶树种, 也是驰名中外的珍贵用材树种[3]。在生产实践中, 将杉木和浙江楠进行混交是一种有潜力的人工林发展模式[4]。
20世纪中叶以来, 随着人口膨胀和工农业的迅猛发展, 人为源活性氮输入逐年增加。我国氮沉降形势严峻, 已成为世界三大高氮沉降区之一。氮沉降导致土壤有效氮含量增加, 进一步增加了植物可利用氮含量, 并改变土壤pH和化学计量比等[1]。Lu et al[5]对中国的网络监测数据研究表明, 中国中南部是氮沉降的高发区域, 最高达到65 kg·hm-2·a-1, 当地林业产量受氮沉降影响颇深。杉木与浙江楠混交的人工林发展模式在生产实践中将不可避免地受到高氮沉降的影响。因此, 氮沉降条件下杉木与浙江楠的混交栽培研究受到了科研工作者的广泛关注。氮沉降背景下混交林生长受到不同程度的影响, 包括地上和地下的响应特征有所差异, 已有研究表明, 一定量的氮沉降可以促进混交林生物量和根系的生长, 且氮沉降会引起氮在楠木和杉木幼苗体内的氮含量增加, 对植株中其他元素的含量也产生不同程度的影响[6-7]。
竞争就是两个或两个以上的个体为争夺资源而发生的相互关系。植物个体间的竞争是一种很普遍现象, 彼此相邻的植物主要是为土壤资源而竞争[8]。植物根系间通过干扰式竞争和掠夺式竞争两种形式争夺土壤资源, 干扰性竞争是指植物根系直接依靠分泌对别的个体有抑制作用或表明其它竞争者存在的无毒信号等有机物而相互影响; 掠夺式竞争是指当根系间发生竞争时, 根系还可以直接和土壤相互作用而彼此间以间接方式联系[9]。前人的研究表明, 种间竞争可以提高混交林营养空间利用率, 增加林分的生产力, 但目前研究主要使针对地上部分[10], 对地下部分的研究较少。因此, 本研究将重点关注种间竞争对混交林根系生长、分布和适应能力的形状指标——根长、根系表面积、体积和平均直径[11]。
氮沉降和种间竞争是影响杉木和浙江楠人工林生产实践的重要因素, 氮沉降改变土壤资源背景, 使得种间竞争特别是地下部分根系的竞争态势发生改变, 并表现出不同的竞争策略[12], 根系的形态连同土壤养分的变化进一步改变植物植株对营养元素吸收。所以, 植物的生物量、根系形态以及植株营养元素的变化能够较为准确地反映植株应对环境变化的机理。因此, 本研究选取杉木及浙江楠苗木为对象开展混交和模拟氮沉降控制实验, 利用不同的分隔方式来代表种间竞争的程度。主要目的是: (1)探究施氮、种间竞争及两者交互作用对杉木和浙江楠苗木生物量的影响; (2)探究施氮、种间竞争及两者交互作用对杉木和浙江楠苗木根系生长及植株根系竞争策略的影响; (3)探究施氮、种间竞争及两者交互作用对杉木和浙江楠苗木植株元素含量的影响。
1 材料与方法
1.1 试验地概况
试验地位于浙江临安浙江农林大学东湖校区温室(119 ° 44 ′ E, 30 ° 16 ′ N), 属于中亚热带季风气候区, 气候温暖湿润, 光照充足。该地区年降水日约158 d, 多集中在春夏两季, 年降水量达1454.1 mm, 年平均气温为16.6 ℃, 年日照总时长1825.7 h, 无霜期历年平均237 d, 土壤为红黄壤[13]。
1.2 试验材料
试验土壤为当地红黄壤, 土壤养分值为: 全氮7.12 g·kg-1, 速效氮57.3 mg·kg-1, 速效钾71.0 mg·kg-1, 速效磷2.88 mg·kg-1, pH值5.18。试验用花盆直径30 cm, 高40 cm, 每花盆装入土壤6 kg。浙江楠幼苗来自浙江省丽水市庆元县永青林场良种繁育基地, 杉木幼苗为自行栽培的1年生实生苗。杉木幼苗的平均株高为3.6 cm, 平均冠幅都为3.8 cm; 浙江楠幼苗的平均株高为2.8 cm, 平均冠幅都为4.8 cm。2019年5月取长势一致的杉木、浙江楠幼苗各60株作为研究对象, 参照生产中针阔混交林的最优比例[3],在每盆混栽两种幼苗各1株。缓苗15天后移入对应的花盆中, 常规管理。
1.3 试验设计
试验为双因素完全随机试验。因素1为施氮浓度, 水平为(1)对照组, 施氮量0 kg·hm-2·yr-1; (2)施氮组, 施氮量45 kg·hm-2·yr-1(参考浙江省氮沉降本底值[14])。外源氮由NH4NO3和蒸馏水配置而成。将需要施氮的处理组NH4NO3溶解在15 L的蒸馏水中(水量以临安市近10月各月平均降水量为准, 平均分配到每周, 每周2次), 以电子喷雾器人工来回均匀喷洒在施氮组幼苗上, 对照组喷洒等量蒸馏水。
因素2为植株根系竞争程度, (水平植株地上部分均使用塑料板分隔, 以消除地上部分竞争对植株根系带来的影响)对照组和施氮组的地下部分区分以下3种种植方式: 采用尼龙网分隔(存在干扰性竞争)、塑料膜分隔(无竞争)或不分隔(干扰性竞争和掠夺性竞争共存)。共6种处理: 不施氮塑料膜、不施氮尼龙网、不施氮无分隔、施氮塑料膜、施氮尼龙网、施氮无分隔, 分别标记为I、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ。每种处理10个重复。
试验时间为2019年5月至12月, 缓苗结束后, 每隔3 d用电子喷雾器对施氮组进行NH4NO3溶液的施加, 同时每隔21 d进行株高、冠幅的测定, 每隔90 d收获30盆, 每个处理随机选择5盆, 测定其生物量、根系形态指标(根长、根系表面积、体积和平均直径)和植株的氮、磷、钾元素。
1.4 植物样品的收集与测定
根形态和生物量测定: 筛选施氮和不施氮条件下不同分隔处理的杉楠混交植株共60盆, 收获后将根系和地上部分分离, 使用根系扫描仪(Perfection V850 Pro, Epero Scannar)将根系扫描成电子图像后, 利用WinRHIZO根系分析系统(Regent Instruments Inc, Ottawa, ON Canada)测定根系的根长、根系表面积、体积和平均直径等各项形态参数后, 将其与新鲜地上部, 70℃, 48 h分别烘干到恒重, 测量干物质质量。
植物全株元素含量测定: 将烘干后的各处理植株粉碎过筛(100目), 分别称取0.3 g植物样品, 加入2 mL去离子水, 5 mL浓硫酸, 当样品转为棕黑色液体时, 分批次加入双氧水, 直至消煮液变为清亮色为止。将消煮液转移至100 mL容量瓶定容并过滤。植物全氮含量采用凯氏定氮法[15]进行测量, 量取10 mL过滤样至蒸馏管中蒸馏, 利用0.01 mol·L-1的硫酸溶液滴定。全磷采用钼锑抗比色法[15]进行测量, 量取2 mL过滤样显色定容到50 mL于880 nm处测定。全钾采用火焰光度计法[15]进行测定, 量取10 mL过滤样定容至50 mL于火焰光度计上测定含量。
1.5 数据处理
利用SPSS19.0 (SPSS Inc., Chicago, IL, USA) 软件对数据进行统计分析, 采用单因素方差分析(One-way ANOVA), 结合最小显著差异法(LSD)进行不同处理之间植株根系形态、生物量、全株元素的方差分析和多重比较(=0.05), 所有数据都满足方差齐性和分布正态性检验。图由Microsoft Excel 2010制作。
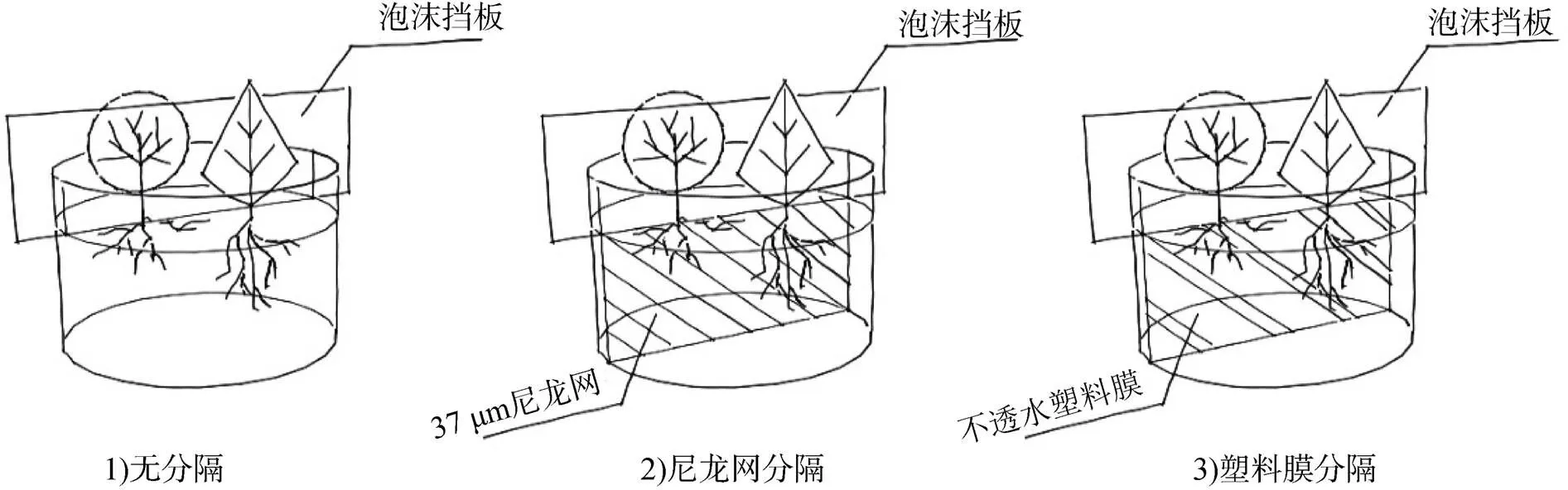
图1 三种混栽方式示意图
Figure 1 Schematic diagram of three mixed planting modes
2 结果与分析
2.1 施氮与不同根系分隔方式对杉木、浙江楠苗木生物量的影响
不施氮情况下: 杉木和浙江楠的根、冠生物量在各处理组上差异不显著(>0.05), 因此, 可以认为在不施氮条件下, 竞争对杉木和浙江楠根、冠的影响并不明显。
施氮情况下: 尼龙网分隔组(Ⅴ)的杉木冠生物量显著高于塑料分隔(Ⅳ)和无分隔组(Ⅵ)(<0.05), 由图1可以看出, 杉木的冠生物量随着竞争强度的变化呈现: 尼龙网>无分隔>塑料网。杉木的根生物量在各处理组上差异不显著, 但是根生物量趋势同冠生物量趋势一致。尼龙网分隔组(Ⅴ)的浙江楠根生物量显著高于塑料分隔组(Ⅳ)(<0.05), 尼龙网分隔组(Ⅴ)的浙江楠冠生物量显著高于塑料分隔(Ⅳ)和无分隔组(Ⅵ)(<0.05)。即, 在施氮情况下, 杉木和浙江楠的根、冠生物量随着竞争强度的变化呈现: 尼龙网>无分隔>塑料网, 与根、冠生物量产生明显相反趋势。
2.2 施氮与不同根系分隔方式对杉木、浙江楠苗木根系形态的影响
不施氮情况下: 塑料膜分隔组(Ⅰ)杉木幼苗的根系总长度显著高于无分隔组(Ⅲ)(<0.05), 根系表面积和根系体积则显著高于尼龙网分隔组(Ⅱ)及无分隔组(Ⅲ)(<0.05)。由于三种分隔带来的竞争抑制作用逐步加大(塑料膜>尼龙网>无分隔), 因此, 可以认为在不施氮情况下, 杉木幼苗根系生长呈现出随竞争加剧而受抑制的趋势。浙江楠幼苗根系在不施氮情况下, 各组均无明显差异, 不施氮情况下, 浙江楠幼苗对杉木带来的竞争胁迫并不敏感。

注: 不同大写字母表示杉木在不同处理间差异显著(p<0.05), 不同小写字母表示浙江楠在不同处理间差异显著(p<0.05)。
Figure 2 The root and crown biomass and root/shoot ratio of mixed seedlings ofand
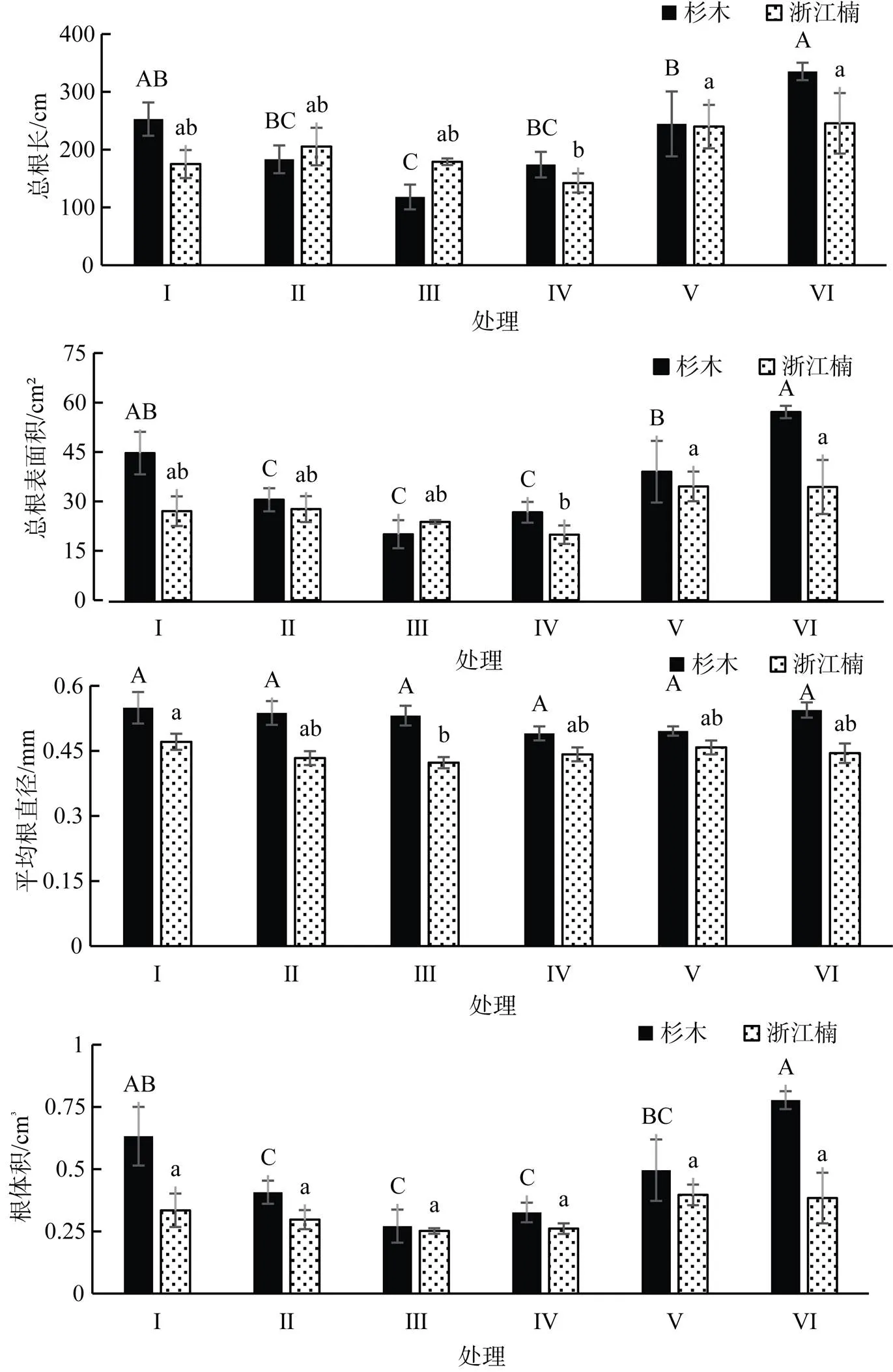
注: 不同大写字母表示杉木在不同处理间差异显著(p<0.05), 不同小写字母表示浙江楠在不同处理间差异显著(p<0.05)。
Figure 3 The root morphological characteristics ofand
施氮情况下: 塑料膜分隔组(Ⅳ)杉木幼苗的根系总长度、表面积及体积均显著低于无分隔组(Ⅵ)(<0.05), 由柱状图可以看出, 施氮情况下的杉木幼苗根系形态参数在不同竞争强度胁迫下, 与不施氮情况下产生明显相反趋势, 即: 施氮情况下, 杉木幼苗根系生长呈现出随竞争加剧而受促进的趋势。另外, 塑料膜组(Ⅳ)浙江楠幼苗根系总长度和表面积在施氮情况下, 显著低于尼龙网组(Ⅴ)和无分隔组(Ⅵ)(<0.05), 浙江楠幼苗根系生长也表现出随竞争加剧而受促进的趋势。
2.3 施氮与不同根系分隔方式对杉木、浙江楠苗木全株元素含量的影响
不施氮情况下: 由表1可以看出, 无分隔组(Ⅲ)的杉木幼苗的氮磷钾含量显著高于塑料膜分隔组(Ⅰ)(<0.05)。由于三种分隔带来的竞争促进吸收氮磷钾元素的作用逐步加大(塑料膜>尼龙网>无分隔), 因此, 可以认为在不施氮情况下, 杉木幼苗对氮磷钾的吸收能力随着种间竞争的加剧而逐渐提高。浙江楠幼苗在不施氮的情况下, 无分隔组(Ⅲ)的浙江楠幼苗的氮含量显著高于塑料膜分隔组(Ⅰ)(<0.05), 塑料膜分隔组(Ⅰ)的浙江楠幼苗的磷、钾含量显著高于无分隔组(Ⅲ)(<0.05), 浙江楠幼苗的氮含量表现出随竞争加剧而受促进的趋势, 而磷钾含量随竞争加剧而受抑制作用。
施氮情况下: 杉木幼苗的氮磷钾含量随着竞争的加剧而逐渐提高的趋势与不施氮情况一致, 而浙江楠幼苗在施氮情况下, 无分隔组(Ⅵ)的浙江楠幼苗氮、磷含量显著高于塑料膜分隔组(Ⅳ)(<0.05), 塑料膜分隔组(Ⅳ)的浙江楠幼苗的钾含量显著高于无分隔组(Ⅵ)(<0.05), 浙江楠幼苗的氮、钾含量随竞争变化的趋势与无施氮条件下一致, 而浙江楠幼苗磷元素含量却随着竞争加剧而受促进。
综上所述, 竞争的加剧促进杉木幼苗的氮磷钾元素和浙江楠幼苗氮元素的吸收, 而抑制浙江楠对磷、钾元素的吸收。施氮后, 总体趋势不变, 但浙江楠幼苗随着竞争加剧对磷元素的吸收的趋势与不施氮情况相反。另外, 施氮对杉木和浙江楠的氮元素吸收影响不明显。
3 讨论
在共享相同养分或资源的前提下, 不同植物之间将会发生竞争现象[16], 它们对竞争的响应机制也不同。适量的氮会增加植物的生物量、改变植物根系结构促进植物对养分的吸收等, 土壤养分分配较为均衡, 从而外界环境压力较小, 改变了物种个体间的竞争强度[17]。研究表明, 杉木和浙江楠对施氮和不同竞争强度及其交互作用均存在不同程度的响应。在本研究中, 在施氮情况下, 杉木和浙江楠的根、冠生物量随着竞争强度的变化呈现: 尼龙网>无分隔>塑料膜。在施氮情况下, 根系干扰性竞争促进杉木和浙江楠根、冠生物量增加, 而根系掠夺式竞争抑制杉木和浙江楠的根、冠生物量, 其中, 根系干扰式竞争带来的促进作用大于根系掠夺式竞争带来的抑制作用。根冠比反映了生物量在地上和地下部分之间分配的情况, 经常作为衡量植株生长状态特别是植株对土壤水分、养分状态反应的重要指标。杉木和浙江楠根冠比随竞争强度的变化呈现与根、冠生物量变化的相反的趋势: 塑料膜>无分隔>尼龙网, 说明施氮和竞争的交互作用能够影响生物量的分配, 根冠比随竞争的存在而下降是因为氮增加的同时增加了地上和地下的生物量, 但是地下生物量增加的量少于地上生物量。这与左有璐等[18]研究一致, 已有研究表明, 土壤状况良好的条件下, 植物将更多的生物量分配到获得外部资源的构件(地上部分)中, 这样有利于植物的繁衍和扩张。
竞争植物的生长形式, 特别是植物器官的形态能够影响其竞争能力, 根系特征是植物对土壤环境变化的适应能力指标之一, 对植物竞争的成败具有关键作用[19]。根系构型是植物根系生长和分枝的结果[19], 反映植物吸收、传导水分和养分的能力, 植物间竞争对根系最为明显的特征就是影响根系的分布[20-21]。植物根系的形态学特征对植物吸收土壤中有效性差、扩散速度慢的营养元素非常重要, 植物根系越发达、伸展越充分, 与土壤的接触范围就越大[22-23]。本研究结果表明, 不施氮情况下, 杉木幼苗根系生长呈现出随竞争加剧而受抑制的趋势, 而浙江楠幼苗对杉木带来的竞争胁迫并不敏感。
植株元素含量对植物生长起着至关重要的作用, 植物通过根系从土壤中吸收各所需元素和养分供应自身生长[24]。通过对植株元素的分析发现, 种间竞争对杉木和浙江楠对氮、磷、钾元素的吸收产生了影响, 种间竞争促进了杉木对氮、磷、钾元素的吸收, 种间竞争使土壤酶活性增加, 活化了土壤养分, 提高其有效性[25], 促进了杉木根系对这些营养元素的吸收, 同时也有研究发现, 在拟南芥的试验中, 增加硝酸盐的有效性会降低主根的长度[26], 因此杉木根系形态指标随着竞争加剧而受到抑制。另外, 种间竞争促进浙江楠对磷元素的吸收, 干扰性竞争使其根系向根际大量分泌有机酸, 通过螯合和络合作用或者酸化根际环境活化土壤中难溶性磷, 促进了它对磷元素的吸收, 但掠夺性竞争的存在又造成了浙江楠磷、钾元素的流失。俞元春[27]等研究发现, 缺磷胁迫下杉木和马尾松苗木根系有机酸的分泌量显著增加; 房莉[28]等研究发现低分子有机酸对根际土壤中难溶性磷活化作用较为显著; 这表明, 杉木和浙江楠在受到种间竞争威胁时, 会增加根系中有机酸含量, 其中主要为等低分子有机酸增量较多, 促进根际土壤中难溶性营养物质活化, 以提高林木根系的营养吸收。对于掠夺性竞争的具体机制尚不清楚, 有待于进一步研究。

表1 施氮与根系隔断对杉木与浙江楠全株元素含量的影响
注: 不同字母表示处理间差异显著性, 杉木与浙江楠分开标注显著性(单因素方差分析, LSD多重比较法,<0.05)。
大量研究表明, 施氮带来的土壤养分含量变化会影响外界压力变化从而影响植物根系竞争策略[29-30]在本研究中, 在对植物全株元素含量分析中, 可以进一步推测施氮将改变环境中的土壤养分水平, 增加环境资源, 改变竞争态势。齐玉春等[31]研究发现, 土壤有效氮含量随着施氮强度的增加而增加, 并且这种影响在土壤表层尤为显著。同时, 施氮显著增加了土壤有效磷含量, 且随着施氮含量的增加有效磷的含量也增加[31-32]。在资源丰富环境下, 高生长率物种占据竞争优势, 但当资源贫瘠时, 新陈代谢过快造成的营养损失不能够由正常的吸收得以弥补, 最终生长慢的植物根系生长会超过生长快的植物根系[33-34]。氮沉降将改变环境中的土壤养分水平, 增加环境资源, 在这种条件下, 速生树种如杉木与生长较慢的树种如浙江楠根系竞争态势将遭到改变, 甚至导致优劣互换[33]。
本研究还发现, 施氮促进了浙江楠对磷元素的吸收, 造成这种变化的原因可能主要有: 施氮使得土壤中速效磷的含量的增加, 从而促进了浙江楠对磷元素的吸收; 菌根能吸收土壤中的磷元素并传递给宿主植株; 凋落物的存在增加了土壤表层有效磷的含量。本实验结果表明, 浙江楠对磷元素的吸收的增加, 减少了种间竞争对浙江楠的影响, 但是浙江楠植株中磷元素增加的具体机制, 还需要进一步研究。并且根据庞丽[34]等研究发现模拟氮沉降显著增加了根际有机酸的分泌量, 提高了马尾松磷吸收效率。而施氮对杉木和浙江楠的氮元素吸收的影响并不明显, 可能是土壤中氮元素已经饱和的原因。另外, 由于本试验持续时间较短, 在施氮条件下, 杉木和浙江楠在不同的生长时期所表现出的生长情况可能会有一定差异, 这需要较长时期的试验进行研究。
4 结论
综上所述, 施氮和种间竞争的交互作用能够增加杉木和浙江楠根、冠生物量且对冠生物量增加更有利; 种间竞争能促进杉木和浙江楠对营养元素的吸收, 但却对杉木的根系生长起到了抑制作用, 施氮和种间竞争的交互作用能同时促进杉木和浙江楠对营养元素的吸收和根系的生长; 施氮将改变环境中的土壤养分水平, 增加环境资源, 改变竞争态势。因此, 本研究对于揭示施氮条件下, 杉木和浙江楠幼苗生长策略和种间竞争机制有重要意义。
[1] 毛晋花, 邢亚娟, 马宏宇, 等. 氮沉降对植物生长的影响研究进展[J]. 中国农学通报, 2017, 33(29): 42–48.
[2] 贾才锋. 浅谈杉木速生丰产的栽培技术及病虫害防治[J]. 农业与技术, 2015, 35(14): 116.
[3] 田晓, 刘苑秋, 魏晓华, 等. 模拟楠木杉木人工混交林不同混交比例对净生产力和碳储量的影响[J]. 江西农业大学学报, 2014, 36(1): 122–130.
[4] 邢海涛. 马尾松针阔混交人工林种间关系和作业法研究[D]. 北京: 中国林业科学研究院, 2017.
[5] LU Chaoqun, TIAN Hanqin. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data[J]. Journal of Geophysical Research, 2007, 112, doi:10.1029/2006jd007990.
[6] 史顺增, 熊德成, 冯建新, 等. 模拟氮沉降对杉木幼苗细根的生理生态影响[J]. 生态学报, 2017, 37(1): 74–83.
[7] 樊后保, 廖迎春, 刘文飞, 等. 模拟氮沉降对杉木幼苗养分平衡的影响[J]. 生态学报, 2011, 31(12): 3277–3284.
[8] 毛瑢, 曾德慧. 农林复合系统植物竞争研究进展[J]. 中国生态农业学报, 2009, 17(2): 379–386.
[9] WILSON S D, TILMAN D. Competitive responses of eight old field plant species in four environments[J]. Ecology, 1995, 76(4): 1169–1180.
[10] 徐思瑜, 陈圣贤, 陈雨清, 等. 氮添加对混栽杉木-楠木叶性状的影响[J]. 林业科学研究, 2020, 33(3): 184–192.
[11] 薛立, 傅静丹. 影响植物竞争的因子[J]. 中南林业科技大学学报, 2012, 32(2): 6–15.
[12] 黎磊, 周道玮. 红葱种群地上和地下构件的密度制约调节植物生态学报, 2011, 35(3): 284–293.
[13] FANG Xiaobo, SHI Jian, LIAO Xinfeng, et al. Heavy metal pollution characteristics and ecological risk analysis for soil inof Lin' an[J]. Chinese Journal of Applied Ecology, 2015, 26(6): 1883–1891.
[14] ZHAO Xu, YAN Xiaoyuan, XIONG Zhengqin, et al. Spatial and Temporal Variation of Inorganic Nitrogen Wet Deposition to the Yangtze River Delta Region, China [J]. Water, Air, and Soil Pollution, 2009, 203(1): 277–289.
[15] HESS T M. Tropical Soil Biology and Fertility: A Handbook of Methods[M]. Anderson JM, Ingram JSI. Wallingford: CAB International, 1990: 245–245.
[16] 李放, 乌云娜, 张小红, 等. 模拟氮沉降对克氏针茅和碱韭种间竞争的影响[J]. 中国草地学报, 2019, 41(2): 44– 50.
[17] 姜勇, 徐柱文, 王汝振, 等. 长期施肥和增水对半干旱草地土壤性质和植物性状的影响[J]. 应用生态学报, 2019, 30(7): 2470–2480.
[18] 左有璐, 王振孟, 习新强, 等. 川西北高寒草甸优势植物生物量分配对策[J]. 应用与环境生物学报, 2018, 24(6): 1195–1203.
[19] 南宏伟, 刘庆, 肖群英. 基于根系竞争的云杉幼苗细根内部异质性研究[J]. 山西农业科学, 2015, 43(5): 526– 531.
[20] 毛齐正, 杨喜田, 苗蕾. 植物根系构型的生态功能及其影响因素[J]. 河南科学, 2008, 26(2): 172–176.
[21] 向言词, 彭少麟, 彭秀花. 地下根竞3种木本植物幼苗生长的影响[J]. 热带亚热带植物学报, 2005, 2(2): 105–108.
[22] 高超, 李明思, 蓝明菊. 土壤局部湿润条件下棉花群株根系竞争生长研究[J]. 灌溉排水学报, 2016, 35(9): 26– 31.
[23] ANNA A, ZAIGHAM S. To respond or not to respond? Natural variation of root architectural responses to nutrient signals[J]. Journal of Experimental Botany, 2017, (11): 2636–2640.
[24] 春亮. 玉米氮高效品种选育及根系形态对低氮反应的遗传分析[D]. 北京: 中国农业大学, 2004.
[25] 王丹. 飞机草入侵过程中的种间竞争与协同作用研究[D]. 广州: 华南农业大学, 2016.
[26] 蒙好生, 冯娇银, 胡冬冬, 等. 植物根系发育与养分的吸收[J]. 山西农业科学, 2017, 45(6): 1048–1052.
[27] 俞元春, 余健, 房莉, 等. 缺磷马尾松和杉木苗根系有机酸的分泌[J]. 南京林业大学学报自然科学版, 2007, 31(2): 9–12.
[28] 房莉, 俞元春, 余健, 等. 低分子量有机酸对森林土壤磷[J]. 浙江林学院学报, 2007, 24(1): 28–32.
[29] JIANG Zhilin, LIU Wangxue, WAN Fanghao, et al. Competitive effects between(Asteraceae) and(Gramineae)[J]. Scientia Agricultura Sinica, 2008, 41(5): 1347–1354.
[30] 马杰, 易津, 皇甫超河, 等. 入侵植物黄顶菊与3种牧草竞争效应研究[J]. 西北植物学报, 2010, 30(5): 1020– 1028.
[31] 齐玉春, 彭琴, 云社, 等. 温带典型草原土壤总有机碳及溶解性有机碳对模拟氮沉降的响应[J]. 环境科学, 2014, 35(8): 3073–3082.
[32]赵河, 张志铭, 赵勇, 等. 模拟氮沉降对荆条灌木“肥岛”土壤养分的影响[J]. 生态学报, 2017, 37(18): 6014–6020.
[33] 王平, 王天慧, 周道玮, 等. 植物地上竞争与地下竞争研究进展[J]. 生态学报, 2007(8): 3489–3499.
[34] 刘延滨, 牟溥. 植物养分捕获的菌根塑性——外生菌根的塑性[J]. 植物生态学报, 2010, 34(12): 1472–1484.
[35] 庞丽, 张一, 周志春, 等. 模拟氮沉降对低磷胁迫下马尾松不同家系根系分泌和磷效率的影响[J]. 植物生态学报, 2014, 38(1): 27–35.
Effects of nitrogen addition on plant growth and element content of mixed seedlings of×
DU Jiaqi1, WANG Manli1, TONG Jiali1, WANG Jingru1, ZHU Guoliang2, YI Lita1,*
1. School of Forestry and Biotechnology, Zhejiang Agriculture and Forestry University, Hangzhou 311300, Zhejiang, China 2. Forestry Bureau of Jinyun City, Zhejiang Province, Jinyun 321400, Zhejiang, China
In order to investigate the growth characteristics ofandseedlings under the background of nitrogen(N) deposition, two N levels (0 kg·hm-2·yr-1·N and 45kg·hm-2·yr-1·N) and three mixed planting patterns (1and 1seedlings were cultivated in each pot; the above-ground competition was completely isolated by clapboard, and the underground competition was controlled by three separation modes: plastic film separation-no roots competition, nylon net partition-root interference competition, and no separation-root complete competition). During the growing season (June to December) in 2019, the biomass, root morphology and plant element content ofandseedlings were analyzed under N application. The results showed that rootcompetition hadno significant effect(>0.05) on the root and crown biomass of×seedlings. However, the root growth of these two kinds of seedlings was inhibited with the increase of competition. Interspecific competition, especially interference competition, could promote the absorption of phosphorus. After N application, root competition promoted the increase of root and crown biomass, and was more beneficial to the increase of crown biomass. At the same time, the promotion effect of root interference competition was the most obvious. In addition, the root growth of these two kinds ofseedlings was promoted with the increase of competition.Both N application and interspecific competition can promote the absorption of nutrient elements.
nitrogen deposition; growth; plant element content;;
杜佳奇, 王曼丽, 童佳丽,等. 施氮对杉木-浙江楠(×)混栽苗木生长与植株元素含量的影响[J]. 生态科学, 2022, 41(3): 214–221.
DU Jiaqi, WANG Manli, TONG Jiali, et al. Effects of nitrogen addition on plant growth and element content of mixed seedlings of×[J]. Ecological Science, 2022,41(3): 214–221.
10.14108/j.cnki.1008-8873.2022.03.025
Q945
A
1008-8873(2022)03-214-08
2020-07-17;
2020-08-29
国家自然科学基金项目(31971623)
杜佳奇(1999—), 女, 浙江绍兴人, 本科生, 生态学专业, E-mail: 240048723@qq.com
伊力塔, 男, 博士, 副教授, 主要从事森林生态学研究, E-mail: yilita@126.com
