A conceptual framework and an empirical test of complementarity and facilitation with respect to phosphorous uptake by plant species mixtures
2022-05-11ChunjieLIThomasKUYPERWopkeVANDERWERFJunlingZHANGHaigangLIFusuoZHANGandEllisHOFLLAND
Chunjie LIThomas W.KUYPERWopke VAN DER WERFJunling ZHANGHaigang LI*Fusuo ZHANG and Ellis HOFLLAND
1 College of Resources and Environmental Science,National Academy of Agriculture Green Development,China Agricultural University,Beijing 100193(China)
2 Wageningen University,Soil Biology Group,P.O.Box 47,6700 AA Wageningen(The Netherlands)
3 Wageningen University,Centre for Crop Systems Analysis,P.O.Box 430,6700 AK Wageningen(The Netherlands)
4 Inner Mongolia Key Laborator y of Soil Quality and Nutrient Resources,Key Laborator y of Grassland Resource(IMAU),Ministr y of Education,College of Grassland,Resources and Environment,Inner Mongolia Agricultural University,Hohhot 010018(China)
ABSTRACT Plant species have different traits for mobilizing sparingly soluble phosphorus(P)resources,which could potentially lead to overyielding in P uptake by plant species mixtures compared to monocultures due to higher P uptake as a result of resource(P)partitioning and facilitation.However,there is circumstantial evidence at best for overyielding as a result of these mechanisms.Overyielding(the outcome)is easily confused with underlying mechanisms because of unclear definitions.We aimed to define a conceptual framework to separate outcome from underlying mechanisms and test it for facilitation and complementarity with respect to P acquisition by three plant species combinations grown on four soils.Our conceptual framework describes both mechanisms of complementarity and facilitation and outcomes(overyielding of mixtures or no overyielding)depending on the competitive ability of the species to uptake the mobilized P.Millet/chickpea mixtures were grown in pots on two calcareous soils mixed with calcium-bound P(CaP)and phytate P(PhyP).Cabbage/faba bean mixtures were grown on both acid and neutral soils mixed with P-coated iron(hydr)oxide(FeP)and PhyP.Wheat/maize mixtures were grown on all four soils.Rhizosphere carboxylate concentration and acid phosphatase activity(mechanisms)as well as plant P uptake and biomass(outcome)were determined for monocultures rhizosphere and species mixtures.Facilitation of P uptake occurred in millet/chickpea mixtures on one calcareous soil.We found no indications for P acquisition from different P sources,neither in millet/chickpea,nor in cabbage/faba bean mixtures.Cabbage and faba bean on the neutral soil differed in rhizosphere acid phosphatase activity and carboxylate concentration,but showed no overyielding.Wheat and maize,with similar root exudates,showed overyielding(the observed P uptake being 22%higher than the expected P uptake)on one calcareous soil.We concluded that although differences in plant physiological traits(root exudates)provide necessary conditions for complementarity and facilitation with respect to P uptake from different P sources,they do not necessarily result in increased P uptake by species mixtures,because of the relative competitive ability of the mixed species.
Key Words: carboxylate,intercrop,phosphatase,phosphorus acquisition,phosphorus mobilization,phoshorus source,rhizosphere,root exudation
INTRODUCTION
Available phosphorous(P)is often inadequate to support crop yield because a large proportion of P is adsorbed to metal(hydr)oxides,precipitated as calcium phosphate,or bound to organic matter.Intercropping,the mixed cultivation of crop species in the same field(Vandermeer,1989;Willey,1990),may improve the use of P resources(Zhanget al.,2010;Fauconet al.,2015,2017;Messaoudiet al.,2020).However,the increase in P uptake may result in overyielding,defined as a greater than expected biomass of a species or a species mixture(Songet al.,2007;Latatiet al.,2014).The improved acquisition of P has been explained by two main mechanisms:facilitation and complementarity(Hinsingeret al.,2011).
Facilitation of P mobilization and acquisition is here defined as the process whereby one species enhances the uptake of the other through the excretion of molecules that increase P availability in soil.Facilitation therefore involves the combination of a nutrient-mobilizing species with a non-mobilizing neighbor(Liet al.,2014).For instance,facilitation can occur when legumes increase P availability to the benefit of cereals through rhizosphere modification(e.g.,acidification and secretion of carboxylate or phosphatases)when the roots of the intercropped species are nearby(Liet al.,2014;Fauconet al.,2015;Xueet al.,2016;Houassineet al.,2020;Wang and Lambers,2020).Enhanced P acquisition has been reported in cereal/legume intercropping,such as wheat/faba bean(Songet al.,2007),maize/chickpea(Heet al.,2013;Xiaet al.,2013),and maize/faba bean(Liet al.,2007)in field experiments.
Complementarity refers to niche differentiation or resource partitioning.It is defined as the process whereby each species mobilizes and subsequently acquires P from its“own”P pool,due to excretion of specific compounds in the rhizosphere.Resource partitioning could reduce interspecific competition because both species have unique traits,defined as heritable physiological abilities for P mobilization through different mechanisms(Fridley,2001;Brookeret al.,2016;Ducheneet al.,2017).Complementary acquisition of different forms of the same nutrient has been well demonstrated for nitrogen(N)in cereal/legume intercropping.The unique capacity of legumes to access atmospheric N(N2)through symbiotic fixation relaxes their competition for soil N with cereals(Hauggaard-Nielsenet al.,2001;Bedoussacet al.,2015).Complementarity for N could also occur if plant species differ in their preference for ammonium or nitrate or their preference for mineral N or organic N(amino acids).Turner(2008)hypothesized that complementarity for accessing different organic P(Po)pools could be a form of partitioning of different forms of soil Poin species mixtures.Hinsingeret al.(2011)extended this hypothesis to intercropping systems,in which two component species would tap into distinct soil P pools.Only a few empirical studies have investigated the uptake of P from different pools by different species through analyzing the depletion of different P pools by different species(Cuet al.,2005;Liet al.,2008).However,because P was not the limiting factor in these studies,the results were inconclusive with respect to the resource partitioning hypothesis.Currently,there is limited evidence for complementarity in P acquisition from different P sources in species mixtures,and it is not clear whether this complementarity leads to increased P uptake in such stands.
In a previous study,we tested for complementarity in P acquisition from different sources using pot experiments with inert quartz sand,where P resources were added separately,and there was no native P supply from the sand itself and no interaction of added P with the mineral phase(Liet al.,2019).Increased P uptake was observed for mixed P sources by one(millet/chickpea)of the crop species mixtures tested,compared to the average P uptake by the mixture from a sole P source(Liet al.,2019).That outcome suggested differential P acquisition from mixed P sources in the species mixtures and a reduced competition for P.Compared to this inert quartz sand,soils always contain a mixture of multiple P sources,and the desorption of P is essential for plant P acquisition in soil.Therefore,whether there is complementarity in P acquisition by species mixtures from different P sources in soil is still unclear.
Under our definition,facilitation is unidirectional and complementarity bidirectional.However,as both facilitation and complementarity can result in overyielding,they cannot be distinguished by the outcomes of the experiments.Because of this practical difficulty,they have been collectively referred to as the complementarity effect when quantitatively analysing the biodiversity effects on biomass production(Loreau and Hector,2001).Unlike N,there is no P form exclusively available to one species because P can only be taken up as orthophosphate(H2PO-4and HPO2-4)and,to the best of our knowledge,all plant transporters related to P uptake from the soil solution are orthophosphate transporters(Wanget al.,2017).A study using double-labelled(carbon(C)and P)phytate showed that enzymatic hydrolysis of phytate was essential for P uptake and that direct uptake of Powas negligible(Becket al.,1989).Once an unavailable form of P has been made available as orthophosphate,it can be taken up by all species.This common increased orthophosphate pool changes the competitive balance between species but this was not indicated in the model proposed by Hinsingeret al.(2011).
Based on the difference between complementarity and facilitation(bilateral and unilateral,respectively)and the consequences of enhanced orthophosphate availability for interspecific competition,we aimed to develop a conceptual framework for the model proposed by Hinsingeret al.(2011).This model should help interpreting the outcome(overyielding)of species mixtures in terms of mechanisms.As such,we assumed that Powas mobilized through the exudation of phosphatase,while sparingly soluble inorganic P(Pi)sources and metal(hydr)oxide-adsorbed P were mobilized through exudation of carboxylates.
We selected four low-P soils and three species combinations to test whether complementarity(as a mechanism)or facilitation of P uptake could lead to increased P uptake by species mixture.The soil selection comprised two high-pH soils,to which we added calcium-bound P(Ca-P)and Po(sodium phytate P,PhyP),and a neutral soil and an acid soil,to which we added metal(hydr)oxide-sorbed P(FeP)and Po(PhyP).Millet is commonly intercropped with legumes such as peanut(Feng L Set al.,2014)and soybean(Feng Y Zet al.,2014).In this study,we selected millet/chickpea because they differed in their abilities to uptake P from CaP and PhyP(Liet al.,2019).Similarly,we selected cabbage/faba bean because of their different abilities to mobilize P from FeP and PhyP(Liet al.,2019).The millet/chickpea and cabbage/faba bean mixtures were expected to uptake more P than the average of their monocultures due to the different P acquisition from different sources.Wheat and maize had similar abilities to uptake P from different sources(Liet al.,2019),and thus we expected no increase in P uptake in the wheat/maize mixtures.
MATERIALS AND METHODS
Conceptual framework
We developed a conceptual framework(Fig.1)based on a model by Hinsingeret al.(2011)to assess the mechanisms underlying the contribution of species interactions(competition,complementarity,or facilitation)to P uptake by mixtures after the addition of different,sparingly available P sources.Competition dominates the interaction when both species in the mixture effectively mobilize the same sparingly available P source and hardly mobilize the other P source,or when both species hardly mobilize both P sources and only compete for orthophosphate P in the soil solution(Fig.1a,d).Complementarity in accessing P from different sources dominates when each of the two species effectively mobilizes P from only one source and has a weak capacity to mobilize the alternative P source(Fig.1b,e).Facilitation dominates when one species in the mixture has a higher capability to mobilize one or both sparingly available P sources than the other,while the other species has a low capability to mobilize both P sources but benefits from the P mobilization by the first species(Fig.1c,f).This conceptual framework only describes the mechanisms of complementarity and facilitation.These mechanisms can result in two different outcomes,depending on whether the mixed species are equally strong competitors for orthophosphate P(Fig.1a,b,c)or whether one species is a stronger competitor(Fig.1d,e,f).In the latter case,there can both be overyielding and underyielding of P uptake and growth,depending on relative P acquisition gains by one species and P acquisition losses by the other.
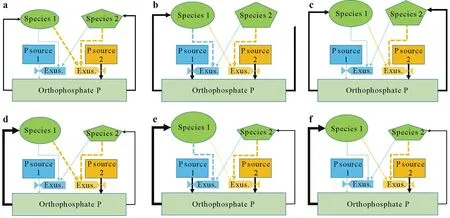
Fig.1 Conceptual framework of competition,complementarity,and facilitation of P acquisition.Two plant species(1 and 2)acquire and compete for P in the soil solution orthophosphate pool:both species effectively mobilize the same sparingly available P source and can hardly mobilize the P source,or both species can hardly mobilize both P sources and can only compete for orthophosphate P(competition,a and d);species 1 and 2 effectively mobilize P sources 1 and 2,respectively,and have a weak capacity to mobilize the alternative P source(complementarity,b and e),species 2 can effectively mobilize P source 2,while species 1 has a low capability to mobilize either P source(facilitation,c and f).In a,b and c,two species have equal competitive strength for P uptake.In d,e,and f,species 1 outcompetes species 2.Thin and thick lines represent weak and strong abilities to mobilize P sources,respectively.Exus.=root exudates.
Our conceptual framework differs from that of Hinsingeret al.(2011)in three major aspects.(i)It adds that there is competition for orthophosphate when roots are close enough to tap into the same orthophosphate pool,the only form of P that plants can take up.Competition is not mentioned in the framework of Hinsingeret al.(2011).(ii)It assumes plants can mobilize both P sources to some extent,although they may differ in the extent to which the mobilization occurs,while in the framework of Hinsingeret al.(2011),differential mobilization is absolute,where each species mobilizes only one P source in the case of complementarity or one species accesses two sources while the other accesses only one.(iii)It separates the mechanism from the outcome.In all cases,the addition of sparingly soluble P sources can equalize the competitive ability or result in competitive superiority(as competitive ability in a P-limiting soil is determined by acquisition of orthophosphate P,but not by the mobilization of sparingly soluble P).
Empirical test
Soils.Low P soils were collected from the top 20 cm of plots at experimental stations at four locations in China:Changping District in Beijing(39°59′N,116°17′E),Zhangye City in Gansu Province(38°85′N,100°38′E),Guangzhou City in Guangdong Province(23°10′N,114°16′E),and Kunming City in Yunnan Province(25°2′N,102°42′E)in China.Changping and Zhangye soils were both calcareous with loam and silt loam texture,respectively.Guangzhou soil was a typical acid red soil,and the Kunming soil was a neutral red soil.Texture of Guangzhou and Kunming soils was clayey.Soil P fractionation was determined using the P fractionation method of Tiessen and Moir(1993)(File S1,see Supplementary Material for File S1).Properties of the four soils are listed in Table I.After collection,soil samples were air-dried and sieved through a 2-mm mesh prior to potting.
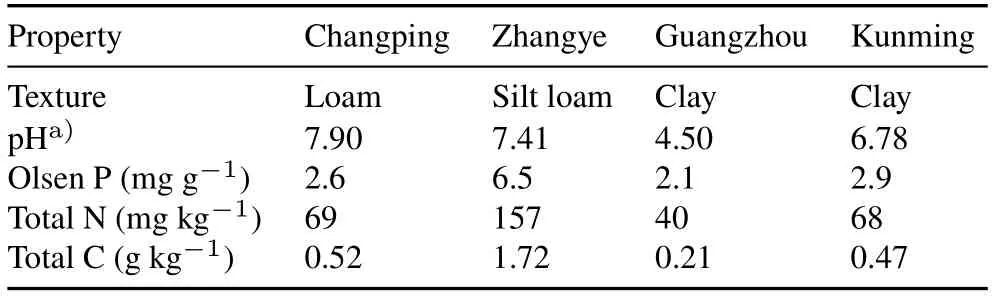
TABLE I Physical and chemical properties of the four soils collected from Changping,Zhangye,Guangzhou,and Kunming,China
Plant growth.A pot experiment was conducted from the end of September to December 2016in the glasshouse with natural light in China Agricultural University,Beijing.The temperature in the glasshouse ranged from minimally 13°Cduring the night to maximally 25°C during the day.Plants were grown in pots(160 mm in height,170 mm in diameter)containing 1 kg air-dried soil.The calcareous soils(Changping and Zhangye)were supplemented with hydroxyapatite(Ca5(PO4)3(OH),CaP,Shanghai National reagents,China)and sodium PhyP(C6H6O24P6Na12,Sigma,USA),both at 50 mg P kg-1soil as sparingly soluble P substrates under alkaline conditions.Kunming and Guangzhou soils were mixed with quartz sand(2:1,weight/weight)to prevent soil compaction and then supplemented with PhyP and P-coated iron(Fe)sludge(FeP)both at 50 mg P kg-1soil as sparingly soluble P substrates under acid conditions.The FeP was prepared by shaking(180 r min-1)6g Fe sludge(a byproduct of Brabant Water Ltd.,The Netherlands)in 170 mL of nutrient solution for 2 d at 25°C(Chardon et al.,2012).To support the initial growth of plants,KH2PO4was applied to each pot at 40 mg P kg-1soil.Other nutrients were mixed into each pot as follows:200 mg N as NH4NO3,226mg potassium(K)as K2SO4,126mg Ca as CaCl2·2H2O,39 mg magnesium(Mg)as MgSO4·7H2O,5.5 mg Fe as EDTAFe,6.7 mg manganese(Mn)as MnSO4·H2O,10 mg zinc(Zn)as ZnSO4·7H2O,2 mg copper(Cu)as CuSO4·5H2O,0.68 mg boron(B)as H3BO3,and 0.12 mg molybdenum(Mo)as(NH4)6Mo7O24·4H2O.
The crop species were millet(Setaria italicaL.,cv.Longgu-11),chickpea(Cicer arieti-numL.,cv.Longying-1),cabbage(Brassica oleraceaL.,cv.Sulv),faba bean(Vicia fabaL.,cv.Yundou-324),wheat(Triticum aestivumL.,cv.Kenong-9204),and maize(Zea maysL.,cv.Zhengdan-958).Monocultures and mixtures of millet/chickpea and wheat/maize were sown in Changping and Zhangye soils.Monocultures and mixtures of cabbage/faba bean and wheat/maize were sown in Guangzhou and Kunming soils.There were 24 treatment combinations in total,with four replicates arranged in a completely randomized design.
Before sowing,seeds of each species were surfacesterilized with 10%hydrogen peroxide(H2O2)for 30 min,rinsed thoroughly in deionized water,and pre-germinated on filter paper.The germinated seeds of millet were sown 2 weeks later than chickpea to prevent competition for light due to the rapid growth of millet.The seedlings of monocultures were thinned to six individuals for wheat,four for millet and chickpea,and two for cabbage,maize,and faba bean 7 d after sowing.There were two seedlings of each species in the millet/chickpea mixture,one seedling of each species in the cabbage/faba bean mixture,and three seedlings of wheat and one of maize in the wheat/maize mixture.The chickpea and faba bean seedlings were inoculated withMesorhizobium muleiense(the specific inoculant strain of chickpea)(Zhanget al.,2017)andRhizobium leguminosarum(the specific inoculant strain of faba bean)(Qiaoet al.,2015),respectively(provided by Culture Collection of Beijing Agricultural University,China).The germinated chickpea and faba bean seeds were soaked in a bacterial suspension for 30 min before sowing,and 5 mL inoculum was added to each pot.No nodules were formed on chickpea and faba bean in the experiment,most likely because considerable amounts of N were supplied to ensure that P was the unique limiting factor.Pots were watered with deionized water to 75%water holding capacity every 2 d.
Harvest and sample analysis.Millet and chickpea were harvested 51 d after sowing chickpea.Wheat and maize were harvested 47 d after sowing,and cabbage/faba bean 65 d after sowing when plant growth was slow.Shoots were cut at the soil surface,and roots were collected carefully.Roots with tightly adhering rhizosphere soil were immersed in 50 mL of 0.2 mmol L-1CaCl2solution and shaken carefully to extract rhizosphere soil.To prevent the degradation of carboxylates,we added the microbial inhibitor Micropur(Sicheres Trinkwasser,Germany)at 0.01 g L-1and three drops of concentrated phosphoric acid to a subsample of the rhizosphere extract(approximately 8 mL)before storing it at-20°C for high-performance liquid chromatography analysis,in which six carboxylates(tartrate,malate,citrate,succinate,fumarate,and trans-aconitate)were identified(Liet al.,2010).Another subsample,without addition of other chemicals,was stored at 4°C for determination of acid phosphatase(APase)activity within 48 h after sampling.The remaining rhizosphere suspension was air-dried to determine the rhizosphere soil dry weight(dw).
Because the solution:rhizosphere soil ratios differed depending on root size and amount of rhizosphere soil,we could not directly compare the absolute carboxylate concentrations in the extracts of different treatments.We therefore calculated the exudate ratios of each species when grown in the mixtures and when grown alone under two extreme assumptions.i)The soil solid phase is inert and does not buffer the carboxylate concentration.Under this assumption,the carboxylate concentration in the extract is a dilution of the rhizosphere solution concentration and can be expressed inμmol g-1rhizosphere soil dw.ii)The soil solid phase completely buffers the carboxylate concentration,such that the carboxylate concentration in the extract is the same as the concentration in the rhizosphere soil solution(expressed inμmol L-1).We then evaluated whether the ratios,calculated under both assumptions,showed an increase(if ratio values under both assumptions(mean±2×SE)>1)or decrease(if ratio values under both assumptions(mean±2×SE)<1)of carboxylate concentration as a response to a heterospecific neighbor.In other cases,we refer to the outcome as no increase or no decrease.We also calculated the ratio of carboxylates in the rhizosphere of one species over that of the other species to test for differences between the two species in the mixture.
We also determined rhizosphere pH.However,due to the different soil solution:rhizosphere soil ratios and different pH buffering capacities of the four soils,measured pH values were not necessarily representative of the rhizosphere pH and therefore are not shown.
Determination of rhizosphere APase activity was performed as described by Neumann(2006).The analysis involved colorimetric estimation of thep-nitrophenol(PNP)released by APase after incubation of soil with 4 mL of 0.04 mol L-1sodium maleate buffer(pH 5.3)at 28°C for 30 min.The reaction was stopped by 0.5 mol L-1NaOH and the absorbance was measured spectrophotometrically at 405 nm.One unit of APase activity was defined as the activity per gram soil that produced 1μmol PNP per hour.A similar evaluation procedure for carboxylates was followed for APase activity to determine whether there was an increase or decrease.
All harvested shoots and roots were washed with deionized water and then oven-dried at 70°C for 72 h before weighing.The shoot and root samples were ground and digested with HNO3-H2O2in a microwave accelerated reaction system(CEM,Matthews,USA).The P concentrations in the digests were determined by inductively coupled plasma optical emission spectroscopy(OPTIMA 7300 DV,Perkin-Elmer,USA).
Data analyses and statistical analysis
Total P uptake per plant was calculated by subtracting seed P content(Pseed)from the sum of shoot P content(Pshoot)and root P content(Proot)as follows:

One-way analysis of variance(ANOVA)was performed for biomass,P uptake,and tissue P content using R(version 3.1.2)(R Core Team,2014).All data were checked for normality prior to ANOVA and transformed when necessary.We used log10Xtransformation for data of maize root biomass in Kunming soil,and wheat shoot and root biomasses in Guangzhou soil.
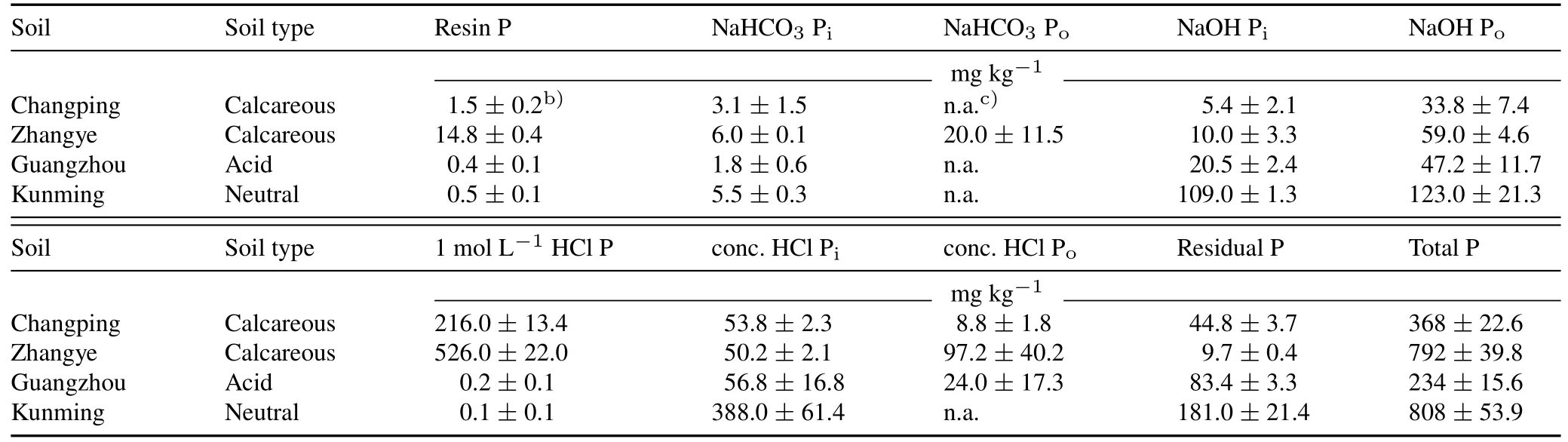
TABLE II Phosphorus fractions a)of four soils collected from Changping,Zhangye,Guangzhou,and Kunming,China
RESULTS
Soil P fractionation
Phosphorus content of the different P fractions varied among the four soils.Total P was relatively high in the calcareous Zhangye soil and the neutral Kunming soil.The largest P fractions of both calcareous soils were 1 mol L-1HCl Pi(59%—66%,Table II),representing Ca-bound P.Guangzhou and Kunming soils mainly contained occluded P and P held within Fe or aluminum(Al)(hydr)oxides,including residual P(22%—35%),NaOH-Pi(9%—13%),and concentrated HCl Pi(24%—48%),and recalcitrant Po.
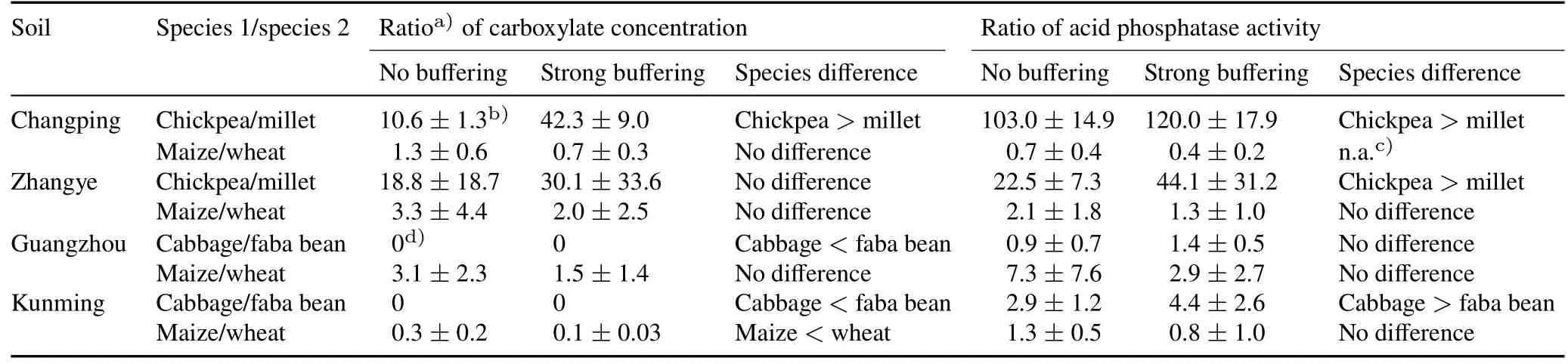
TABLE III Ratio of rhizosphere carboxylate concentration or acid phosphatase activity in the rhizosphere of one species to that of the other species in monoculture grown on four soils from Changping,Zhangye,Guangzhou,and Kunming,China
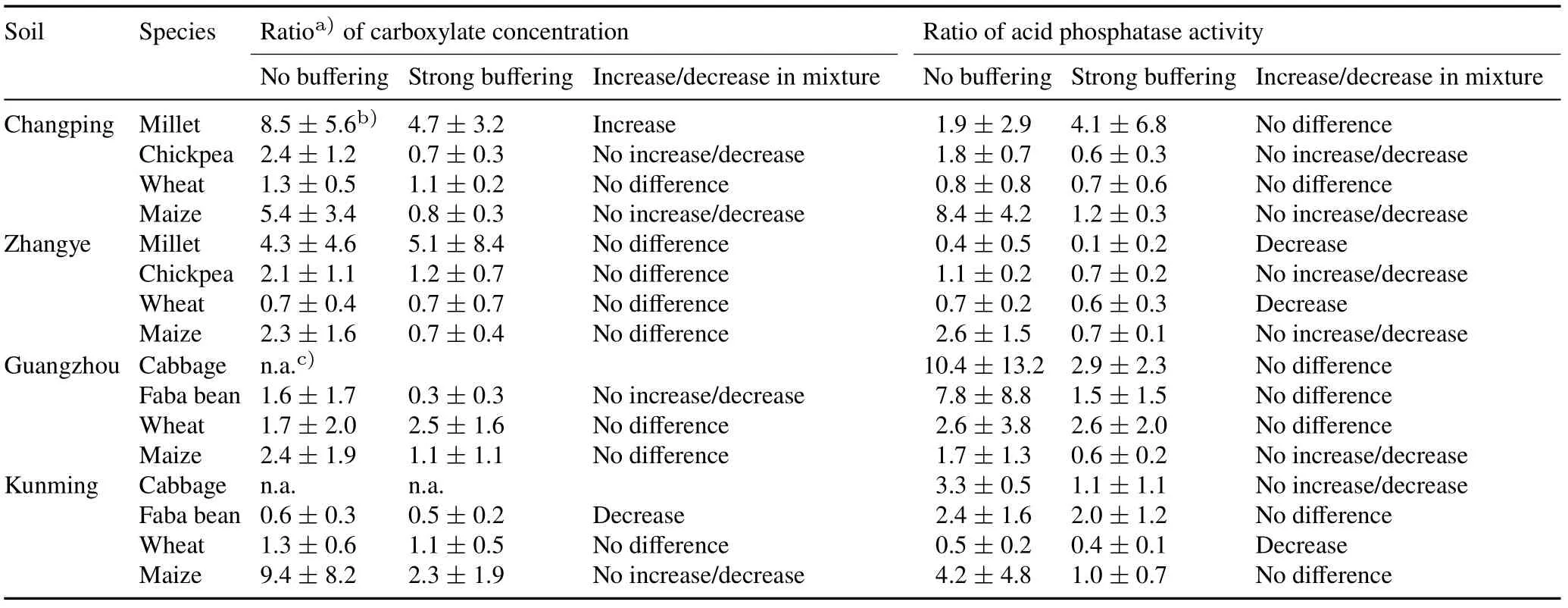
TABLE IV Ratio of rhizosphere carboxylate concentration or acid phosphatase activity of mixed species to monocultured species grown on four soils from Changping,Zhangye,Guangzhou,and Kunming,China
Species combinations on calcareous soils
Millet/chickpea.In Changping soil,both carboxylate exudation and APase activity in the rhizosphere were higher in monocultured chickpea than monocultured millet(Table III).While there was an increase in carboxylate exudation for mixed millet(the main carboxylates were malate,citrate,and succinate,Fig.S1,see Supplementary Material for Fig.S1)compared to the monocultured millet(the main carboxylates were malate and citrate,Fig.S1)(Table IV),the carboxylates were predominantly produced by chickpea(Table III).Phosphorus uptake by chickpea was around 0(Fig.2a),suggesting plant P content of chickpea was not significantly different from seed P(Eq.1);hence,no net uptake of P could be demonstrated.Shoot and root biomasses were similar in the monocultured and mixed chickpea(Fig.3a,e).Phosphorus uptake(Fig.2a)and shoot and root biomasses(Fig.3a,e)of mixed millet were 2.8,2.2,and 2.7 times higher than the monocultured millet,respectively.However,the increase in P uptake and biomass of mixed millet did not result in overyielding of the millet/chickpea mixtures(Fig.4a,e).The combined results on P uptake,biomass,and root exudation indicated facilitation(Fig.1f).
In Zhangye soil,APase activity of monocultured chickpea was higher than that of monocultured millet,but no difference in carboxylate exudation was found between them(Table III).Carboxylate exudations of both millet and chickpea were similar when grown in monoculture and in mixture(Table IV).Only the APase activity of mixed millet was lower than that of monocultured millet.Phosphorus uptake(Fig.2a)and biomasses of millet and chickpea(Fig.3a,e)were not affected by mixture on this soil.None of the mechanisms represented in Fig.1 matches these results.
Wheat/maize.In Changping soil,there was no difference in carboxylate exudation between monocultured wheat and monocultured maize(Table III),and there was no increase in the rhizosphere of wheat in response to mixing with maize(Table IV).The results did not allow for any conclusion on the modification of root exudation by maize as a response to mixing with wheat.Phosphorus uptake and shoot biomass of mixed wheat were 56%and 29%higher than those of monocultured wheat,respectively(P<0.01,Fig.2c,3c).In contrast,Puptake and shoot biomass of mixed maize were 89%(P<0.05,Fig.2c)and 31%(P<0.05,Fig.3c)lower than that of monocultured maize,respectively.There was no overyielding of biomass in the wheat/maize mixtures(Fig.4 g).However,the observed P uptake was 22%higher than the expected P uptake(Fig.4c).The outcome of mixing wheat and maize indicated unequal competitive ability(Fig.1d).
In Zhangye soil,there was no difference in exudation between monocultured wheat and monocultured maize(Table III)nor in carboxylate concentrations between both wheat and maize grown in monoculture or in mixture(Table IV).A decrease in APase activity was found only in the rhizosphere of wheat in response to mixing with maize(Table IV).Phosphorus uptake in mixed wheat increased by 56%compared to that of monocultured wheat(P<0.01,Fig.2c),while P uptake of mixed maize decreased by 44%compared to that of monocultured maize(P<0.01,Fig.2c).Shoot and root biomasses of wheat and maize were not affected by mixture(Fig.3c,g).No overyielding was found in P uptake or biomasses of wheat and maize mixtures(Fig.4).The outcome of mixing wheat and maize in Zhangye soil was in line with what we expected in the case of unequal competitive abilities(Fig.1d).
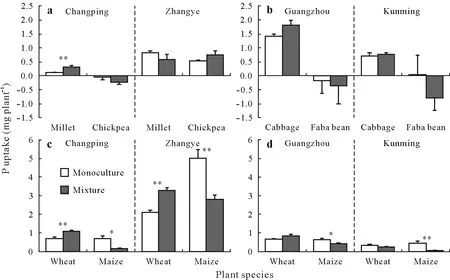
Fig.2 Phosphorus uptake of species in monocultures and species mixtures grown on four soils from Changping,Zhangye,Guangzhou,and Kunming,China:millet and chickpea on Changping and Zhangye soils(a),cabbage and faba bean on Guangzhou and Kunming soils(b),wheat and maize on Changping and Zhangye soils(c),and wheat and maize on Guangzhou and Kunming soils(d).Asterisks**and*indicate significant differences between monocultures and mixed species at P<0.01 and P<0.05,respectively.
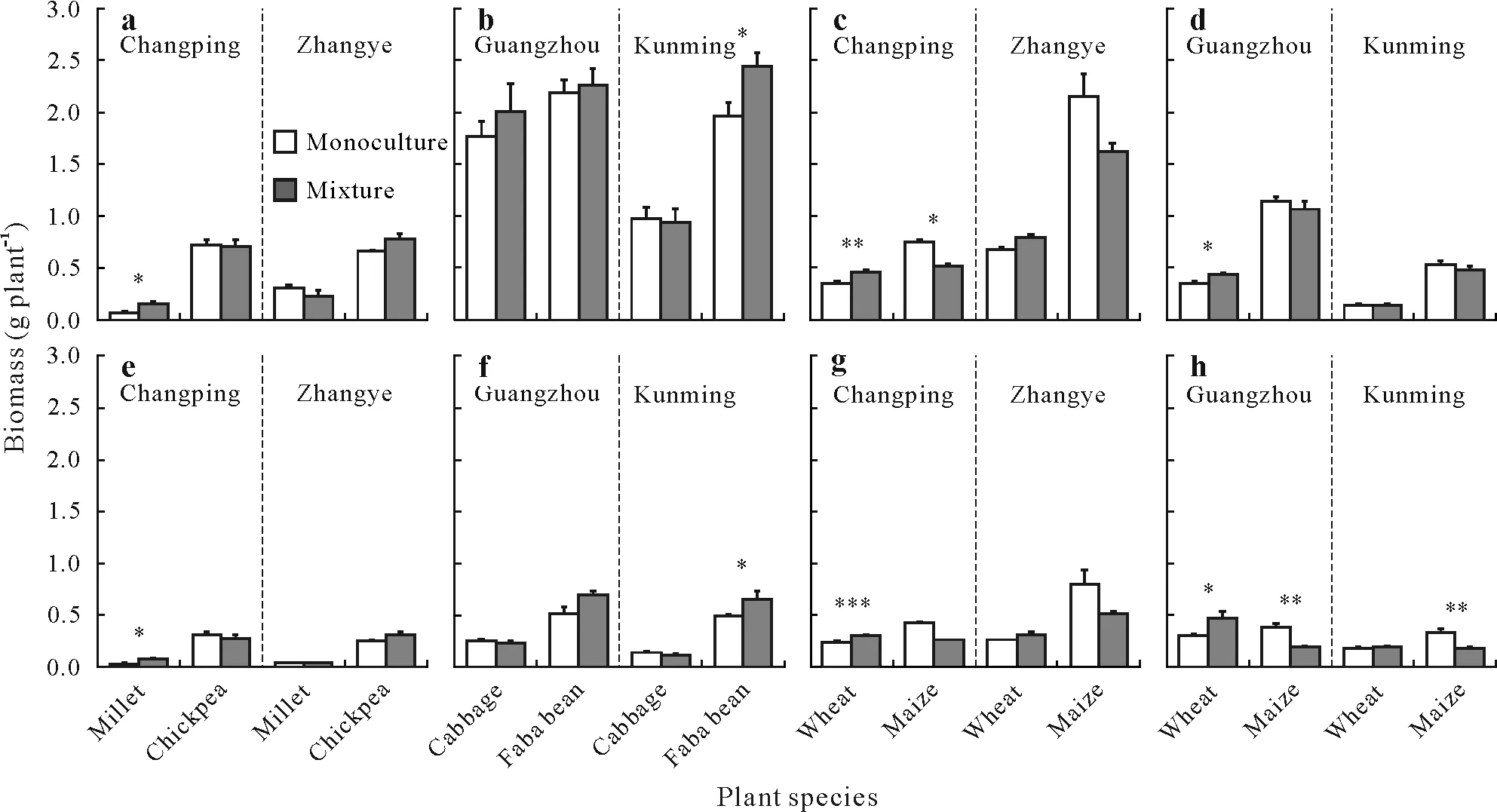
Fig.3 Shoot biomass(a—d)and root biomass(e—h)of species in monocultures and species mixtures grown on four soils from Changping,Zhangye,Guangzhou,and Kunming,China:millet and chickpea on Changping and Zhangye soils(a and e),cabbage and faba bean on Guangzhou and Kunming soils(b and f),wheat and maize on Changping and Zhangye soils(c and g),and wheat and maize on Guangzhou and Kunming soils(d and h).Asterisks**and*indicate significant differences between monocultures and mixed species at P<0.01 and P<0.05,respectively.

Fig.4 Expected and observed P uptake and biomass of species mixtures grown on four soils from Changping,Zhangye,Guangzhou,and Kunming,China:millet(species 1)and chickpea(species 2)on Changping and Zhangye soils(a and e);cabbage(species 1)and faba bean(species 2)on Guangzhou and Kunming soils(b and f),wheat(species 1)and maize(species 2)on Changping and Zhangye soils(c and g),and wheat(species 1)and maize(species 2)on Guangzhou and Kunming soils(d and h).The expected P uptake(or biomass)is the average of the P uptake(or biomass)of the species in monoculture.Asterisk*indicates a significant difference between expected(exp)and observed(obs)response variables at P<0.05.
Species combinations on acid and neutral soils
Cabbage/faba bean.In acid Guangzhou soil,carboxylate exudation by faba bean was higher than that by cabbage,but their rhizosphere APase activities were similar(Table III).No carboxylates were observed in the rhizosphere of either monocultured or mixed cabbage(Table IV).Mixing cabbage and faba bean had no effect on the APase activity of either species.There was no difference in P uptake or biomass between monocultured and mixed cabbage or faba bean(Fig.2b,3b).The difference in carboxylate exudation between cabbage and faba bean was consistent with facilitation(Fig.1c);however,there was no increase in carboxylate exudation by faba bean in the mixtures.In neutral Kunming soil,the rhizosphere carboxylate concentration of monocultured faba bean was higher than that of monocultured cabbage,while the rhizosphere APase activity of faba bean was lower than that of cabbage(Table III).There was a decrease in carboxylate exudation by faba bean in the mixtures(Table IV).However,APase activity was similar for mixed and monocultured faba bean.Carboxylate exudation was hardly observed in the rhizosphere of mixed or monocultured cabbage,and the response of APase activity of cabbage to mixing with faba bean was not clear.The uptake of P by both species was not affected by mixing(Fig.2b,3b,f).Shoot and root biomasses of mixed faba bean were 24%(P<0.05,Fig.3b)and 36%(P<0.05,Fig.3f)higher,respectively,compared to monocultured faba bean.Biomass of cabbage was not affected by mixing.Differences in rhizosphere parameters between the two species met the prerequisite for complementarity(Fig.1 b,c).Mixing cabbage and faba bean did not result in any overyielding of crop P uptake or biomass on either of the two soils(Fig.4b,f).
Wheat/maize.In acid Guangzhou soil,rhizosphere parameters were similar for wheat and maize(Table III)and were not affected by mixing(Table IV).We found both carboxylates and APase activity in their rhizospheres.Uptake of P was similar for mixed and monocultured wheat(Fig.2c).Shoot and root biomasses of mixed wheat increased by 23%and 52%,respectively(bothP<0.05,Fig.3d,h),compared to monocultured wheat.For maize,P uptake and root biomass of mixed maize were 33%(P<0.05,Fig.2d)and 50%(P<0.01,Fig.3 h),respectively,lower than that of monocultured maize.There was a difference(but not significant)between the observed and expected P uptake and biomasses of wheat/maize(Fig.4 h)in Guangzhou soil.These results indicated competitive inequality(Fig.1d).In neutral Kunming soil,APase activity was similar between monocultured wheat and monocultured maize,but carboxylate exudation of wheat was higher than that of maize(Table III).A decrease in APase activity was found in the rhizosphere as a result of mixing for wheat but not for maize(Table IV).Biomass was similar for monocultured and mixed wheat(Fig.3d).In contrast,P uptake decreased by 97%(P<0.01,Fig.2d)and root biomass decreased by 46%(P<0.01,Fig.3 g)for maize due to mixing.There was no significant difference in the observed and expected biomasses of wheat/maize(Fig.4d)in Kunming soil,but the observed P uptake by wheat/maize was significantly lower than expected(Fig.4d).These results indicated competitive inequality(Fig.1d).
37. Her girdle: In this instance, a girdle is a band of material around the waist that strengthens a skirt or trousers (WordNet). The girdle is not part of her underwear but an outer garment, often embroidered or decorated for women. The girdle would be one of her more valuable articles of clothing.Return to place in story.
DISCUSSION
We expanded a conceptual framework,originally proposed by Hinsingeret al.(2011),to test for rhizosphere mechanisms driving enhanced P uptake and overyielding as the outcomes of mixing species.In addition to the“P for Two”model of Hinsingeret al.(2011),our framework took into account that an increased pool of available orthophosphate P resulting from complementarity and facilitation could shift the competitive balance between the species mixed in a stand(Fig.1).We then analyzed the results of an experimental test for complementarity in mobilizing mixed P sources(resource partitioning)and facilitation of P uptake by mixing two species with differential P acquisition traits.We expected to find enhanced P uptake by species mixtures based on the different abilities of each species to mobilize different P forms.However,we found no evidence for complementarity in P acquisition from different P sources(resource partitioning)in any of the species combinations.We found that facilitation could have occurred in millet/chickpea combination in one calcareous soil(Changping soil,Fig.2a).We found differences between cabbage and faba bean in the ability to release carboxylate and APase on neutral soil(Kunming soil,Table III),but this difference in P mobilization traits did not lead to overyielding(Fig.2b).In contrast,acid phosphatase activity and carboxylate concentration of wheat and maize were similar(Tables III,IV)but there was overyielding of P uptake by wheat/maize mixtures on one calcareous soil(Fig.2c).
A prerequisite for testing whether complementarity or facilitation in P mobilization and acquisition drive overyielding is that plant growth is limited by P,but not by other factors.In all soils,Olsen P ranged between 2.1 and 6.5 mg kg-1(Table I),well below the criterion of 10 mg kg-1,often used to assess P deficiency.Plants were fertilized with all other essential nutrients in adequate amounts,further maintaining the status of the soils as P-limited.Further evidence for P limitation was found in the(very)low shoot P concentrations(mostly<2 mg g-1plant dw,Fig.S2,see Supplementary Material for Fig.S2),which were lower than the approximate critical shoot P concentration of these plant species(5.5 mg g-1plant dw for wheat,2.9 mg g-1plant dw for cabbage,2.4 mg g-1plant dw for chickpea,4.0 mg g-1plant dw for faba bean,and 2.7 mg g-1plant dw for maize)(Pearseet al.,2006;Wenet al.,2017).
In the current literature,complementarity is often used both as a cause(mechanism)of enhanced ecosystem functioning and as a result(outcome)of some community processes(Barryet al.,2019).The complementarity effect,calculated using the additive partitioning method(Loreau and Hector,2001),land equivalent ratio(LER),or relative yield total,represents the consequences but not the underlying mechanisms.Calculated on the basis of plant P content,LER is often used to demonstrate complementarity and facilitation with respect to P uptake and overyielding.However,several species interactions often occur simultaneously,resulting in enhanced resource acquisition and yield gain of intercrops.To avoid confusion,we separated the mechanisms and outcomes in our framework.
Our hypothesis on the complementarity of P acquisition from different P sources and enhanced P uptake by species mixtures was not confirmed.Species differences in root exudates were only found in one of the four hypothesized cases:in Kunming soil,faba bean rhizosphere carboxylate concentrations were higher than those of cabbage,while cabbage rhizosphere APase activity was higher than that of faba bean(Table III).This would suggest partitioning of metal(hydr)oxide-bound P and Po,and thus complementarity(as shown in Fig.1b,e).However,the expected resource partitioning by cabbage and faba bean did not result in extra P uptake by the species mixtures.The high APase activity of cabbage was probably a response to P deficiency(Tadanoet al.,1993).Few carboxylates were found in the cabbage rhizosphere in response to P deficiency,which was not consistent with a previous study(Dechassa and Schenk,2004).The lack of enhanced P uptake by cabbage/faba bean mixtures compared to monocultures could be due to low exudation of carboxylates,which did not release the phytate from FeP and other metal oxides in this strongbuffering soil(File S2,see Supplementary Material for File S2).The efficiency of APase may have been limited,as the mineralization of P from organic sources is limited by the availability of Poin the soil solution rather than by the concentration of phosphatases.Desorption of Pofrom metal(hydr)oxides through competitive adsorption of exuded carboxylates and/or acidification is essential(Tinker and Nye,2000;Georgeet al.,2005;Gerke,2015).
In Changping soil with a mixture of CaP and PhyP,we expected complementarity through partitioning of different P sources and enhanced P uptake by millet/chickpea mixtures based on differential P mobilization traits as in our previous study(Liet al.,2019).However,there was neither enhanced P uptake nor overyielding in species mixtures(Fig.4a,e);only millet benefited from mixing with wheat(Fig.2a).Increased P uptake and biomass of mixed millet compared to monocultured millet indicated enhanced competitive inequality due to the fast root growth of millet(Fig.3e).These results are consistent with previous studies showing that cereals benefit from mixing with legumes without affecting the legumes(Liet al.,2004;Cuet al.,2005;Dissanayakaet al.,2015).In line with previous studies(Tadano and Sakai,1991;Hayeset al.,1999;Neumann and Römheld,1999),both APase activity and carboxylate concentrations were higher in the rhizosphere of chickpea compared to millet(Table III).This is consistent with the fact that enhanced phosphatase activity is only effective in combination with enhanced dissolution or desorption of P(Hayeset al.,2000;Tinker and Nye,2000;Georgeet al.,2004).The rhizosphere of millet and chickpea likely overlapped in the mixtures,which might have facilitated millet P acquisition by chickpea.One of the two species(chickpea)was better able to mobilize sparingly soluble P sources and the other species(millet)benefited.This agrees with the unidirectional facilitation of millet by chickpea,followed by increased competitive ability of millet(Fig.1f).There was a small but significant increase in carboxylates of mixed millet compared to monocultured millet(Table IV),indicating that millet could mobilize Ca-bound P in the mixture.However,as the amount of carboxylates exuded in the mixtures by chickpea was much higher than that of millet(Table III),we considered that the upregulation of carboxylates by mixed millet was not very important.Because of the relatively poor growth of millet at low P availability on Changping soil(File S3,see Supplementary Material for File S3),the increased P uptake by millet did not lead to significant overyielding in millet/chickpea mixtures(Fig.4a).However,on Zhangye soil,millet and chickpea performed differently in terms of rhizosphere modifications,and there was no positive effect on P uptake due to species mixing.The immediately available P(resin P)of Zhangye soil was 10 times higher than that of the Changping soil(Table II).Shoot P concentrations(Fig.S2),P uptake(Fig.2a),and shoot biomass(Fig.3a)of both millet and chickpea were higher in Zhangye soil than Changping soil.The lack of facilitation of P uptake in millet/chickpea mixtures in Zhangye soil compared to Changping soil might be due to the higher soil P availability in Zhangye soil.
Consistent with our hypothesis,complementary P uptake did not occur in wheat/maize mixtures in calcareous soils.There was almost no difference in rhizosphere parameters between wheat and maize(Table III).Therefore,wheat and maize should have similar capacities to access the two P sources,consistent with our previous study(Liet al.,2019).In Changping soil,the wheat/maize mixtures took up more P than expected based on their respective monocultures(Fig.4a).This may be caused by competitive inequalities.Wheat is a stronger competitor for P than maize,because the root length density and phenotypic plasticity of wheat are larger than maize in intercropping(Liet al.,2001,2006;Liuet al.,2015).Assuming a low soil buffering capacity,there could have been an increase in root exudation by mixing maize(Table IV),which increased P availability and enhanced competition for P by mixed wheat at the expense of mixed maize.These enhanced competitive inequalities caused by increased P availability could also occur in the field but can be reduced by arranging the species with temporal niche differentiation in the mixtures(Liet al.,2020).In Zhangye soil with higher available P(Table II),we did not observe higher P uptake by wheat/maize mixtures compared to their respective monocultures.
For wheat/maize mixtures on acid and neutral soils,the growth and P uptake of wheat and maize were highly limited by P.The outcomes of mixing wheat and maize on these soils were consistent with the mechanism of competition,since wheat and maize showed similar rhizosphere exudation(Table III).These results were in line with our hypothesis.Field studies often showed overyielding by wheat/maize intercropping(Liet al.,2001).However,this overyielding is probably mainly due to temporal niche differentiation as a result of differences in sowing and harvesting time(Liet al.,2020).
In the present study,there were no indications that partitioning of P resources leads to overyielding by species mixtures,as predicted based on our earlier study(Liet al.,2019).We studied two rhizosphere mechanisms related to the differential mobilization of P from various sparingly available P sources.Several other mechanisms could also drive overyielding,including temporal and spatial complementarity in P uptake.In field studies with relay strip intercropping(Liet al.,2007;Songet al.,2007;Xiaet al.,2013),temporal complementarity in resource acquisition(e.g.,light,water,N,and P)is the dominant mechanism responsible for enhanced P uptake compared to that in monoculture.
Our expanded conceptual framework provides a tool to assess the potential role of co-occurring mechanisms contributing to the outcome of root interactions in species mixtures.Separating physiological mechanisms from outcomes(also driven by other root traits such as root morphology)helps to better analyze the contribution of rhizosphere modifications to P uptake in species mixtures.The mobilized P from sparingly soluble P pools goes into the same orthophosphate-P pool,which is available to both species(Fig.1).The increased P availability could enhance P uptake by the dominant species,thus increasing resource availability caused by facilitation and promoting species competition for resources(Holmgrenet al.,1997;Maestreet al.,2009;O’Brienet al.,2017).
It may be incorrect to infer mechanisms from outcomes because the absence of overyielding may coincide with facilitation.Alternatively,it may be incorrect to infer outcomes from mechanisms because overyielding can occur in case of competitive inequality(selection effect)(Loreau and Hector,2001).Our study therefore serves as a reminder that physiological root traits(exudation,an underlying mechanism for complementarity and facilitation)might be insufficient to result in overyielding,as the final outcome can also be influenced by morphological and architectural traits such as root length,diameter,or biomass(Sunet al.,2019),through which thin-rooted species may have the larger benefit of enhanced P mobilization.
CONCLUSIONS
We developed a conceptual framework that helps to relate outcomes of species mixing in terms of biomass and P uptake to underlying mechanisms,including rhizosphere processes and competition for orthophosphate P.The rhizosphere modifications partly supported the access to certain P sources and facilitation of P uptake in species mixtures.Under low P conditions,the increased P availability due to root exudation likely enhanced rather than decreased the competition for P by the dominant species in the mixtures because complementarity P use and facilitation do not necessarily result in increased P uptake by species mixtures.
ACKNOWLEDGEMENTS
This research was supported by the National Key R&D Program of China(Nos.2017YFD0200200 and 2017YFD-0200202),Projects of International Cooperation and Exchanges NSFC(No.31210103906),and Wageningen University Sandwich Ph.D.Fellowship(The Netherlands).We thank the anonymous reviewers for their helpful comments on this manuscript.
SUPPLEMENTARY MATERIAL
Supplementary material for this article can be found in the online version.
杂志排行

Pedosphere的其它文章
- Ultraviolet B radiation-mediated stress ethylene emission from rice plants is regulated by 1-aminocyclopropane-1-carboxylate deaminase-producing bacteria
- Drying-rewetting rather than sieving stimulates soil respiration
- Methanotrophy-driven accumulation of organic carbon in four paddy soils of Bangladesh
- Complementary effect of zoo compost with mineral nitrogen fertilisation increases wheat yield and nutrition in a low-nutrient soil
- Comparison of potential potassium leaching associated with organic and inorganic potassium sources in different arable soils in China
- Evaluation of immobilizing agents as soil quality conditioners in addition to their metal(loid)immobilizing effect