基于仿生学原理的改进KIII模型
2022-05-10陈玲钰田恬恬
张 锦,陈玲钰,田 森,刘 宏,田恬恬
1(湖南师范大学 信息科学与工程学院,长沙 410081)
2(湖南师范大学 数学与统计学院,长沙 410081)
3(郑州科技学院 信息工程学院,郑州 450064)
1 引 言
人类的大脑是一个非常复杂的功能结构.它包括听觉神经系统、味觉神经系统、视觉神经系统、触觉神经系统和嗅觉神经系统.其中嗅觉神经系统较为原始,信息处理没有时空结构,因此关于嗅觉神经系统的研究已经有着悠久的历史.多年来,关于嗅觉神经系统的研究已经发展出不同的模型来模拟嗅觉神经系统.
De Almeida等[1]构建了嗅球层到嗅皮层的多房室模型,用于研究输入到嗅球层和嗅皮层的胆碱能如何调节气味的表示.Zhang等[2]提出了一种嗅觉系统的前馈压缩传感模型.Barreiro等[3]提出了嗅球到梨状皮质通路的尖峰神经网络模型,研究在嗅觉信息加工过程中,结构内和结构间的突触是如何被调节的.Li和Cleland[4]针对嗅球试图构建一个通用的多房室模型,以生成影响僧帽细胞尖峰时间的伽马振荡.Reddy等[5]针对哺乳动物嗅感觉神经元对气味混合物的加工过程构建化学速率模型,研究不同气味在嗅感觉神经元上的竞争性拮抗作用.Viertel等[6]构建哺乳动物嗅球内外部簇状细胞的生物物理模型来研究其阵发性活动的发作机制.Ascione等[7]为研究嗅感觉神经元、球周细胞、僧帽细胞和颗粒细胞的放电活动,提出了一个分层模块化网络的随机模型来描述每种神经元的动力学行为.
以上提到的模型只模拟了部分嗅觉神经系统,而美国Freeman教授研究出的K系列模型从低到高层次逐步实现了整个嗅觉神经系统的模拟,包括嗅上皮、嗅球层和嗅皮层,模型的结构更加接近真实的嗅觉神经系统.K系列模型参数的设定建立在大量的神经生理学实验上,参数优化后,模型能够模拟脑电等神经活动,例如,模型产生非周期性振荡,具有类似1/f型的功率谱,峰值在伽马范围内,模拟出了类似嗅觉神经系统输出的脑电[8].
KIII模型是一种优秀的仿生模型.该模型不仅从多个方面对嗅觉神经系统进行了真实的模拟,包括模型的结构、神经元模型、神经元的功能特性等,而且还具有与深度学习模型相似的性能,即无需复杂的特征提取过程,即可直接识别脑电信号[9],这是KIII模型这一仿生模型非常独特的优点.
嗅觉神经系统到颞叶的神经通路如图1所示,整体分为3个部分:前段、中段和后段.本文针对前段嗅觉神经通路的关键部分进行模型研究,涉及嗅上皮、嗅球层和由前嗅核、梨状皮质组成的嗅皮层,以KIII模型作为前段嗅觉神经通路的基础模型,然后基于仿生学原理对KIII模型结构进行改进,并从结构上分析改进前后的KIII模型的小世界特性.

图1 简化的嗅觉神经通路图
本文的贡献如下:1)本文第一次基于仿生学的原理对KIII模型结构中前段的梨状皮质结构进行改进;2)本文基于小世界网络理论分析了改进前后KIII模型在结构上的小世界特性.
2 基于仿生学原理改进KIII模型
2.1 KIII模型
KIII模型的建立依据整个前段嗅觉神经通路的解剖结构.如图2所示,前段嗅觉神经通路主要是由嗅球(olfactory bulb,OB)、前嗅核(anterior olfactory nucleus,AON)和梨状皮质(prepyriform cell,PC)组成.每一个嗅感受器(R)的轴突即初级嗅神经(primary olfactory nerve,PON)都延伸到嗅球层的一个嗅小球,每个嗅小球大约聚集了4000万个嗅感受器的轴突,并且这些轴突之间没有连接.球周细胞(periglomerular cell,PG)对来自嗅感受器的信号进行预处理,嗅球中的僧帽细胞(mitral cell,M)接受嗅感受器和PG的输入.僧帽细胞是兴奋性神经元,颗粒细胞(granule cell,G)是抑制性神经元,两种类型的细胞通过负反馈相互连接形成振荡回路.嗅觉信息在嗅球层处理完毕后,由僧帽细胞通过外侧嗅束(lateral olfactory tract,LOT)传递给AON和PC.AON和PC中同样存在兴奋性神经元和抑制性神经元,分别为E和I、A和B.E和I、A和B相互连接形成类似于M和G的振荡回路.PC的输出从其锥体细胞到达外囊(external capsule,EC),也沿中嗅束(medial olfactory tract,MOT)向AON和OB发出反馈,而AON沿中嗅束向G和PG发送反馈信号.

图2 嗅觉神经系统的拓扑结构图
Freeman教授的K系列模型包括K0、KI、KII、KIII模型[10,11].其中,K0、KI和KII模型是KIII模型的基本组成单元,这是基于神经团理论建立的.神经团理论[12]是指相似的神经元组成的细胞团具有相似的功能和一致的特性,可以作为整个神经系统的组成模块.K0、KI、KII、KIII模型在不同的层次和规模上模拟了真实的嗅觉神经系统,其中,K0模型是K系列模型最基本的组成模块,表示相似神经元组成的具有相似功能和一致特性的细胞团.KIII模型主要由K0、KI、KII通过前馈、延时反馈等方式构成,模拟了整个嗅觉神经通路.这是关于嗅觉神经系统研究的突破性进展.
K系列模型的所有神经团可以用统一的公式来描述其动力学行为,具体描述见式(1)、式(2):
(1)

(2)
其中,N表示模型中的并行单元数;xi(t)、xj(t)分别是第i、j个神经团的电位状态变量;Wij表示第j个神经元到第i个神经团的突触连接强度;Ii(t)表示第i个神经团接受的外部输入;a和b分别表示神经电生理活动的两个时间常数,由生理实验得出a=0.220,b=0.720.Q(xj(t),qj)是一个由H-H(Hodgkin-Huxley)方程导出的非线性的S型输入/输出函数,q表示Sigmoid函数的最大渐近线.
K0、KI、KII模型通过前馈和延时反馈耦合在一起形成一个5层的KIII模型,用于模拟整个嗅觉神经系统.这5层分别是PG层、OB层、AON层、PC层和EC层,涵盖了从嗅上皮、嗅球层到嗅皮层的主要神经元、神经通路和突触连接等,如图3所示.
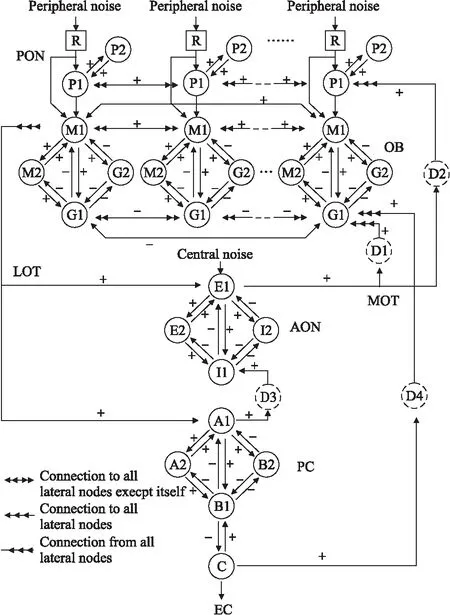
图3 KIII模型的拓扑结构图
2.2 改进后的KIII模型
尽管Freeman的KIII模型模拟了整个前段嗅觉神经通路,但是其重点在于对嗅球层的深入研究,简化了对梨状皮质的模拟,这部分的模型结构仅使用一个KI模型和一个KII模型来表示梨状皮质中神经元的连接情况.梨状皮质是嗅球层投射的最大区域,在气味信息处理、嗅觉产生的过程中起着非常重要的作用.为了进一步完善KIII模型,本小节将结合梨状皮质内部的细胞结构、神经回路,以及嗅上皮、嗅球层和嗅皮层的神经元比例关系,对KIII模型进行改进.
如图4(该图引用自文献[13])所示,梨状皮质主要分为第I层、第II层和第III层,其中第I层又分为Ia层和Ib层.第Ia层主要包含来自嗅球层的轴突,即传入纤维(afferent fibers,AFF),第Ib层主要包含梨状皮质中神经元的轴突,即联合纤维(association fibers,ASSN),也包含外部其他一些结构投射的连合纤维(commissural fibers).整个第I层还包含一些中间神经元,如水平细胞(horizontal cell)和神经胶质细胞(neurogliaform cell).第II层包含浅层锥体细胞和半月细胞,也包含一些中间神经元,例如双极细胞(bitufted cell,BC)和多极细胞(multipolar cell,MP).第III层包含深层锥体细胞和一些中间神经元(如多极细胞).浅层锥体细胞、深层锥体细胞和半月细胞属于兴奋性神经元,浅层锥体细胞和深层锥体细胞的顶树突、半月细胞的树突延伸至第I层,接受来自外侧嗅束和梨状皮质其他细胞的投射.但这两种锥体细胞和半月细胞的投射区域不同,两种锥体细胞的轴突可投射到梨状皮质以外的区域,例如嗅球、杏仁核、内嗅皮质等区域,而半月细胞的轴突仅能在梨状皮质内作用于两种锥体细胞的胞体[14].第I层的中间神经元属于抑制性神经元,作为梨状皮质内部前馈抑制(feedforward inhibition,FF)过程的中间导体,用于调节来自嗅球层的兴奋性输入,而第II层和III层的中间神经元介导PC内部的反馈抑制(feedback inhibition,FB).

图4 梨状皮质的主要细胞类型和解剖结构图
基于仿生学原理,本文提出了改进的KIII模型,如图5所示.与原KIII模型相比,改进的KIII模型遵循前段嗅觉神经通路的生理结构,包含这条神经通路的主要神经元,不同类型神经元的数量、比例和连接符合神经生理学数据,具体描述如下:

图5 改进的KIII模型拓扑结构图
1)涵盖前段嗅觉神经通路的关键部分
在改进的KIII模型中,PC层的关键部分分别对应于梨状皮质的不同部分,其中特别地,Ff对应于介导前馈抑制的中间神经元,Py对应于提供梨状皮质主要输入输出的锥体细胞,Fb对应于介导反馈抑制的中间神经元.其余部分则同原KIII模型一样:R对应于嗅上皮层中传入气味信息的嗅感受器;P对应于嗅皮层中对气味信息进行预处理的球周细胞;M和G分别对应于嗅球层中的兴奋性的僧帽细胞和抑制性的颗粒细胞,共同完成对气味信息的空间变换;E和I分别对应于前嗅核中的兴奋性神经元和抑制性神经元.
2)不同部分之间加入连接和延迟
模型中每一个R代表一类嗅感受器,投射到一个嗅小球,可直接投射或是通过P完成投射.Stettler等对梨状皮质关于气味反应的光学成像显示,梨状皮质中的每个神经元只对特定的气味有响应,来自嗅球层中的僧帽细胞的投射在空间分布上没有明显的偏好,呈现出不连续的投射域,不同的气味各自激活一个独特、离散的梨状皮质神经元集合[15].因此,可采用随机连接的方式模拟M到Py的投射过程.在PC层内部,Ff也接受来自M的投射,Ff的存在用于调节来自M的兴奋性输入,Py发出兴奋性连接,Fb发出抑制性连接,Py和Fb的局部互连形成振荡回路.PC层向AON层和OB层的反馈,由Pr分别投射到I和G.
3)神经元的数量遵循解剖学结果
对KIII模型的改进基于神经团理论进行,在PC层的输入部分,Ff采用分布式的KI(i)模型,Py和Fb的相互作用采用分布式的KII(ei)模型.相比于实际的嗅觉神经通路,模型中的神经元数量进行了缩减.嗅小球与梨状皮质锥体细胞的数量比例近似为1:1,模型PG层和OB层中的基本单元数会随着输入层R的数目改变.
3 实验结果
3.1 小世界网络理论
要想了解生物神经系统复杂的网络结构,可以通过解剖学知识,但更多的是借助相关的分析工具来分析神经系统结构背后所隐藏的一些特性.通常会使用图论来分析生物神经系统,将神经元表示为图的节点,神经元之间的连接表示为图的边,这个等效图类比神经系统的连接模式,进而研究等效图中所包含的某些特性.其中,小世界网络理论非常适合用于研究高聚类、紧密互连的稀疏网络.
为了量化网络结构的拓扑特征,早在1998年,Watts研究了一类平均路径长度较短,而聚类系数较大的网络,这种网络称为小世界网络[16](Small-World network).在这种网络中,有两个关键指标:反映全局连通性的平均路径长度L(Average path length,L)和衡量局部连通程度的聚类系数C(Clustering coefficient,C).对这两个指标的定义具体如下:假定一个有N个节点的图G,一个节点到另一个节点必须经过的最小边数称为两个节点之间的最短路径长度d,平均路径长度L指的是所有节点对的最短路径长度的平均值.节点i的平均路径长度见式(3),整个网络的平均路径长度见式(4).一个具有k个邻居的节点最多可有k(k-1)/2条边,计算出每个节点实际和邻居间存在的边数E与k(k-1)/2的比值,再对所有节点取该比值的平均值即为聚类系数C,节点i的聚类系数见式(5),整个网络的聚类系数见式(6).
(3)
(4)
(5)
(6)
判断一个网络是否具有小世界特性时,无法直接根据L和C的大小进行判断,通常先找出该网络对应的具有相同节点数、边数和平均度的等效随机图,并计算Lrand和Crand,见式(7)和式(8).若L≥Lrand且C>>Crand,则说明该网络具有小世界特性,这个判断方法称为小世界网络的半定量分类定义(semi-quantitative categorical definition)[16].
(7)
(8)
为了更加准确判断一个网络是否是小世界网络,Humphries等[17]给出了小世界网络的定量分类定义(quantitative categorical definition).首先,分别计算出给定网络的C与对应等效随机图Crand的比值γ,见式(9),给定网络的L与对应等效随机图Lrand的比值λ,见式(10),然后计算γ与λ的比值S,见式(11),若λ≥1且γ>>1,即S>1,则认为该网络是一个小世界网络.
(9)
(10)
(11)
3.2 小世界特性分析
本小节将对KIII模型改进前后的结构特性进行分析.首先,计算当输入通道数从5逐渐增加到100时,KIII模型在改进前后的平均路径长度L和聚类系数C的取值,其变化情况分别如图6和图7所示.原KIII模型的C值随着输入通道数增加而减小,改进KIII模型的C值随着输入通道数的增加而增大,并且两个模型C值的变化逐渐变得缓慢,分别收敛于某两个固定值.原KIII模型和改进KIII模型的L值均随着输入通道数的增加呈直线增大,从输入通道数大于16开始,在相同通道数的情况下,原KIII模型的L值均大于改进KIII模型,即原KIII模型的L值增长的幅度更大.输入通道数的增大意味着模型中神经元数目的增加.从两种模型的结构来看,原KIII模型的神经元数目主要集中在OB层,OB层到AON层、PC层和EC层的连接较少,而改进KIII模型扩展了PC层的神经元,增加了OB层到PC层的连接,由此也加强了模型内部的连通性,尤其是随着输入通道数的增加,更能体现出这个优势,使得改进KIII模型与原模型相比,有着较低L和较高的C.
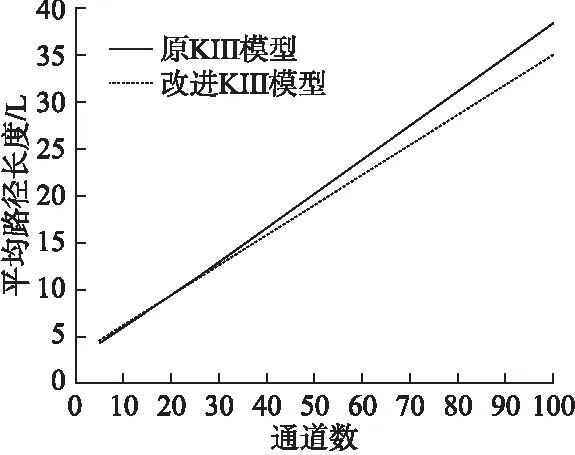
图6 原KIII模型和改进KIII模型的平均路径长度变化情况图

图7 原KIII模型和改进KIII模型的聚类系数变化情况图
接着,计算两个模型在输入通道数为10-100整十的情况下,对应等效随机图的L和C,并通过计算小世界特性统计量来分析两个模型的小世界特性,计算结果如表1和表2所示.由表可知,当输入通道数为10-100整十值时,对于每个通道数,两个模型均有λ>1,γ>>1,S>1.按照小世界网络的定量分类定义,在这些通道数对应的模型规模下,原KIII模型和改进KIII模型均具有小世界网络特性.这可以被认为是在改进前后用小世界网络理论评估KIII模型的一个有前途的方面.与此同时,当输入通道数为10-100整十值时,两个模型的λ相差比较小,随着输入通道数的增加,改进KIII模型的γ值迅速增长;且当输入通道数为20起(≤100且为整十值),在通道数相同时,改进KIII模型的γ远远大于原KIII模型的γ.我们的数据表明改进KIII模型较原KIII模型神经元之间连接更紧密,具有更强的小世界特性.

表1 原KIII模型在输入为10-100整十信道数时的小世界特性统计量表

表2 改进KIII模型在输入为10-100整十信道数时的小世界特性统计量表
4 结 论
本研究介绍了KIII模型,从结构上基于仿生学原理对KIII模型进行了改进,重点是对梨状皮质模拟部分进行了改进,并使用小世界网络理论分析了改进前后KIII模型的小世界特性.在实验中,计算了输入通道数为5-100时的平均路径长度和聚类系数的值.虽然改进前后的KIII模型均具有小世界特性,但在相同输入通道数下,改进后的KIII模型具有较大的聚类系数.当输入通道数超过16时,改进KIII模型的平均路径长度小于原始KIII模型;当输入通道数超过20时,改进的KIII模型的y值增长更快,且远远大于原KIII模型的y值.可以得出的主要结论是,改进的KIII模型比原KIII模型神经元之间的联系更紧密,具有更强的小世界特性.实验证明,基于仿生学原理对KIII模型的改进是有利的,达到了预期的目的.
KIII模型是一种优秀的嗅觉系统仿生模型,对比一般的深度学习模型,KIII模型更适合于嗅觉神经系统的模拟与研究.基于仿生学原理对其进行改进优化,使其具有更强的小世界特性,对模型的后续完善具有重要的意义.
