春芝麻娇驼跷蝽的发生与分布指标
2022-05-09任向辉赵庆杰崔建新余昊时文举
任向辉 赵庆杰 崔建新 余昊 时文举

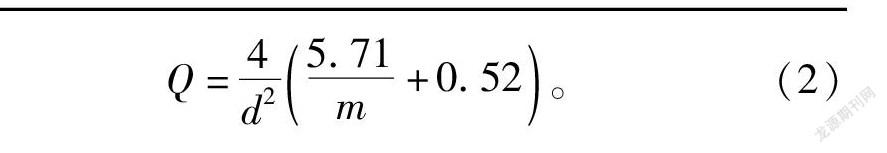
摘要:对3块春芝麻地的娇驼跷蝽进行2年不同月份密度、终花期空间分布以及寄主植物的株高、穗长、蒴果数、叶片均长等生态因子的调查,对该害虫6月发生量数据进行分布型指标计算与序贯抽样研究,结合相关环境因子数据进行了对应分析(CA)、R语言随机森林(RF)模型分析与发生因素的加权列联表分析。结果表明,此虫在春芝麻田为弱聚集分布型, M-M *回归方程为 M *=4.71+1.52 M ( r =0.92),其密度与月平均气温、降水量的二元线性回归方程为 y (密度)=14.82 x 1(均温)+4.31 x 2(降水量)-227.66( R =0.99)。对应分析与随机森林模型分析表明娇驼跷蝽在春芝麻植株上的个体数与植株株高负相关而与叶片长度呈正相关,借助软件绘出CA和RF分析的各因子二维效应图。此外加权列联表分析表明株高与叶长指标深度影响此虫发生水平,结合泡桐数据可看出此虫发生与环境关联度高,可借助其芝麻发生数据预测乔木发生密度。此虫食性杂、春季增长率惊人,特殊生境应警惕暴发性流行。
关键词:春芝麻;娇驼跷蝽;分布型;随机森林模型;加权列联表分析
中图分类号: S433.3;S435.653 文献标志码: A
文章编号:1002-1302(2022)07-0118-06
收稿日期:2021-06-27
基金项目:国家现代农业产业技术体系建设专项(编号:CARS-45-SYZ10);国家自然科学基金面上项目(编号:31272103);河南省大学生创新创业项目(编号:S201810467010);国家大学生创新创业项目(编号:20130515403)。
作者简介:任向辉(1967—),男,河南温县人,硕士,讲师,从事昆虫生理生态与害虫综合防治研究。E-mail:renxianghui@sina.com。
娇驼跷蝽[ Gampsocoris pulchellus (Dallas)]系危害兰考泡桐( Paulownia elongata S. Y. Hu)、木槿( Hibiscus syriacus Linn.)、蔷薇科(Rosaceae)果木、芝麻( Sesamum indicum Linn.)与大豆[ Glycine max (Linn.) Merr.]等寄主的全国广布性刺吸式害虫,其喜在芝麻果穗上聚集,木槿较多的蓉城成都春季它是随处可见的 “城市害虫”[1]。该虫生活史短、食性杂、春季增长率高且迁移距离远,体型细小、飞行快且天敌较多,种群年际动态规律复杂,所以它一般不被关注,近20年国内外研究文献几乎空白,笔者没查到它在寄主植物丰富且气候很适宜其生存的江苏地区的近期文献[2-4]。该虫是多种植病传播的媒介,试验表明其携带泡桐丛枝病类菌体后侵染辣椒( Capsicum annuum L.)苗致病时间达 31 d[5-7]。为充分利用空闲地和采收保健蔬菜芝麻叶菜,现在小片种植的春芝麻很多,将来随着中叶芝1号等叶用芝麻品种的推广春芝麻将更常见,此害虫的关注度将会提高。这些分散分布的春芝麻植株上娇驼跷蝽地面聚集点是研究该虫理想的数量生态学调查点,可开展增长型、分布格局、分布因素排序、序贯抽样以及预测建模等方面的研究。笔者针对这些内容在3块系统田进行了2年的相关数据调查,利用DPS平台计算了娇驼跷蝽最适抽样数以及该地田间防治指标,并进行了生态学对应分析(CA)、R语言随机森林(RF)建模分析和加权列联表预测,结合泡桐调查数据探讨了此虫暴发性发生的可能性以及利用芝麻地面调查数据预测乔木寄主发生量的技术可行性。
1 材料与方法
1.1 调查场地和试虫数据采集
调查场地选择河南科技学院校新东校区中被小麦( Triticum aestivum L.)与玉米( Zea mays L.)輪作大田分隔开来的有代表性的3块不相邻的春芝麻试验田,3块地光照和水肥条件比较好,面积为 5 000 m2 左右,栽培的品种为当地最常见的郑芝12号白芝麻。每年这些地块不套种其他作物,开春不打农药。3个地块周围的大田混有苘麻( Abutilon theophrasti Medicus)、牛筋草[ Eleusine indica (L.) Gaertn.]等杂草,便于娇驼跷蝽不同时间从1 000 m以外乔木寄主迁移过来栖息,此外每年各试验地块4月初不进行间苗操作且不打顶,所以6月芝麻植株丛较大且各个芝麻丛高度与成熟程度差异较大,适合做该虫相同生态背景下针对寄主植株不同生长指标的系统调查田[8-10]。
通过2020年4月初以来对新乡东郊方圆 6 000 m 内路边小块春芝麻地的调查,发现此虫迁移能力强所以各个地块密度近似,6月与7月初的发生高峰百株虫量都是4 000头左右。将3块地采用棋盘格取样法分散取30个点,于2020年4—7月的1个春芝麻完整生长期与2021年4—6月的1个与泡桐正比例同步增长阶段,分别进行每月1次100株1点的百株虫量调查,以推算每株发生均值。春季调查时此虫为外迁而来的第1代新发个体或越冬复苏个体,因此统计寄主植株上附着的成虫与极少量稚虫数。该虫对芝麻有强趋性所以可对芝麻植株各部分直接准确计数,但因该虫飞行快且泡桐植株高大,对泡桐只能采用细眼捕虫网快速对个别低矮枝梢罩头的方法计数,约1/4虫子逃逸所以所得数据为枝梢的发生量局部数据,每次取统计学大样本40个低矮枝梢进行调查。淇县北阳镇史庄近千亩(1 hm2=15亩)连片桐麦间作的国道附近泡桐枝梢3次计数与2020年9月校东区夏芝麻的娇驼跷蝽均值调查也进行对比分析。害虫发生与寄主生长指标关系的调查选择在春芝麻株形较大且叶、花、果俱现的2021年6月上旬终花期进行,共进行了12个样方60个点的调查,1~4、5~8、9~12号样方所在的3块地各20个点[9-11]。调查时发现有极少量绿色的锤胁跷蝽[ Yemma signatus (Hsiao)]与娇驼跷蝽混合危害,计数时对其不予统计[9]。
由于6月上旬初步调查时发现生长高度与花序成熟程度相似的芝麻株发生量十分接近,为进一步保证数据的稳定性,每个计数点芝麻丛选取10株高低、枝叶色泽与花序形态相似的相邻芝麻植株进行。取这10株芝麻的株高、穗长、蒴果数、叶片平均长度、所附着的娇驼跷蝽等5个因素数据的平均数作为以后排序分析时1个调查点的5个变量值。每个样方中心点取1个芝麻丛,在此调查点周围采用五点取样法再选取4个沿90°角度方向延伸3 m处的5个植株丛的调查点[11-13]。
1.2 各月百株虫量与气候相关度计算
查阅中国天气网、新乡市气象局官网的2020年农业气象周报与月报,用Excel绘制春芝麻完整生长期4个月的发生量数据与月均温、月降水量等因素柱形图,利用SigmaPlot软件进行二元线性回归计算与3-D回归效果图绘制并计算相关度 R 值[11-13]。
1.3 聚集度指标计算与理论抽样数确定
利用DPS 18.1的单因素方差分析模块判断2021年6月上旬3田块发生量数据的差异性,将各样方因变量数值均值与方差输入DPS 18.1进行分布型参数与相关回归方程的计算,采用DPS分布格局模块的默认有效数字位数。分析指标为DPS自动生成的平均拥挤度( M*指标)、聚集度指数(M*/M指标)、丛生指数(I指标)、K指标与CA 指标等聚集度指標,其含义见相关文献[11]。此外DPS还给出了Taylor幂法则回归方程和 M*-M 回归IWAO方程。根据IWAO方程的 α与β确定序贯抽样模型不同允许误差与不同虫口密度条件下的最适抽样数Q,利用防治阈值x o、α与β 这3个参数采用IWAO提出的序贯抽样模型( D=nx 0±tn[(α+1)x 0+(β-1)x2 0])确定抽样数n 时的防治界限[11-13]。
1.4 CA分析与随机森林建模分析方法
利用生态学软件CONOCO对2021年终花期调查得到的4个环境因子自变量和1个害虫发生数量因变量进行CA分析(对应分析),分析前将4个自变量数据的每个数值进行减平均值并除方差的常规无量纲化转换,所有相关关系计算采用CONOCO默认算法。环境因子与生物因子之间的二维排序图由CONOCO根据CA分析所得到的各个变量的相关矩阵自动绘出[11-13]。将无量纲转换后的环境变量与发生量数据作CSV格式Excel输入R软件后运行美国加州大学伯克利分校Andy Liaw 和Matthew Wiener开发的randomForest程序包进行RF回归分析模型建模,模型决策树数目设定为1 000,运行varImpPlot与partialPlot命令进行环境因素的排序图与偏效应图绘制[14-15]。
1.5 加权列联表预测
初步分析表明,对于迁飞性微小害虫娇驼跷蝽来说,栖息环境条件就大致决定了其分布密度,对15%~85%的本次调查点在终花期此虫数量为每株30~40头数值的中间水平级别。应用DPS软件的scale命令与ffx命令对终花期调查的所有变量数据进行分布样本数在[0~15%]、(15%~85%)、[85%~100%]等3个级别的分级分类数据变换,然后输入DPS加权列联表分析模块进行发生级别预测。将第1个样方的1~5号样点数据做预测样本,对 6~60号样点数据进行5因素3水平的加权列联表分析。计算得到的4个自变量的随机系数矩阵与预测样本的自变量数据矩阵相乘,则可以得到预测值与四舍五入后的具体预测水平。运行SigmaPlot软件绘制出这55个样点带95%置信区间的发生水平与预测值的线性相关图[11-13,15]。
2 结果与分析
2.1 百株虫量与气候相关度分析结果
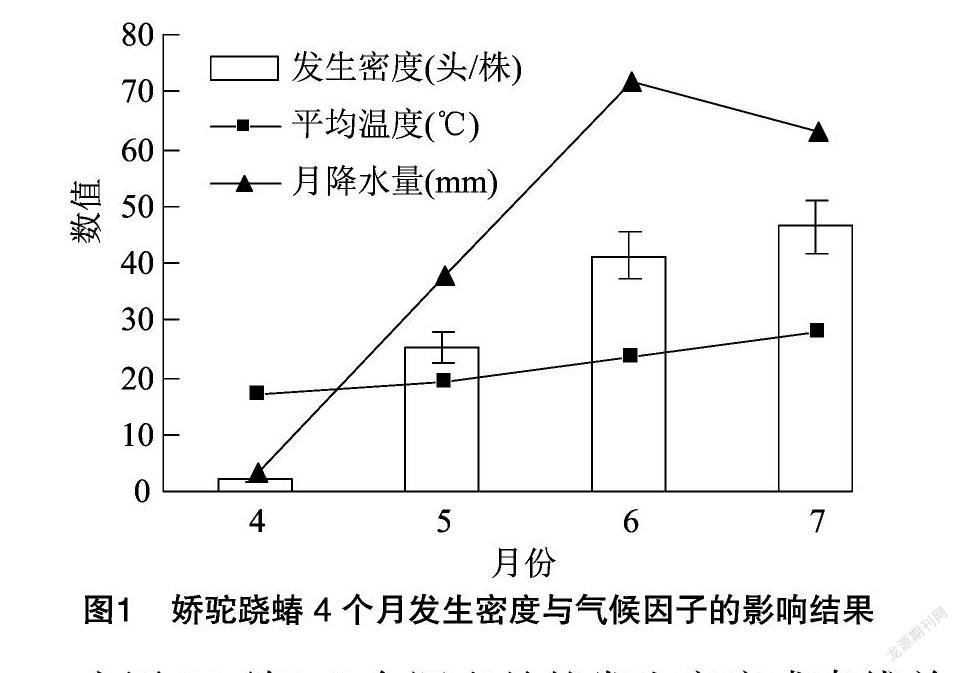
2020年30个样点百株虫量4个月的调查结果依次为199.0、2 520.7、4 112.7、4 643.4头,换算为每株虫量为1.99、25.21、41.13、46.43头。4个月的每株虫量及其误差范围以及月均温(℃)与降水量(mm)用Excel绘制带折线的柱形图见图1。
由图1可知,4个调查月的发生密度成直线关系增长,这和文献中春夏季泡桐苗圃娇驼跷蝽发生密度的月增长比例是一致的[6]。将温度与降水量分别与发生量进行一元线性回归发现 R 值都大于0.85。将3列数据输入SigmaPlot软件进行发生量与环境因子的二元线性回归可以得到更精准的回归结果,计算得回归方程为
y=1.48x 1+0.43x 2-22.77(R=0.99)。(1)
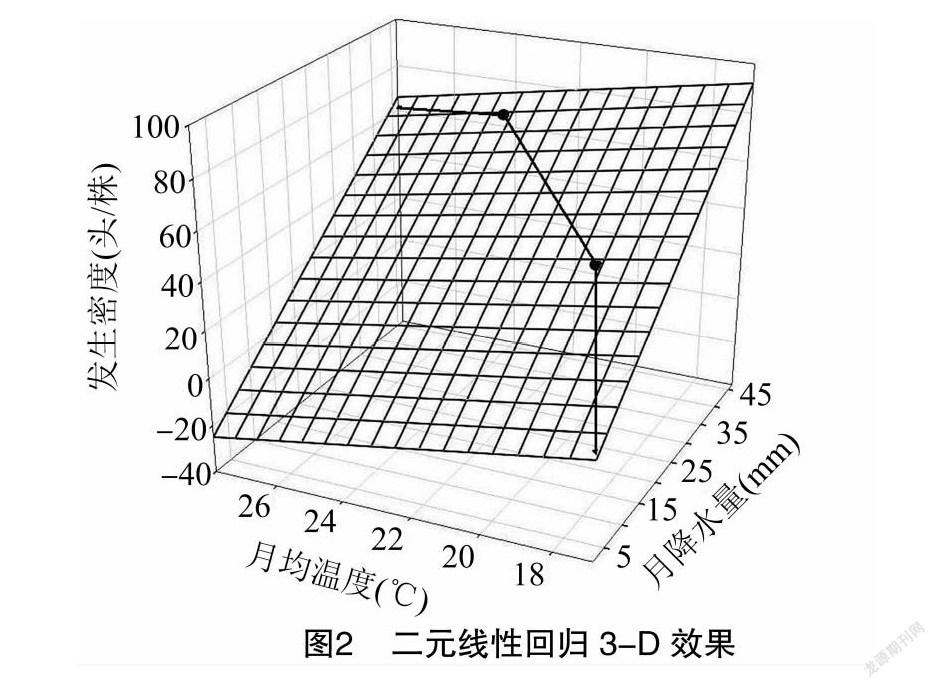
式中:y为发生密度,头/株;x 1为月平均温度,℃; x 2为月降水量,mm。SigmaPlot软件绘出的二元线性回归3-D效果图见图2。
由图2可知,5、6月数据点紧贴拟合方程平面上方而4、7月紧贴下方,SigmaPlot给出的拟合相关度为0.992,拟合效果很好。说明春芝麻娇驼跷蝽的发生量与温度及降水量在理想生存条件下呈典型的正比例关系。2020年由于倒春寒的影响4、5、6月等3个月试验田平均发生密度为0.08、12.39、38.87头/株,史庄平均枝梢调查结果为0.09、8.81、18.87头/株。2组数直线回归相关度 R 为0.986,未来完全可利用泡桐林旁的春芝麻娇驼跷蝽发生密度的线性变换值测报泡桐密度,其数据较捕虫网罩头法调查的数据稳定可靠。
由于气候湿热与天敌较多,系统田周围夏芝麻田的百株虫量调查点数据差异颇大,2020年9月夏芝麻试验田30个调查点百株虫量均值高达 4 932.07头。由于迁移新寄主后其种群将出现世代重叠,之后该虫有呈指数增长型暴发性发生的可能性。
2.2 聚集度指标计算与理论抽样数确定
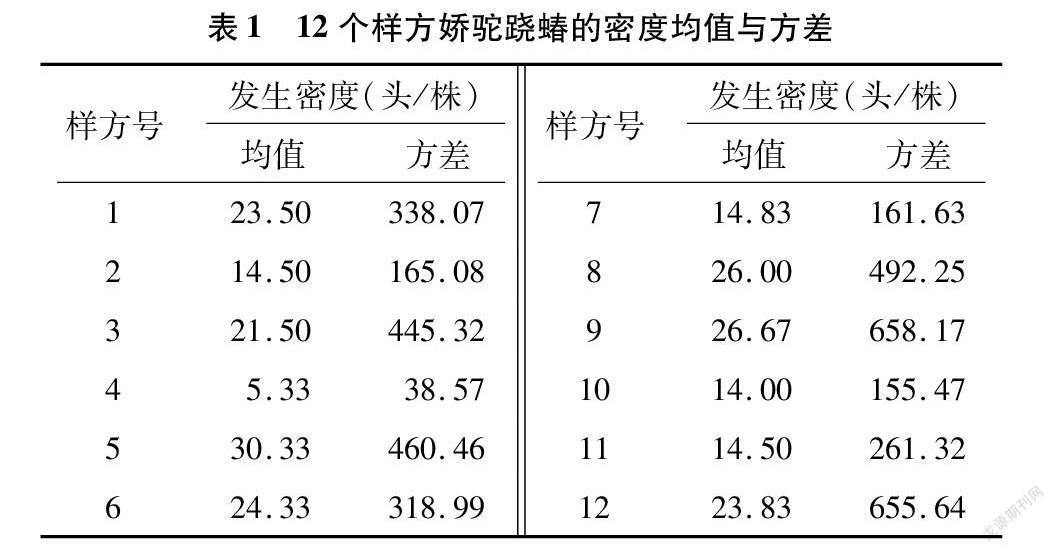
2021年6月上旬分布规律调查的12个调查样方娇驼跷蝽发生头数均值与方差如表1所示。
由表1可以看出,娇驼跷蝽在春芝麻植株上分布的聚集程度差别较大。将3块试验田的每株发生量均值数据输入DPS18.10进行3组数据的单因素方差分析,3地块平均每株发生量依次为16.21、23.87、19.75,置信度 P 0.05与 P 0.01等2水平 LSD 检验的结果均为差异不显著。所以3块地虽被大田隔开,但娇驼跷蝽发生的生态学特征是一致的。将表1 数据输入DPS分布型数据处理模块得到了每个调查样方各个聚集度指标的具体值,12个样方 M*指标、I指标、M*/M 指标、Kuno模型CA指标、扩散系数C、负二项分布 K 指数等各分布型参数的均值与变化区间依次为35.04±15.31、15.10±11.41、1.80±0.37、0.80±0.37、16.10±11.41、1.37±0.77。由于调查地块间苗时间较晚且没有打顶,导致田间芝麻植株丛生长状况差异性较大,所以各分布型指数数据差异性较大。12个样方CA指标全部大于0,因此娇驼跷蝽在芝麻大田呈现弱聚集分布型,一般密度大聚集度略大,平均扩散系数C为16左右[11-13]。
DPS还给出了IWAO 平均拥挤度-密度( M*-M)回归方程:M*=4.71+1.52M(r=0.92)。此方程的α与β 值依次为4.71与1.52,说明该虫为弱聚集分布型且個体之间是相互吸引的,这可能是昆虫信息激素的作用以及喜吸食芝麻幼嫩的叶与蒴果等原因造成的。将IWAO方程的 α与β 值(4.71与1.52)代入IWAO最适抽样数公式可得到序贯抽样中该害虫最适抽样数公式(2):
Q=4d25.71m+0.52。(2)
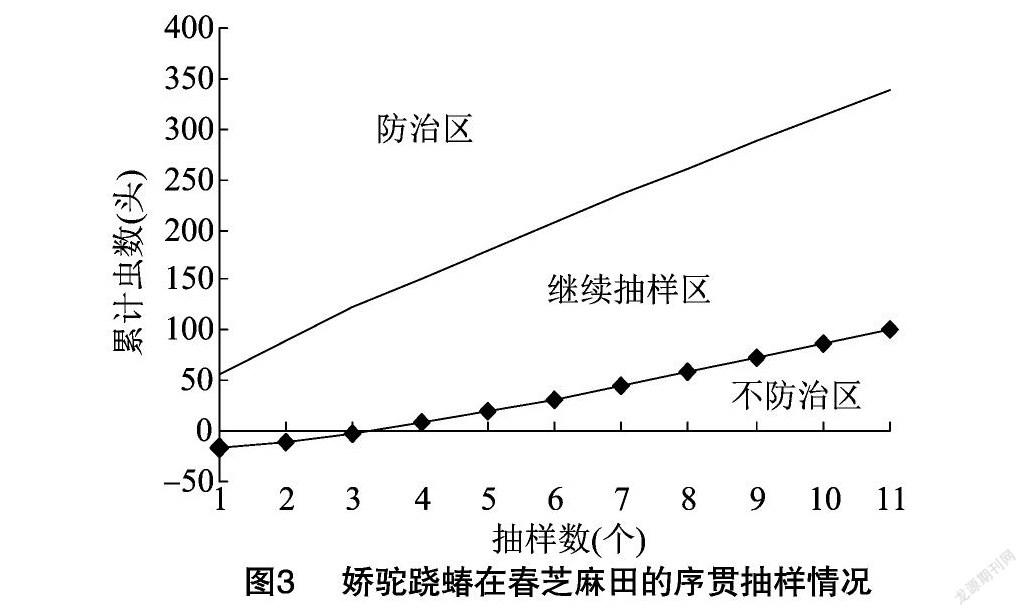
式中:d为允许误差;m为虫口密度;t为在一定置信度p下的t分布值,当p=95%时,t =1.96≈2。如果允许误差为0.3,害虫平均密度为20头/株,式(2)计算可得 Q 为35.8,所以本次调查设计合理。式(2)为这种春芝麻田春季新发害虫在我国中部地区的发生密度可靠性调查确定了一个标准。在河南省6月上旬发生密度20头/株以下时,每张叶片平均不到1头虫,将 α与β 值以及20头/株防治阈值代入序贯抽样累计虫数公式进行绘图,结果见图3。
一种农田害虫对不同地理条件下的不同寄主作物有着不同的分布格局特征和为害特点,图3为娇驼跷蝽的科学调查与有效防治提供了一个参考标准。
2.3 CA分析结果
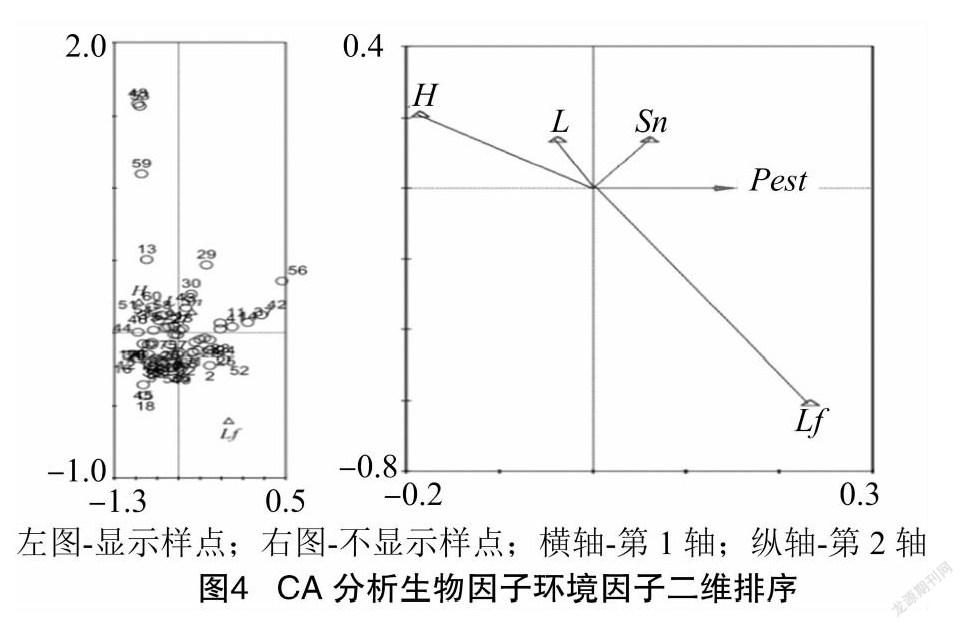
将2021年终花期调查的株高(height)、穗长(length)、蒴果数(num. of Sesamum indicum capsule)、叶片均长度(mean of foliage length)、所附着的娇驼跷蝽虫数(pest)5个变量依次命名为 H、L、Sn、Lf与Pest ,将60个调查点的经过无量纲化变换的4列自变量数据与害虫发生密度因变量数据输入CONOCO,进行1个生物因素4个环境因素的CA分析。CA分析结果显示物种累计关联度百分率第1轴到第4轴依次为4.2%、85.6%、 96.2%、100.0%,因此此害虫在植株上分布与生境条件密切。CONOCO绘图程序绘出CA分析的带调查点信息(图4左)与不带调查点信息(图4右)的生物因子与环境因子二维排序图。
由图4左图可知,60个调查点针对各个环境因素与生物因素的关联度差异较大,所以各样点对应散点分布分散,说明该次调查取的样点有代表性,散点越分散越好。图4右图显示去掉调查点散点后5个变量的相关关系。生态学排序图中,2个矢量之
间的相关度大小可以从一个矢量箭头在另一个矢量所在直线的垂直投影点距原点的距离直观看出[13]。所以娇驼跷蝽虫量与芝麻植株高度呈现典型的负相关关系,即高而粗壮的植株虫子少。芝麻茎杆各节的叶片大小与娇驼跷蝽分布呈现典型的正相关关系,即叶片较长便于虫体躲藏与吸食危害的植株虫子多。蒴果的数量与花穗的长度与因变量关联度不大但呈现一个呈正而一个呈负的关系,说明娇驼跷蝽喜在花穗短而蒴果多的、过早老熟一些的果穗上聚集。图4中 H与Lf 典型负相关的原因是这个季节青色蒴果开始快速生长,高的植株叶子并不茂密。
2.4 RF建模分析结果
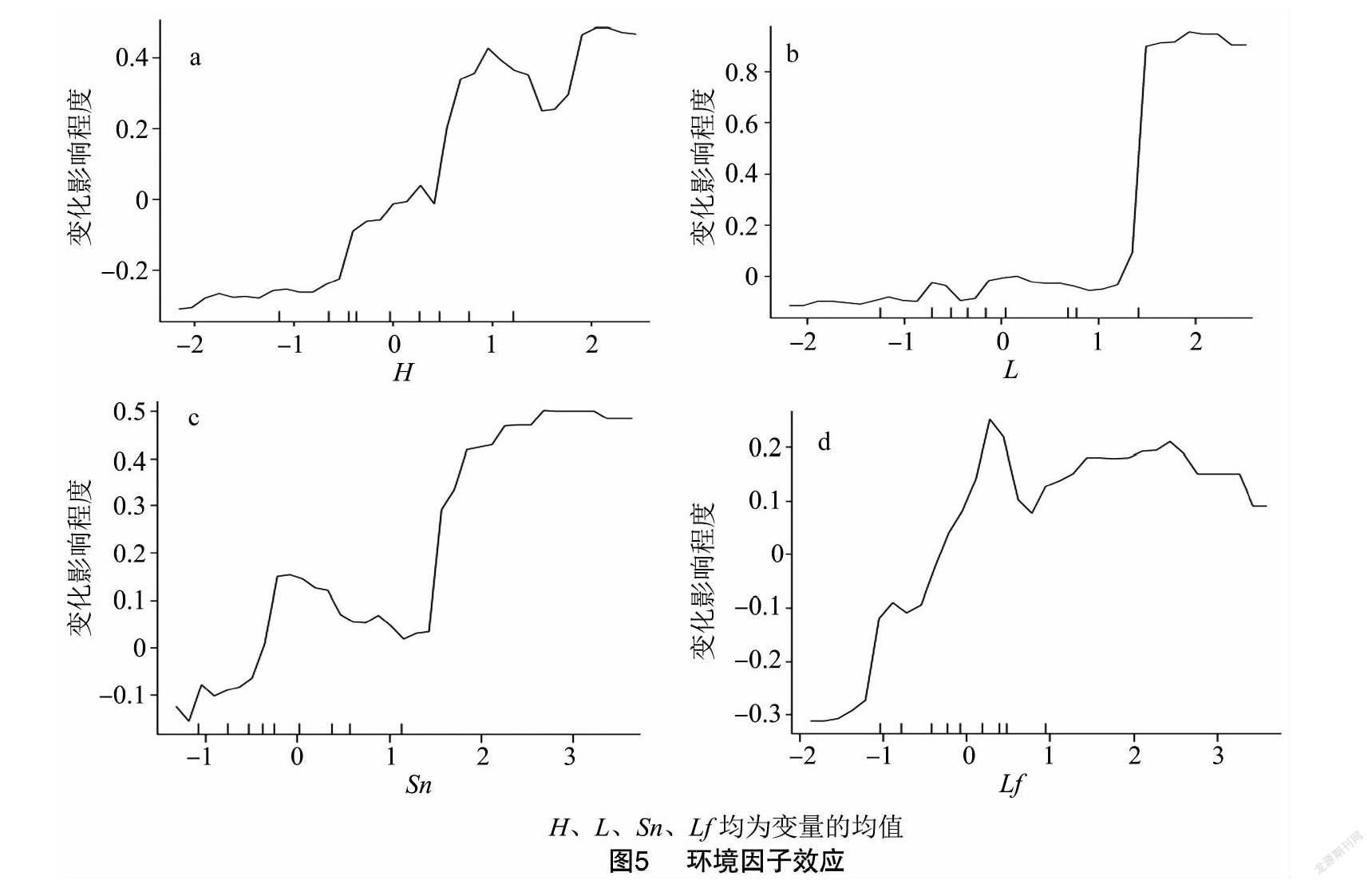
运行R软件RF建模的模型名称命令得模型总残差平方和仅为0.63,拟合效果很好。运行varImpPlot得到均方误差增量与结点纯度2种算法的4个环境因子对因变量影响效应的重要值与排序图,重要性顺序为 L>H>Sn>Lf ,可看出与CA分析结果一致。partialPlot命令所绘制的环境因子对娇驼跷蝽发生的各个效应见图5。
图5为4因子不同幅度的偏效应图,纵坐标为对因变量的影响值而横坐标为自变量标准化变换值,横坐标0点显示自变量均值对因变量的影响力。 H变量偏效应图显示该地块高于均值的植株发生量远低于低矮植株;L变量偏效应图显示穗长较均值略长的植株与略短于均值的发生量差别不大,但穗最长的一批健壮植株发生量变小;Lf 变量图说明叶越长虫子越多,但叶长高于均值一定程度虫子发生量差别不大。
2.5 加权列联表预测分析结果
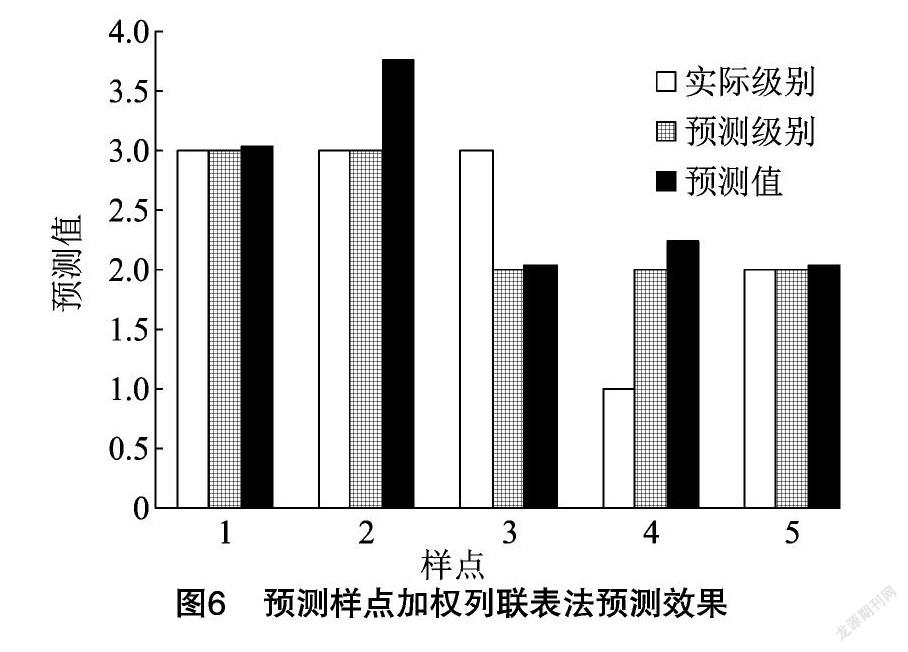
将55个样点的分级后数据进行加权列联表分析得到了 H、L、Sn、Lf 等4个环境因子变量的随机系数矩阵为0.53、0.47、0.37、0.34,其对应的卡方值依次为21.49、15.38、8.78、7.08。随机系数可以间接看出各个自变量对因变量的影响大小,因此可看出加权列联表分析结论和CA、RF分析相同,即前2个自变量对娇驼跷蝽发生量影响较大而后2个变量影响小[11]。DPS给出的55个训练样点预测级别,有35个完全正确,而且预测级别错误的20例预测值也全部接近正确的结果。5个预测样点预测值如图6所示。
图6显示四舍五入虽然仅3例预测级别正确,但5个预测值和实际级别有明显同步关系。对于娇驼跷蝽这样体积小迁移性强的外来害虫,寄主植物提供的食物与躲避条件、微气候条件与害虫分布密切相关,年度总体发生规模确定情况下单凭植物生长指标与小规模调查就可以大致预测其密度水平[15-16]。
3 讨论与结论
目前我国夏芝麻产量远远大于春芝麻,但一些小形地块春芝麻也较常见,大田地边的春芝麻便于捕食螨与东亚小花蝽( Orius sauteri )等多种微小天敌昆虫栖息生存,因而具害虫天敌植物支持系统的生物防治功效[17-18]。栽培春芝麻采收菜用芝麻叶也是农民的增收手段,以后中芝叶1号等叶用芝麻大规模推广将使得娇驼跷蝽在农业昆虫中的研究地位提高。根据江西、豫南、陕南、安徽等地以前的研究报告,该害虫在亚热带地区春夏季可达很高的种群增长速度,本研究表明豫北倒春寒情况下早春每月亦递增10倍以上[4-6]。研究表明,春芝麻娇驼跷蝽的密度与环境气象条件、寄主生长状况有对应关系,可利用春芝麻数据线性变换后测报周围林木害虫密度,克服乔木寄主对微小飞虫不易精确定量统计的缺点。
春芝麻此害虫若为害较重达30头/株以上,可用20%增效氯氰菊酯3 000倍液或1.8%阿维菌素乳油3 000倍液、3.5%锐丹乳油1 000倍液等药剂防治。4—5月此虫系杂草、枯枝落叶或泡桐等处迁移而来,因此切断迁移途径对防治效果至关重要,此外田间大草蛉[ Chrysopa pallens (Rambur)]与寄生蜂等天敌昆虫抑制其发生亦有重要作用。
本研究表明娇驼跷蝽具典型的r-生存策略特征,必须警惕其暴发性发生的可能性,对其测报与防治要考虑寄主生长指标、天敌与气候条件,此虫林业与农业2个方面数据可以实现相互测报。此外,此虫为害夏芝麻与秋芝麻的发生规律、防治标准、防治技术与其他生态学的研究有待进一步进行。
致谢:许蔓飞、陈静、陈静、李丹、陈小会等参与了部分试验与分析工作,浙江大学应用昆虫研究所唐启义老师提供DPS正版注册号,在此一并致谢。
参考文献:
[1]萧采瑜. 中国跷蝽科记述(半翅目:异翅亚目)[J]. 昆虫学报,1974,17(1):56-66.
[2]李祉宣,周 钰,陈赛赛,等. 江苏省文化树种认知情况调查及园林应用建议[J]. 江苏农业科学,2020,48(15):170-175.
[3]唐桂梅,姜卫兵,翁忙玲. 蔷薇科树种在南京园林绿化中的应用与评价[J]. 江苏农业科学,2013,41(7):182-185.
[4]孙志强,乔 杰,傅建敏,等. 泡桐林内同翅目、半翅目昆虫种类及其动态研究[J]. 林业科学研究,1999,12(5):510-514.
[5]吴学萱,陈绪山. 娇驼跷蝽的发生及其防治[J]. 昆虫知识,1991,28(6):344-345.
[6]张存立. 泡桐娇驼跷蝽生物学特性及防治研究[J]. 安徽农业科学,2007,35(6):1721-1731.
[7]孙志强. 泡桐丛枝病媒介昆虫传播植原体特点的研究[D]. 北京:北京林业大学,2000:20-22.
[8]韩俊梅,吕 伟,任果香,等. 200份芝麻种质资源农艺性状遗传多样性分析[J]. 江苏农业科学,2019,47(13):95-99.
[9]孙 路. 中国跷蝽科的分类(异翅亚目:长蝽总科)[D]. 北京:中国农业大学,1990:2-11.
[10]Aukema B,Reiger C. Catalogue of the Heteroptera of the Palaearctic Region. Vol. l. Enicocephalomarpha,Dipsocaromorgha,Nepomarpha,Gerromorpha and Leptopodomorpha[M]. Amsterdam:The Netherlands Entomological Society,1995:2-221.
[11]唐启义,冯明光. DPS数据处理系统:实验设计、统计分析及数据挖掘[M]. 北京:科学出版社,2007.
[12]黄胜先,李佳林,范 斌,等. 思州柚园黑蚱蝉若虫羽化空间分布型及羽化过程[J]. 江苏农业科学,2020,48(10):121-124.
[13]张金屯. 数量生态学[M]. 北京:科学出版社,2004:131-192.
[14]张 雷,王琳琳,张旭东,等. 随机森林算法基本思想及其在生态学中的应用:以云南松分布模拟为例[J]. 生态学报,2014,34(3):650-659.
[15]陳西亮,张佳华,艾天成. 基于支持向量机法提取江汉平原三湖农场棉蚜危害程度的空间分布[J]. 江苏农业科学,2016,44(9):157-162.
[16]殷 茵,龚卫良,陆 彦,等. 张家港市不同测报工具对甜菜夜蛾的监测比较[J]. 江苏农业科学,2019,47(24):70-74.
[17]陈学新,刘银泉,任顺祥,等. 害虫天敌的植物支持系统[J]. 应用昆虫学报,2014,51(1):1-12.
[18]Blaauw B R,Isaacs R. Larger wildflower plantings increase natural enemy density,diversity,and biological control of sentinel prey,without increasing herbivore density[J]. Ecological Entomology,2012,37(5):386-394.
