白马雪山自然保护区植物物种多样性海拔分布格局
2022-05-07聂佩潇张彩彩龙晓斌李延鹏肖文
聂佩潇, 张彩彩,2,3*, 龙晓斌, 李延鹏,2,3, 肖文,2,3
1. 大理大学东喜玛拉雅研究院,农学与生物科学学院,云南 大理 671003;
2. 中国三江并流区域生物多样性协同创新中心,云南 大理 671003;
3. 大理大学三江并流区域生物多样性保护与利用省创新团队,云南 大理 671003;
4. 白马雪山自然保护区维西管护分局,云南 迪庆 674600
山地植物群落生物多样性随海拔梯度的变化规律一直是生态学研究的热点[1]。海拔梯度作为生态学研究的天然实验室,综合了温度、水分和光照等多种环境因子,形成了显著的梯度效应[2]。沿海拔梯度的环境因子的剧烈变化导致物种分布呈现出一定的变化规律[3]。因此,海拔梯度成为生态学家们研究生物多样性格局的理想场所。国内外学者围绕物种多样性的海拔分布格局进行了广泛而深入的研究[4]。早期的研究发现,随着海拔的升高,物种多样性呈单调递减分布格局[5]。直到Rahbek (1995) 发表的综述,才让人们对物种多样性的海拔格局有了新认识,Rahbek在总结前人研究案例的基础上发现物种的丰富度在海拔梯度上呈先升高后降低的单峰分布格局,中海拔的物种丰富度最高,单峰格局的数量远远大于单调递减格局的数量[6]。目前有近一半的研究支持单峰格局,近25%的研究支持单调递减格局,并且在多种类群中得到了充分的验证[7]。由于低海拔受人为干扰比较严重,一些研究还发现物种多样性随着海拔的升高呈单调递增格局[6]。除了上述三种分布格局外,科学家们还发现,随着海拔的升高,物种多样性呈双峰、先下降后升高、无明显规律和基本保持不变等其他分布格局[8-10]。
研究发现不同生活型的植物种类沿海拔梯度的多样性分布格局存在差异[11]。例如,徐成冬对高黎贡山北段植物物种多样性的垂直分布格局研究表明木本植物物种丰富度随海拔升高呈递减趋势,草本植物随海拔升高呈先下降后上升趋势,海拔梯度上热量和光照的变化是影响木本植物和草本植物物种丰富度格局形成的主要原因[11];高辉等人的研究结果表明不同类别的环境因子是影响不同生活型植物物种丰富度的主要因素,乔木层随海拔变化呈单峰模式,灌木和草本呈双峰变化模式。乔木物种丰富度主要受干扰程度、坡向等的影响,灌木物种丰富度主要受海拔的影响,草本物种丰富度受海拔和湿度的影响[12]。
白马雪山自然保护区,隶属横断山脉东部云岭,是世界上高山植物区系最丰富的地区[13]、中国低纬度高海拔且生物资源保存完整的高山针叶林区[14],也是生物多样性保护的热点区域。本研究以白马雪山保护区响古箐片区海拔梯度上的森林植物群落为研究对象,开展森林群落植物多样性垂直格局研究,进一步探究白马雪山森林群落不同生活型植物在海拔梯度上的分布规律,为预测气候变化下物种多样性的变化规律提供数据支撑。
1 材料与方法
1.1 研究区概况
白马雪山国家级自然保护区(27°36′N,99°15′E)位于云南省西北部[15],横断山脉东部云岭的一部分,属于“三江(金沙江、澜沧江、怒江)并流”地区,也是青藏高原向云贵高原的过渡地带。区域内地形复杂,物种丰富,累计有维管植物52科,87属,145种。白马雪山国家级自然保护区属高原寒温带气候,具有“小气候复杂,气温年较差小,日较差大”等特点。年平均气温9.8℃[16],年均降水量1 200~1 500 mm[17]。该区土壤类型丰富主要有褐土、山地棕壤、山地暗棕壤、山地针叶土、亚高山草甸土和高山荒漠土[16]。参照《云南植被》[18]将该地区植被类型沿海拔梯度由低到高依次划分为云南松(Pinus yunnanensis)林、常绿阔叶林、针阔叶混交林、寒温性针叶林和高山草甸。
1.2 研究方法
1.2.1 样地选择与设置
本研究在白马雪山国家级自然保护区南端的响古箐片区,沿海拔2 300~3 500 m每隔200 m选择坡向和坡度较一致的区域,设置了一块面积为20 m×20 m样地,将每个20 m×20 m的样地划分为5个5 m×5 m的小样方,对样方内胸径≥5 cm的乔木进行测量,记录乔木的物种名、胸径、树高和生长状况。在20 m×20 m样方内的4个角和中心设置1个5 m×5 m样方进行灌木层调查,记录灌木种名、株丛数、平均高度和盖度。同样在5个灌木丛的边角设置1个1 m×1 m小样方进行草本层调查,记录草本植物种名、株丛数和盖度。另外记录各样地的经纬度、海拔、坡向、坡度、坡位、干扰方式、干扰程度和生境特点等基本信息。
1.2.2 数据处理与计算方法
(1) α多样性测定
α多样性反映了生境内的多样性。物种丰富度指数、Shannon-Wiener指数、Simpson指数和Pielou均匀度指数是测算α多样性的常用指数。
(a)物种丰富度(Richness):R=S

Pi种的个体数占群落中总个体数的比例。
(d) Pielou均匀度指数:E=H/LnS[12]。
H为实际观察的物种多样性指数,LnS 为最大的物种多样性指数(S为群落中的总物种数)。
(2) β多样性测定
β多样性指沿环境梯度不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率也被称为生境间的多样性[19]。
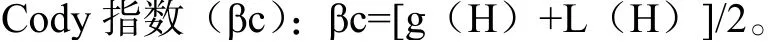
g(H)是沿生境梯度H增加的物种数目;L(H)是沿生境梯度H失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目。
2 结果与分析
2.1 物种组成
在白马雪山响古箐2 300~3 500 m海拔跨度内的7块样地中共记录到145种植物,隶属于52科87个属(见表1)。其中乔木33种,隶属于15科26属,每个海拔样方内选择数量最多的物种作为优势物种,共获得乔木样方的3个优势物种:云南松 (Pinus yunnanensis)、云南铁杉 (Tsuga dumosa)和团花杜鹃 (Rhododendron anthosphaerum) (见表1);灌木65种,隶属于27科37属,优势物种分别是地檀香(Gaultheria forrestii)、光叶栒子 (Cotoneaster glabratus)和野扇花 (Sarcococca ruscifolia) (见表1);草本47种,隶属于27科40属,优势种为高山早熟禾 (Poa alpina)、荩草(Arthraxon hispidus)和长穗兔儿风 (Ainsliaeahenryi) (见表1)。

表1 不同海拔内各生活型的优势物种Tab. 1 Dominant species of different life froms at different elevations
2.2 α多样性随海拔梯度的变化
对不同海拔梯度乔木层、灌木层和草本层群落的丰富度指数、Shannon-Wiener指数、Simpson优势度指数和Pielou均匀度指数的分析,结果表明:随着海拔梯度的上升,乔木层物种丰富度指数和Shannon-Wiener指数呈先上升后下降再上升的“双峰”分布格局。草本层丰富度指数和Shannon-Wiener指数呈递减趋势。灌木层丰富度指数和Shannon-Wiener指数呈先下降后上升趋势。乔木层、灌木层和草本层的Simpson指数和Pielou指数随海拔的上升均呈先下降后上升的“V”字型分布格局(见图1)。

图1 海拔梯度上α多样性指数的变化格局Fig. 1 Altitude pattern of α diversity index in Baima Snow Mountain
2.3 β多样性随海拔梯度的变化
β多样性能够反映样地间或者海拔梯度间植物群落差异,通过对乔木层、灌木层和草本层物种β多样性沿海拔分布模式进行分析,结果表明:乔木层、灌木层和草本层的β多样性Cody指数在海拔梯度上均呈先增加后减小的“单峰”模式,这说明随着海拔梯度的增加,相邻海拔间植物群落物种更替速率先升高后降低,群落内生境异质性先增大后减小(见图2)。
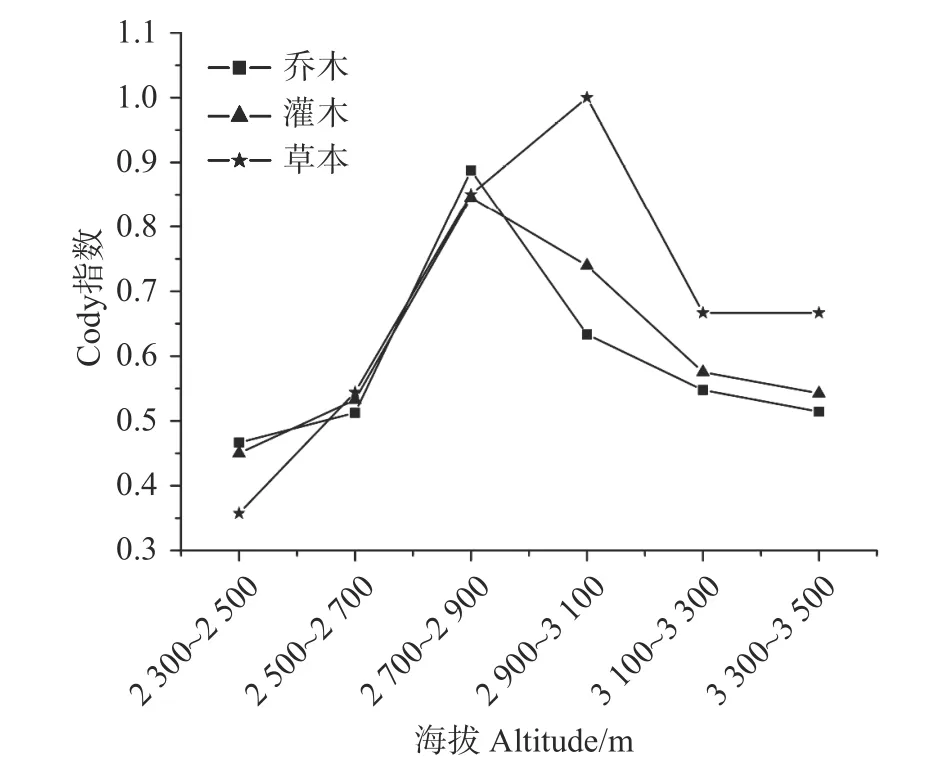
图2 海拔梯度上β多样性指数的变化格局Fig. 2 Altitude pattern of β diversity index in Baima Snow Mountain
3 结论与讨论
3.1 α多样性在海梯度上的分布格局
物种多样性随海拔梯度的变化在群落物种多样性空间分布格局中备受生态学界关注[1]。本文研究结果表明,白马雪山响古箐地区植物群落物种多样性的海拔分布规律在不同生活型植物种类上存在差异。冯建孟等人[20]的结果表明:由于低海拔受人为干扰造就了木本层植物物种多样性随海拔呈“单峰”格局。本研究中乔木层丰富度指数和Shannon-Wiener随海拔升高呈“双峰”格局,丰富度指数在海拔3 300 m出现最低峰值,团花杜鹃(Rhododendronanthosphaerum)为此海拔范围的优势种群。在海拔3 100m出现最高峰值,此海拔区域为针阔混交林,铁杉林混交团花杜鹃、多脉茵芋(Skimmia multinervia)等阔叶树种,因此物种多样性较高。Simpson指数和Pielou指数随海拔升高呈现“V”型格局说明选用不同的多样性指数随海拔梯度的分布格局存在差异。
灌木层的四个α多样性指数和草本层的Simpson指数和Pielou指数均随海拔先减小再增大,呈“V”字型格局,在海拔2 900 m处出现一个低峰值。林下层的灌木和草本容易受到人为干扰和林冠郁闭度的影响[21],2 900 m处的乔木层以云南铁杉(Tsuga dumosa)为主,林冠郁闭度高,故林下光资源可获得性较低[22],而且林下丰富的枯枝落叶导致林下灌木和草本减少,此外中海拔区域又容易形成以野扇花(Sarcococca ruscifolia)为主的单优势灌木层,故中海拔灌木层多样性最低。草本层植物群落物种多样性以“单峰”格局较为普遍[23,24],而本研究结果表明草本层物种丰富度和Shannon-Wiener指数呈现出递减格局。低海拔地区受人为或动物活动(放牧)的影响,可能导致了种子的传播概率增加,最终导致低海拔范围草本层物种多样性相对较高[25],而高海拔地区受热量限制不利于林下草本植物的生长[26]。
3.2 β多样性在海拔梯度上的分布格局
β多样性能够反映出不同生境间植物群落间的梯度变化或环境梯度中的物种更替速率。一些研究表明植物物种β多样性随海拔的升高呈递减趋势,例如赵淑清等[27]人在长白山的植物多样性研究结果表明乔木、灌木和草本的β多样性均随海拔上升呈递减趋势,原因是低海拔生态位窄,较小的生境能引起较高的物种更替,因此呈递减趋势。牛常青[28]在乌金山植物多样性也发现低海拔生境异质性大而导致了β多样性随海拔呈递减趋势。本研究发现随着海拔的升高,乔木层、灌木层和草本层Cody指数呈先升高后降低的“单峰”模式,中海拔区域为次生林到原生林的过渡区故中海拔多样性较高。而且草本植物很大程度上易受光照、群落盖度、微地形的影响因此随海拔变化比乔木和灌木更明显[29]。
综上,本研究发现随着海拔的升高,α多样性有呈先增加后减小再增加的“双峰”模式、递减模式和先减小后增加的“V”型模式,β多样性呈先升高后降低的“单峰”模式,且在不同生活型植物中存在差异。因此白马雪山自然保护区植物群落在实现可持续发展时要根据植物群落在海拔梯度上的变化规律,科学合理地制订保护计划和管理措施才能持久地维持保护区生态系统的稳定。
