玉米DIBOA葡萄糖苷转移酶基因bx8的克隆及其生物信息学分析
2022-05-05黄伟鹏徐碧莹张敏艳吴委林郑大浩
黄伟鹏, 徐碧莹, 张敏艳, 吴委林, 郑大浩
(延边大学 农学院,吉林 延吉 133002)
玉米是全世界也是我国种植范围最广、总产量最高的作物[1],含有丰富的玉米黄质、维生素E及谷胱甘肽,具有补脑、降糖、降压降脂、护眼、抗衰老等功效。自然界中,危害玉米的害虫种类超过90种,蚜虫是玉米虫害之一,对玉米产量和质量有很大影响[5]。
目前,玉米蚜的防治主要依赖于化学防治,但是化学药品易引起环境污染,所以,选育推广抗蚜品种是减轻或避免蚜害的有效途径。而广泛存在于禾本科植物中的次生代谢物苯并噁嗪类化合物可作用于玉米抗蚜中[6-7]。苯并噁嗪类化合物能够帮助玉米从土壤中摄取铁,将有益微生物吸引到与植物根部接触的土壤区域,也能对防御外界胁迫起到一定的促进作用,是玉米适应外环境的重要组件,其中,研究较多的是“的布”和“丁布”。在玉米和麦类作物中,“丁布”含量越高,对蚜虫抗性越好[16]。
植物中,已发现的苯并噁嗪类化合物有20多种,苯并噁嗪合成途径涉及的基因至少有12个, 虽然DIBOA(2,4-二羟基-1,4-苯并噁嗪-3-酮)被认为是玉米抗虫性密切相关的代谢物质[21],但不同的玉米对抗虫性存在很大差异其原因是由于DIBOA的含量高低所引起,还是由于DIBOA葡萄糖苷酶的变异所导致的下游苯并噁嗪类物质的合成受阻而引起的还不清楚。为此,该研究依据玉米标准基因组上所注释的DIBOA葡萄糖苷转移酶基因bx8的参考序列,从高抗玉米Mo17苗期叶片中克隆bx8基因,为bx8基因的表达和功能验证奠定基础。
1 材料与方法
1.1 材料
该试验用于克隆目的基因bx8的试验材料为高抗蚜玉米自交系Mo17苗期的叶片。
1.2 方法
1.2.1 玉米DIBOA葡萄糖苷转移酶基因的查询与引物设计
首先以玉米标准基因组数据库(http://www.maizegdb.org)上下载玉米DIBOA葡萄糖苷转移酶基因的基因序列GRMZM2G085054_P01为基础,利用Primer Premier 6.0和AutoDimerV 1.0软件设计引物,再引入通过DNAStar软件检测出的限制性内切酶位点,即EcoRⅠ(5′-GAATTC)和XhoⅠ(5′-CTCGAG)限制性内切酶位点,正向引物为BX8EC625F1:GAATTCCCAGACAGAGAGGACAGG(下划线部分为EcoRⅠ限制性酶切位点),反向引物为BX8XH2277R2:CTCGAGATGATGAATGATGAGAAGC(下划线部分为XhoⅠ限制性酶切位点)。在玉米标准基因组中引物区间内目标DNA序列长度为1 534 bp。
1.2.2 目标DNA片段的PCR扩增
以玉米自交系Mo17为基因组DNA模板进行PCR扩增,选用Promega的PCR试剂盒(Go Taq Green Mix),PCR反应体系参考试剂盒说明书,采用梯度PCR的方法摸索扩增所需最佳温度,PCR反应基本条件在退火温度48~62 ℃设置温度梯度,最终PCR反应的体系为模板DNA 2.5 μL;Forward primer 1.25 μL;Reverse primer 1.25 μL;Go Taq Green Mix 12.5 μL;ddH2O 7.5 μL。
1.2.3 PCR产物的回收连接及转化
根据梯度PCR摸索的最佳温度进行目标DNA片段扩增,PCR反应结束后利用1%的琼脂糖凝胶电泳将大小不同的条带进行分离,利用凝胶成像仪,在紫外灯照射下快速切取目的条带,放入事先准备好的TB管中,利用E.Z.N.A.GelExteaction Kit胶回收试剂盒回收DNA目的片段;回收方法按照试剂盒说明书进行操作,并对回收产物进行电泳检测,检测出目的条带后进行连接,连接载体为pMD19-T,连接条件为4 ℃过夜,连接结束后,将含有目标DNA片段的重组质粒导入到感受态大肠杆菌DH5α中,在含有氨苄青霉素、IPTG、X-Gal的培养基上进行培养,利用蓝白斑筛选,选择阳性菌,即重组质粒(白色)。
1.2.4 质粒提取与目标DNA序列酶切及PCR验证
挑取白色阳性单菌落进行液体培养,8~10 h后吸取少量菌液与灭菌后的去离子水按1∶20进行稀释,稀释后充分混匀进行划线,37 ℃恒温箱中过夜培养。准确无误后利用Omega公司的质粒提取试剂盒(E.Z.N.A.PlasmidMiniprep Kit)从大肠杆菌中提取和纯化质粒。经琼脂糖凝胶电泳检测正确后,按限制性内切酶说明书对所提质粒进行双酶切及其鉴定。酶切验证正确后,将相应的菌液进行扩大培养,保存菌种后,将菌液送到上海生工公司测序。
1.3 目标序列的分析
1.3.1 目标序列的同源性分析
测序后得到的DNA序列,通过DNAStar软件和GeneDoc软件进行同源性比对和序列分析,在NCBI下载禾本科bx8蛋白序列,用软件MEGA7进行系统发育树构建。
1.3.2 目标基因的生物信息学分析
目标DNA序列编码的蛋白质序列分析,利用NCBI在线工具(http://www.ncbi.nlm.nih.gov/)对蛋白质序列进行同源序列比对;利用在线工具(http://www.expasy.org/tools/protparam.html)(http://ca.expasy.org/tools/protscale.html)对蛋白质序列进行理化性质及亲疏水性等一级结构分析;利用在线工具(http://www.cbs.dtu.dk/services/TMHMM-2.0/)对蛋白质跨膜区分析;利用SOPMA预测蛋白质二级结构;利用SWISS-MODEL(https://swissmodel.expasy.org/interactive)预测蛋白质三级结构。利用Net Phos3.1预测磷酸化位点。
2 结果与分析
2.1 引物的PCR最佳退火温度摸索
引物BX8EC625F1和BX8XH2277R2分离的目标DNA序列区间长度1 534 bp。为摸索该引物从玉米自交系Mo17中分离目标片段的PCR温度条件,在正向引物和反向引物的适宜范围内设置温度梯度,PCR仪自动设置8个温度梯度为:45.0、46.4、48.8、52.6、57.1、60.9、63.4和65.0 ℃,结果见图1。以Mo17基因组的DNA为模板进行PCR,当温度低于52.6 ℃时,条带不清晰,目标条带效果不好;温度在60.9 ℃时,目标条带清晰,效果较好且未出现非特异性条带。

注:引物扩增的目标DNA片段长度为1 380 bp。
2.2 目标序列的PCR产物回收与克隆
以DNA为模板的PCR扩增产物、PCR扩增产物回收、以重组质粒为模板的PCR扩增产物、重组质粒及重组质粒的限制性内切酶所得片段进行比较(图2)。从图2可知,目标片段均处于同一高度,条带清晰、明显,且无杂带。说明成功克隆了目标片段并插入到pmD-19T载体中,可进行测序分析。

注:第1泳道为2 000 bp DNA Marker;第2泳道为以DNA为模板的PCR产物;第3泳道为PCR回收产物;第4泳道为以重组质粒为模板的PCR产物;第5泳道为插入DNA片段的重组质粒;第6泳道为重组质粒的酶切产物;第7泳道为15 000 bp DNA Marker。
2.3 目标序列分析
2.3.1 目标DNA序列及其目的基因编码的蛋白质序列分析
从自交系Mo17中分离的DNA序列长度为1 534 bp,与玉米标准基因组(Ref_B73)参考序列GRMZM2G085054的目标DNA序列长度相同,但有3处碱基发生替换突变,并且该DNA序列具有完整的开放阅读框(ORF)(图3)。该目标DNA片段所编码的蛋白质序列(bx8Zm5054Mo17P)由459个氨基酸组成,与玉米标准基因组翻译的蛋白质GRMZM2G085054_P01(简称bx8Ref_Zm5054P)相比,存在3处差异,即自N末端起第177个氨基酸由脯氨酸(P)变为亮氨酸(L),第281个氨基酸由甘氨酸(G)变为精氨酸(R),第301个氨基酸由丙氨酸(A)变为苏氨酸(T)(图4)。

图3 来自玉米自交系Mo17的目标DNA序列与标准基因组参考序列的一致性比较
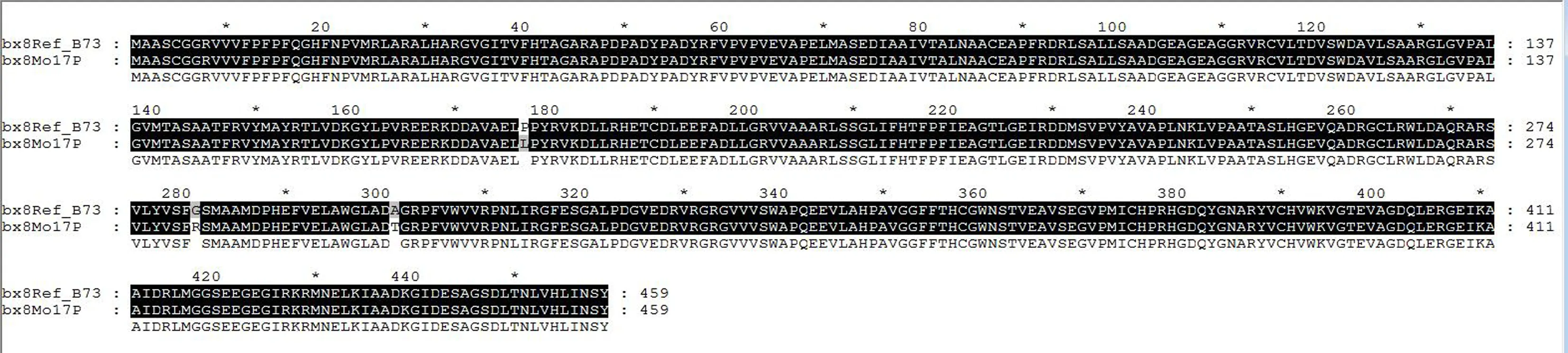
图4 玉米自交系Mo17的目的基因编码的蛋白质与标准基因组参考蛋白的氨基酸序列比较
2.3.2 目标序列的生物信息学分析
2.3.2.1 bx8蛋白的功能预测
将bx8基因编码的氨基酸序列导入到NCBI(Conserved Domain Database)数据库中进行相关蛋白功能结构域预测。结果表明,该蛋白有1个保守结构域(图5),该结构域位于第6~459位氨基酸之间,属于Glycosyltransferase_GTB-type超级家族。

图5 参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P保守结构域及其功能
2.3.2.2 蛋白质的理化性质分析
目的蛋白bx8Zm5054Mo17P和参考蛋白bx8Ref_Zm5054P的总元素数量(表1)分别为6 942和6 917,相对分子量分别为49 601.59和49 456.39。二者在C、H、N、O、S这5种元素构成中,除S元素在参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P中的数量均为19外,其他4种元素含量分别为2 203、3 451、627和642;参考蛋白bx8Ref_Zm5054P序列中分别为2 197、3 436、624和641。并且参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P的亲疏水性分别为0.042和0.045,虽然二者的负电荷残基总数均为59,但正电荷残基总数相差1个碱基,前者为45个,后者为46个。目的蛋白不稳定系数为35.24(<40),说明该蛋白稳定。

表1 参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P的基本理化性质
利用在线工具(http://www.detaibio.com/tools/signal-peptide.html)和在线软件TMHMM Server v.2.0对目的蛋白和参考蛋白的信号肽(图6)、亲疏水性(图7)和跨膜结构进行分析比对(图8)。结果显示,参考蛋白和目的蛋白均无信号肽,二者亲疏水结构极其相似;可以看出该蛋白不属于转运蛋白和分泌蛋白。且参考蛋白和目的蛋白均为无跨膜区,所以该蛋白是无信号肽、无跨膜区的膜内蛋白。

图6 参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P的信号肽

注:峰值<0表示亲水性,峰值>0表示疏水性。

注:蓝色实线表示伸入到细胞内的部分,粉色实线表示伸入到细胞膜外的部分。
2.3.2.3 二级结构和三级结构的预测
通过使用SOPMA预测目的蛋白的二级结构表明(表2),目的蛋白和参考蛋白均由α螺旋、β转角、无规则卷曲与延伸链构成,其中,α螺旋与无规则卷曲占氨基酸总数的比例较大,在参考蛋白中分别占41.18%与39%,在目的蛋白中分别占41.18%与37.69%,而β转角仅占了6.32%与6.97%。图9更为直观地表明参考蛋白和目的蛋白的二级结构中存在的差异。
利用SWISS-MODEL在线工具预测参考蛋白和目的蛋白的三级结构,得到三维结构图(图10)。由于目标DNA片段编码的蛋白质氨基酸序列与标准基因组上所推断的蛋白质氨基酸序列相比,存在3个氨基酸的差异,导致参考蛋白和目的蛋白的三级结构发生变化。

表2 参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P的二级结构
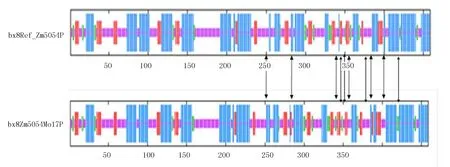
注:蓝色为α-螺旋;玫红色为无规则卷曲;红色为延伸链;绿色为β转角。

图10 参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P的三级结构
2.3.2.4 目标基因编码蛋白bx8Zm5054M017P与其他植物的同源性
来自Mo17的目的蛋白bx8Zm5054Mo17P在NCBI上BLAST结果见表3。bx8Zm5054Mo17P与玉米标准基因组推测的登录号为NP_001144409.2的DIBOA-葡萄糖苷转移酶BX8的氨基酸序列一致性达到99.35%,比对的序列覆盖率也是100%;与另一个登录号为AQK48639.1的蛋白质氨基酸序列一致性也达到99.35%,比对的覆盖率也是100%;与登录号为BAJ07092.1、XP_020168919.1、BAJ07089.1、AEB61490.1的比对序列覆盖率均为100%,但比对序列一致性相对较小,低于70%。目的蛋白bx8Zm5054Mo17P与其他植物的蛋白质同源性对比见图11。

表3 目标蛋白在NCBI在线BLAST同源性比对

图11 目的蛋白bx8Zm5054Mo17P与其他植物的蛋白质同源性对比
2.3.3 磷酸化位点分析
利用在线工具Net Phos3.1预测磷酸化位点(图12),目的蛋白和参考蛋白中磷酸化位点有25个。其中,Ser有16个,Thr有4个,Tyr有5个。Ser磷酸化位点为Ser73、Ser96、Ser100、Ser209、Ser210、Ser233、Ser274、Ser279、Ser282、Ser319、Ser338、Ser362、Ser368、Ser420、Ser444、Ser458;Thr磷酸化位点为Thr146、Thr189、216、224;Tyr磷酸化位点为Tyr161、Tyr237、Tyr277、Tyr383、Tyr388。
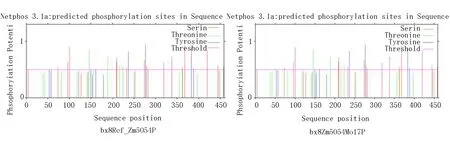
图12 参考蛋白bx8Ref_Zm5054P和目的蛋白bx8Zm5054Mo17P磷酸化位点
2.3.4bx8蛋白同源性分析及进化树
在NCBI在线工具下载与bx8高度相似的序列,并利用MEGA7构建玉米(Mo17)、普通小麦(Triticum aestivum)、节节麦(Aegilops tauschii subsp. Strangulata)、黑麦(Secale cereale)、野生二粒小麦(Triticum dicoccoides)、狗尾草(Setaria viridis)、柳枝稷(Panicum virgatum)、黄粟(Setaria italica)、高粱(Sorghum bicolor)、粳稻(Oryza sativa Japonica Group)、短花稻(Oryza brachyantha)、糙伏毛燕麦(Avena strigosa)、大麦(Hordeum vulgare subsp. Vulgare)、二穗短柄草(Brachypodium distachyon)的进化树(图13)。结果表明,玉米(Mo17)与其他禾本科植物进化树分为4支,玉米、普通小麦、节节麦和野生二黎小麦为1支;柳枝稷、狗尾草和黄粟为第2支;高粱、粳稻和糙伏毛燕麦为第3支;二穗短柄草、短花稻和大麦为第4支。玉米与普通小麦、节节麦和野生二黎小麦的亲缘关系都比较近。
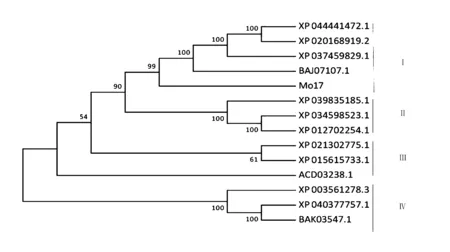
图13 bx8与其他植物氨基酸序列进化树
3 讨论与结论
苯并噁嗪代谢,从吲哚-3-甘油磷酸起始,历经吲哚、吲哚-2-酮、3-羟基吲哚-2-酮合成出2-羟基-苯并噁嗪-3-酮(HBOA);再由HBOA合成出DIBOA。DIBOA在葡萄糖苷转移酶的催化下合成出DIBOA-葡萄糖苷后,才能进入下游合成途径,最终合成出对害虫具有毒杀作用含甲氧基的各种不同的苯并噁嗪类化合物。所以,DIBOA葡萄糖苷转移酶是苯并噁嗪代谢途径中的关键酶之一。该研究从玉米的自交系Mo17中克隆到含有目的基因的目标DNA片段,长度为1 534 bp;在开放阅读框(ORF)有3处碱基发生替换突变。从而造成来自Mo17编码蛋白的氨基酸序列存在3个氨基酸变异,从而导致蛋白质三级结构的差异变化。虽然来自Mo17的bx8Zm5054Mo17P蛋白质分子量和总元素数量未发生变化,但其氨基酸序列中存在的3个氨基酸变异,造成了蛋白质构成元素C、H、N和S组成以及蛋白质多肽链的正负电荷量以及亲疏水特性等理化特性的略微变化。并且与其他禾本科植物的同源性分析有较高的一致性,说明该蛋白的功能较为保守。属于Glycosyltransferase_GTB-type 超级家族。
bx8基因存在于禾本科植物的多种作物中,包括玉米、小麦、水稻等[23],当植物遭受外界侵害,bx8基因主要以DIBOA为底物将DIBOA葡萄糖苷化形成DIBOA-Glc而后进入下游的代谢途径中,从而调控苯并噁嗪类防御化合物的合成;该研究中克隆的bx8基因所编码的蛋白质,存在3处氨基酸变异导致的蛋白质三级结构有两处发生变化是否影响玉米抗虫性的强弱还有待于进一步深入研究。
