云杉球果锈病研究进展
2022-04-29霍怀成杨帆张睿珂徐红霞曹秀文
霍怀成,杨帆,张睿珂,徐红霞,曹秀文
(1.甘肃省白龙江林业科学研究所 甘肃 兰州 730070;2.甘肃白龙江森林生态系统国家定位观测研究站甘肃 武都 746000)
云杉属Picea植物广泛分布在北半球高纬度寒温带、中低纬度和亚高山等区域,是针叶林主要的建群种[1],在亚高山地区天然林保护和林业生态建设中具有不可替代的作用。云杉球果锈病是云杉的主要病害之一,在北半球普遍发生[2]。云杉球果锈病不仅影响种子的质量和产量,还降低了种子活力、发芽率和出苗率[3-5],严重影响云杉种子的生产和云杉林的自然更新。
在查阅、整理国内外球果锈病相关文献资料和初步研究的基础上,从不同球果锈病的危害、症状、病原、发病规律、寄主范围及其防治策略方面对球果锈病进行了综述,以更好的了解球果锈病的种类、病原的生活史及寄主抗病性,合理制定防治策略,为云杉天然林和人工林的更新和可持续发展提供依据。
1 病原菌及其分布
侵染云杉球果,引起球果锈病的病原菌主要有杉李盖痂锈菌Thekopsora areolata、鹿蹄草金锈菌Chrysomyxa pyrolae和独丽花金锈菌C.monensis。杉李盖痂锈菌广泛分布于欧洲和亚洲北部,中美洲加勒比地区也有报道,美国将其作为限定性有害生物列入有害生物名录[6]。鹿蹄草金锈菌广泛分布于北温带偏北地区,在我国的分布区尚不清楚,可能仅限于东北地区[7]。独丽花金锈菌仅分布于北美洲海岸地区。另外,变形金锈菌C. deformans、伏鲁宁金锈菌C. woroninii和杜香金锈菌C. ledi也能侵染球果[8-10]。变形金锈菌只分布在亚洲,主要侵染云杉叶芽,在西喜马拉雅地区和我国新疆发现变形金锈菌也侵染球果;伏鲁宁金锈菌广泛分布于北半球亚高山林缘地区,主要侵染叶芽,也能侵染球果和茎干;杜香金锈菌广泛分布于欧亚大陆,侵染多种云杉针叶,2010年在欧洲云杉球果种鳞上发现金锈菌锈孢子,现已在多种云杉球果上发现该菌种。几种锈菌的分布如表1。
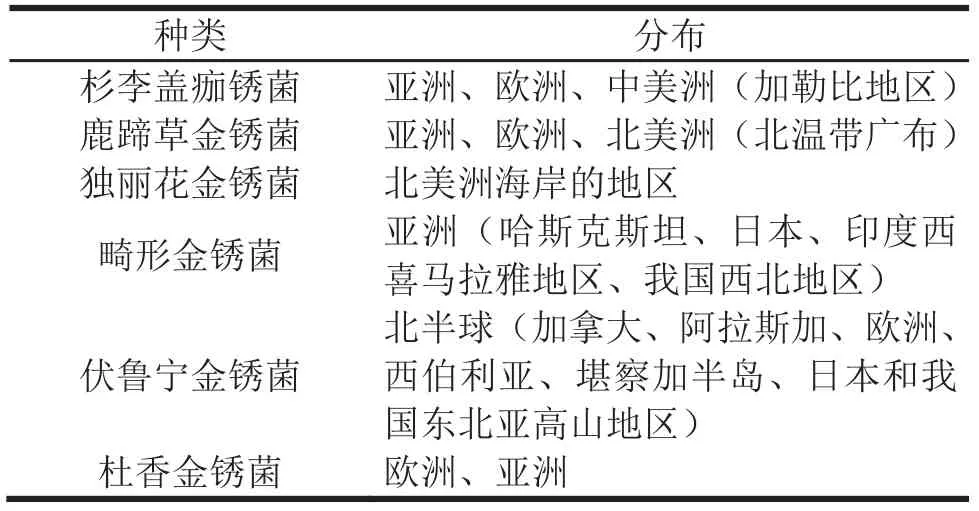
表1 云杉球果锈病病原菌种类及其分布
2 病害分述
2.1 稠李云杉球果锈病
2.1.1 危害
稠李云杉球果锈病对树势的影响很小,主要危害球果,影响种子的质量和产量[11]。2006、2009和2015年,芬兰的欧洲云杉P. abies上球果锈病大发生[11-13];2006年,瑞典北部70%的球果感染此病,2011年和2012年瑞典南部和北部再次发生[14],严重影响了云杉采种园的种子生产;我国小兴安岭和长白山林区,每年发病株率约为5%,病株被害果达20%[14];川西木城沟以粗枝云杉P. asperata为主的河滩林,粗枝云杉球果锈病感病率在64.5%,紫果云杉P. purpurea为30.7%[15]。云杉球果锈病还降低种子活力,Kaitera等发现感病球果的种子发芽率只有4%,而健康球果的种子发芽率约为37%,锈菌使种子的发芽率降低了10倍左右[7]。
2.1.2 症状
稠李云杉球果锈病主要侵染球果,有时也侵染种子和幼苗的茎干[17]。侵染球果,初期在球果种鳞背面出现黄色粘液,随后产生黑色点粒状物,即病菌的性孢子器。然后种鳞表皮下出现黄白色突起,后期突破表皮形成球状锈孢子器;锈孢子器颜色逐渐加深,最后变为红棕色,感病球果种鳞提早开裂。在川西地区,一般粗枝云杉球果10月成熟,感病球果8月下旬种鳞开裂[16]。第二年春季,锈孢子器破裂,散出黄色粉末状物,即病菌的锈孢子。稠李云杉球果锈侵染幼苗,主要危害嫩梢,被害嫩梢通常弯曲,呈“S”型,树皮变为深褐色,略肿胀,有时会流脂,幼苗茎干弯曲部分以上嫩梢枯死[17]。
2.1.3 病原菌及其生活史
病原为杉李盖痂锈菌隶属锈菌目,盖痂锈菌属[18]。
盖痂锈菌与膨痂锈菌属Pucciniastrum非常相似,主要是根据冬孢子堆的着生位置来区分,盖痂锈菌属的冬孢子堆位于寄主表皮细胞中,而膨痂锈菌属的冬孢子堆位于寄主的表皮细胞下[18]。杨婷[19]认为冬孢子堆着生位置不能作区分两属的分类依据,将盖痂锈菌属和膨痂锈菌属合并为广义的膨痂锈菌属,盖痂锈菌属是否与彭痂锈菌属合并为广义彭痂锈菌还需进一步研究和探讨。Cao等[20]和肖玉贵[21-22]描述了病菌性孢子和锈孢子,并研究了生物学特征,杨婷[19]描述了夏孢子特征。
生活史及侵染循环:杉李彭痂锈菌完成生活史需要2年,有5种不同的孢子类型[8],分别寄生在云杉和李属Prunus 的稠李P. padus[11]或微毛野樱桃P. clarorolira和多毛野樱桃P.polytrichakochne[23-24]上。因为在被侵染的球果种鳞上没有发现性孢子器,同时发现在远离转主寄主李属植物的区域病害也有发生,推测杉李盖痂锈菌可能不需要转主寄主也能完成生活史[10-11],最近通过接种实验和分子序列对比及种群遗传学方面的研究,确定杉李盖痂锈菌是转主寄生的真菌,可以远距离传播,以有性生殖为主[14,25-26]。
春季,云杉球果上的锈孢子器破裂,散出锈孢子。锈孢子随风传播,侵染稠李叶片后,形成双核的夏孢子和冬孢子,夏孢子可重复侵染稠李叶片,直至秋季,冬孢子在落叶上越冬。第二年春季,冬孢子萌发产生担子和担孢子,单核的担孢子随风或气流传播[17],侵染云杉雌花(球果)。担孢子萌发产生的菌丝侵入球果中心的轴,并沿轴扩散[2],在种鳞上形成性孢子器。性孢子器破裂释放出性孢子,性孢子和受精丝融合形成双核菌丝,然后形成具有厚壁的锈孢子器。树上宿存2年或3年的球果种鳞上的锈孢子器也能释放出具有萌发能力的锈孢子[4]。
2.1.4 发病规律
在芬兰,夏孢子在稠李萌芽几周后形成,大约5月下旬至6月中旬达到高峰;冬孢子在6月中旬形成,不经过休眠立即萌发,侵染云杉球果,7月下旬结束。6月下旬,种鳞上出现发白、不成熟,部分埋生在种鳞中的锈孢子器,以后,锈孢子器突破表皮,颜色加深,逐渐开始表现为红色壳状,10月份以后变为棕色[4,11]。
2.1.5 寄主抗病性
不同的云杉属植物对杉李盖痂锈菌有明显的抗病性差异。Kaitera等[13]在芬兰研究球果锈病时发现,淡红云杉P. rubens和黑云杉P. mariana的球果免疫,没有发现被侵染的球果,英格曼云杉P. engelmannii和塞尔维亚云杉P. omorika抗病,发病较轻,分别只有3%和6%的球果发病,欧洲云杉P. abies中度发病,26%的球果发病。在我国,长叶云杉P. smithiana免疫,球果上未发现杉李盖痂锈菌[28],粗枝云杉、雪岭云杉P. schrenkiana等感病[11,29]。
2.1.6 防治
Kaitera等[7]认为清除种子园中的感病球果可以减轻病害的发生,陈守长[16]推荐清除采种园附近的转主寄主稠李来防治病害,但孢子远距离传播使得这两种防治措施的防效有限[30]。另外,推测杉李盖痂锈菌性孢子的传播可能和昆虫相关,所以防治球果虫害也被认为是防治采种园球果锈病的主要措施之一[30]。
2.2 鹿蹄草云杉球果锈病
2.2.1 分布与危害
鹿蹄草云杉球果锈病的发生偶尔给欧洲云杉种子生产带来毁灭性危害[31]。2006年,挪威鹿蹄草云杉球果锈病大发生,严重影响了云杉种子生产,但Kaitera等[13]年在芬兰调查时,研究植物园内所有的云杉属球果上均未发现鹿蹄草金锈菌。相对于欧洲,鹿蹄草云杉球果锈病在北美地区病害发生更为普遍。1969年鹿蹄草球果锈病在美国犹他州严重发生,海拔2 200~2 400 m的科罗拉多河沿岸,40%~100%蓝云杉P. pungens发病,20%~67%的球果被感染[3]。在加拿大亚伯达地区,2001年前后,鹿蹄草云杉球果锈病发病率0%~25%[32]。Hedgcock[33]发现在鹿蹄草金锈菌侵染英格曼云杉球果后,幼小球果停止发育,严重影响种子产量;Sutherland[34]试验证明,鹿蹄草金锈菌侵染后,黑云杉种子产量和质量降低,发芽能力也受到影响;美国和加拿大均有报道表明,危害严重的球果不产生种子[3]。另外,由于病菌入侵引起球果流脂,还影响了种子的散播[35]。
2.2.2 症状
侵染初期,云杉球果的鳞片变黄、流胶,随后种鳞背面产生白色至黄色的锈孢子器,锈孢子成熟后,种鳞提前张开,从鳞片间散出黄色粉末状的锈孢子,在云杉树下的植物上也可见黄色的锈孢子[3]。
2.2.3 病原
此次样本数据资料中的68例患者均来自于2016年1月—2018年10月在本院进行老年股骨颈骨折治疗的患者,其中男性患者以及女性患者的比例为33:35,年龄区间界定在60岁至78岁的区间范围内,中位年龄(69.8±1.7)岁。就诊区间界定在1小时至9小时的区间范围内,中位就诊时间(5.1±1.5)小时。68例患者中共包括高坠伤患者36例,交通致伤患者20例,摔伤患者12例。所有患者的基本资料均满足此次样本研究需要,且知情同意书均经过患者以及患者家属签署。
鹿蹄草金锈菌隶属金锈菌属,是转主寄生的长生活史锈菌,共产生5种孢子,分别着生在云杉球果种鳞的背面和鹿蹄草叶柄、叶片和花上,部分地区不产生或很少产生冬孢子[32]。
2.2.4 发病规律
鹿蹄草金锈菌侵染鹿蹄草和云杉球果。鹿蹄草是多年生常绿草本植物,病菌以菌丝体在鹿蹄草叶片上越冬。第二年6月中旬,越冬叶子背面覆盖直径约1 mm的泡囊,包含冬孢子和夏孢子(大多数为夏孢子),叶片在孢子出现后枯死。夏孢子是无性孢子,在鹿蹄草上重复侵染。5月未至6月初,鹿蹄草叶片上产生的冬孢子不经过休眠,直接萌发产生担孢子,侵染云杉球果,这时鹿蹄草叶片上孢子最多,同时也是鹿蹄草在林下最多的时候。大量被侵染的球果可在8~9月发现[3],10月锈孢子开始飞散侵染鹿蹄草。
鹿蹄草叶片着生的角度与锈病的发生有密切的关系。Cater[36]发现鹿蹄草叶片的生长角度越倾向于垂直,越有利于锈病的发生,可能是倾向于垂直的叶片更有利于着生在叶片背面的担孢子飞散,锈菌的入侵可能改变了叶片生长的方向,也可能是倾向于垂直的叶片能吸引更多昆虫传播孢子。
环境因子影响病害发生的关键。鹿蹄草越冬叶片上出现的泡囊产生冬孢子还是夏孢子取决于环境因子,尤其是湿度。湿度大时,有利于冬孢子的形成,所以降雨有利于病菌入侵;相对湿度小,鹿蹄草所处的环境相对干燥时,有利于夏孢子的形成和病害蔓延[32]。
Tillman-Sutela[37]研究发现,被鹿蹄草金锈菌侵染的种子内部也发现病菌的孢子,特别是在珠心组织中,孢子旁边的部分细胞的细胞质已经被破坏。因此,鹿蹄草金锈菌可能阻碍了种子的形成,增加了种子的畸形率[38]。
2.2.5 寄主范围及抗病性
除了P. breweriana、P. chihitahuana和P. mexicana以外,鹿蹄草金锈菌能侵染其他所有美洲云杉树种[3],在欧洲主要侵染欧洲云杉,原产于美洲的淡红云杉 P. rubens、白云杉 P. glauca、P. pungens 和恩格尔曼云杉P. engelmannii等并没有发现鹿蹄草金锈菌,这可能是由于鹿蹄草金锈菌较低的侵染率、云杉对鹿蹄草金锈菌的抗病性,或锈菌与北美地区的锈菌有不同的致病性引起的[13]。在亚洲、俄罗斯远东地区侵染鱼鳞云杉P. jezoensis var.microsperma,在我国东北侵染鱼鳞云杉和红皮云杉P. koraiensis[7]。
2.2.6 防治
在担孢子飞散时期,对云杉喷施福美铁有很好的防治效果[39],但是担孢子飞散时期正是云杉雌球果受粉时期,喷药对种子的萌发有轻微的影响。
2.3 独丽花云杉球果锈病
2.3.1 分布与危害
独丽花云杉球果锈病只发生在北美洲海岸的地区,分布范围略超过北美云杉在太平洋中部和北部的分布区[2]。云杉球果被侵染后,不产生种子,即使形成种子,种子的质量和发芽率明显降低,畸形种子比列增加。病害周期性的发生,严重危害云杉采种园的种子生产。
2.3.2 症状
同鹿蹄草云杉球果锈病。
2.3.3 病原
独丽花金锈菌转主寄生,长生活史。性孢子和锈孢子着生在球果种鳞上,锈孢子橘黄色;夏孢子和冬孢子着生在独丽花上。独丽花金锈菌最初依据形态学、严格的寄主范围(P. sitchensis 和Moneses uniflora分别是锈菌唯一的锈孢子和夏孢子寄主)和地理分布确定为种的。但检测独丽花金锈菌的3个模式标本和从鹿蹄草上收集的夏孢子ITS序列时发现,这两个种之间的差异很小,只有1个SNP(超过3个基因)不同[40],这两种很可能是同一种。
2.3.4 侵染循环
与鹿蹄草金锈菌相似。这两种病菌的菌丝均多年生,并且在冬孢子寄主上产生夏孢子,夏孢子反复侵染,扩散到附近的寄主植物上。在没有云杉分布的地区,也经常能见到这两种锈菌[40],因此推测这两种菌可能在没有转主寄主云杉的时候,也能够完成生活史。
2.4 其他锈菌引起的球果锈病
除了上述3种主要的云杉球果锈病外,还有一些锈菌如变形金锈菌、伏鲁宁金锈菌和杜香金锈菌主要侵染云杉的芽和针叶,偶尔也在球果上发现,但危害相对较小。
2.4.1 变形金锈菌
变形金锈菌是短循环型锈菌,只产生冬孢子[41],侵染云杉新芽,受害的幼芽肥而大,浅橘黄色,抽芽时间提前3~7 d[42],从被侵染芽抽出新梢的各个部分都覆盖有冬孢子。球果被侵染后变为黄色,不再长大,很快枯死,种鳞上着生橘红色的冬孢子堆。变形金锈菌除了侵染芽、球果外,还能侵染幼苗[8];在喜马拉雅西部地区侵染印度云杉P. moringa,在日本侵染鱼鳞云杉和欧洲云杉,在我国疆地区侵染雪岭云杉P. schrenkiana[7]。付作霖[43]认为病菌可能侵染甘肃分布的主要云杉树种。
2.4.2 伏鲁宁金锈菌
伏鲁宁金锈菌是转主寄生的真菌,侵染云杉的芽,主要侵染顶芽,偶尔侵染1或2个侧芽。被侵染芽萌发产生的新叶全部罹病,叶片变为黄色,畸形,病叶上着生性孢子器和锈孢子器[9]。病菌还能侵染幼苗,严重发病时能阻碍幼苗生长。在阿拉斯加,发现伏鲁宁锈菌还可以侵染云杉的球果,被侵染的球果不产生种子,病害严重时可影响亚高山地区云杉林的自然更新。伏鲁宁金锈菌缺少夏孢子阶段,冬孢子着生在杜香Ledum spp.丛枝状枝条抽出的新叶上[2,43]。受到寄主分布范围的限制,伏鲁宁锈菌只分布在林线附近。
伏鲁宁金锈菌锈菌的冬孢子和锈孢子出现在不同年份,Crane[45]通过接种试验推测,春季云杉芽萌动时,杜香上的冬孢子萌发产生担孢子,担孢子侵入云杉萌发的新叶,然后通过叶片蔓延至春季或夏季形成的休眠芽中,以菌丝在在休眠芽中越冬,第二年春天休眠芽萌发形成的新叶全部表现症状。也可能是担孢子侵入新展开芽的顶端后潜伏侵染,直到第二年春节休眠芽开始萌动时,病菌开始生长、蔓延。这和其他金锈菌属真菌生活史不同。
伏鲁宁金锈菌在加拿大主要侵染黑云杉P.mariana和白云杉P. glauca[9],在我国侵染红皮云杉和鱼鳞云杉[7,46]。
2.4.3 杜香金锈菌
杜香金锈菌侵染云杉针叶引起叶锈病。Kaitera等[10]在欧洲云杉球果种鳞上发现杜香金锈菌锈孢子以来,杜香金锈菌已经在多种云杉球果上发现。但杜香金锈菌只在球果个别种鳞上出现,并不影响球果种子的形成,也不影响种子的质量和产量[7,11]。
杜香金锈菌的针叶寄主非常广泛[47],但病菌不能侵染部分针叶寄主上的球果。如淡红云杉和黑云杉的球果上没有发现杜香金锈菌的锈孢子器[12]。
侵染云杉球果锈菌的生活史简要归纳为表2。

表2 云杉球果锈病病原菌生活史
3 小结与展望
随着云杉人工林面积的不断扩大,球果锈病的发生日益严重,尤其是在亚高山地区,球果锈病的发生严重降低了云杉种子的产量和质量,阻碍种子的散播,影响云杉人工林和天然林的自然更新。目前,球果锈病的研究已经从不同角度展开,对球果锈病的主要病原菌种类及其可能的生活史已经初步了解,并在采种园也开展了一些防治工作。
但在病原方面,主要病原菌的生活史需要进一步研究,锈菌完成生活史是否需要转主寄主还需进一步探讨;危害方面,已经明确杉李盖痂锈菌和鹿蹄草金锈菌均能阻碍种子的形成并降低种子的质量和产量,鹿蹄草金锈菌可能影响种子的形成,增加种子的畸形率,但这些研究都是通过测定种子来间接证明的,需要进一步研究确定病菌侵染干扰种子发育;抗性方面,欧美学者基本确定了几种球果锈病病原菌的寄主范围,并发现不同云杉树种对杉李盖痂锈菌的抗性有明显的差异,甚至有几种云杉对杉李盖痂锈菌免疫,也发现鹿蹄草金锈菌在欧洲并不侵染原产于美洲的几种云杉属植物[10,12],但抗性机制的研究尚未开展。病害防治方面,云杉球果套袋对稠李云杉球果锈病和鹿蹄草云杉球果锈病两种主要的锈病都有很好的防治效果,但是套袋在阻止担孢子传播的同时,也阻碍了球果正常发育[48];喷药防治也取得了很好的防效很好[39],但由于锈菌的生活史和是否需要转主寄主等问题还需探讨,预测预报研究尚未开展,如何经济有效的防治病害,避免防治措施带来的负面影响,制定合理的防治策略还需进一步研究和探讨。
