广西乐业野生春兰表型遗传多样性及利用评价
2022-04-27曾艳华龙蔷宇何荆洲李秀玲范继征卜朝阳
曾艳华 龙蔷宇 何荆洲 李秀玲 范继征 卜朝阳




摘 要:为了解广西乐业野生春兰种质间的亲缘关系,提高其利用效率,对66份春兰种质资源的43个表型性状(13个数量性状和30个质量性狀)进行多样性和相关性分析,对乐业县各来源的春兰资源进行聚类分析,并从表型性状主成分分析的基础上对其进行综合评价。结果表明:春兰种质表型多样性丰富。数量性状变异程度较高,变异系数()为11.62%~41.56%,其中花梗长的值最高。质量性状变异系数为9.26%~55.7%,以开花习性、唇瓣主色和花瓣形状的值居高。说明花梗长、开花习性、唇瓣主色和瓣形都是春兰观赏性状的重点选育方向。相关性分析结果表明,花侧萼片宽(SSW)和花中萼片宽(SMW)、花侧萼片长(SSL)和花中萼片长(MSL)、花侧萼片形状(SSS)和花中萼片形状(MSS)、花瓣内侧主色(PIMC)和花萼片主色(SMC)之间呈显著正相关,相关系数分别为0.7951、0.7648、0.771、0.741;花侧萼片夹角(SSA)和侧萼片先端形状(SSPS)、唇瓣形状(LS)和中萼片先端形状(SSPS)、唇瓣色斑数量(LSN)和叶色(LC)之间则呈显著负相关,相关系数分别是–0.4557、–0.4316、–0.4181。数量性状和质量性状主成分分析分别提取5个和11个主成分,共10个因子,累计贡献率分别为78.193%和73.023%,可反映乐业春兰的总体形态表现;聚类分析结果显示,在欧氏距离为18.77处可将66份春兰种质划分为4组,其中第Ⅰ组只有一个55号品种,第Ⅱ组包含9个品种,主要以来源地相同为主,第Ⅲ组28份种质以红色花为主,第Ⅳ组28份种质,主要特征是花朵较大。研究筛选出19138-38、19138-40、18011、18007、19134-1共5份种质综合评价较高。该结果为了解广西野生春兰种质间亲缘关系及其评价与利用提供理论依据。
关键词:春兰;表型多样性;数量性状;质量性状;主成分分析中图分类号:S330 文献标识码:A
Phenotypic Genetic Diversity and Utilization Evaluation of Wild Germplasms from Leye County, Guangxi, China
ZENG Yanhua, LONG Qiangyu, HE Jingzhou, LI Xiuling, FAN Jizheng, BU Zhaoyang
Flowers Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
To understand the relationship among germplasms from Leye County, and to improve the utilization efficiency, genetic diversity and characteristic relativity analysis were conducted based on 43 phenotypic traits, including 13 qualitative characters and 30 quantitative characters of 66 germplasms. The resources were cluster-analyzed and comprehensively evaluated based on the principal component analysis of the phenotypic traits. The results showed that the phenotypic diversity of germplasms was rich and the trait variation was high. The coefficient of variation () of quantitative characters ranged from 11.62% to 41.56%, and the highest value of was detected in peduncle height. The of qualitative traits ranged from 9.26% to 55.7%, and the value of flowering habit, lip main color and petal shape were relatively high. The results indicated that the growing peduncle height, the increasing number of flowers, the stamen petalization lip main color and petal shape were the key breeding directions of spring orchid ornamental traits. Correlation analysis indicated that there was strong positive correlation between side sepal width (SSW) and medium sepal width (MSW), side sepal length (SSL) and medium sepal length (MSL), side sepal shape (SSS) and medium sepal shape (MSS), petal inner main color (PIMC) and sepal main colour (SMC). The correlation coefficient was 0.7951, 0.7648, 0.771 and 0.741, respectively. By contrary, there was strong negative correlation between side sepal angle (SSA) and side sepal preend shape (SSPS), lip shape (LS) and medium sepal preend shape (MSPS), lip spot number (LSN) and leaf color (LC), and the correlation coefficient was –0.4557, –0.4316, –0.4181, respectively. Five and eleven principal components with 10 factors were extracted using the principal component analysis (PCA) of both quantitative and qualitative traits, and the cumulative contribution rate was 78.193% and 73.023% respectively. The phenotypic factors contained in the principal components could be used as the main character indexes for germplasm innovation and parent selection for breeding ornamental spring orchid. Based on cluster analysis, The 66 spring orchid accessions were classified into four major groups when the euclidean distance was about 18.77. There was only one accession, C18019-3 in Class Ⅰ, and there were 9 accessions, mainly of the same origin, in Class Ⅱ. There were 28 accessions in Class Ⅲ and most of them were red flowers. There were 28 accessions in Class Ⅳ, and most of them had larger flowers. 19138-38, 19138-40, 18011, 18007 and 19134-1 were selected after comprehensive evaluation. The research would provide theoretical references for understanding the relationship between wild spring orchid germplasms in Guangxi and provide scientific basis for the evaluation and full utilization of resources.
; phenotypic diversity; quantitative trait; qualitative trait; principal component analysis
10.3969/j.issn.1000-2561.2022.04.011
春兰()又名草兰、朵朵香,属于兰科(Orchidaceae)兰属地生兰,在中国分布很广,北自秦岭,南至广东,西自云南,东至浙江、台湾。多生存于海拔300~2500 m阴坡的林缘、疏林间、灌木丛下或草丛中。广西的乐业县位于广西西北部,地处云贵高原东南麓,年平均气温16.3℃,属亚热带湿润气候区,特别适合兰科植物的生长,被称为中国野生兰花之乡。据统计,该县境内有野生兰科植物52属156种,是一个名副其实的“野生兰花园”。春兰在乐业多分布于海拔700~1300 m通风透光的林下、林缘或多石湿润的山坡上、灌丛中。分布较广,但较为散生,多15~30株一个居群,在花形、花色上面多样性非常丰富。对于众多的野生兰花资源,多被人为采挖、贩卖,却很少对其进行种质创新和育种研究。
表型性状是植物生长的最直观表现,通过各种技术方法开展表型鑒定是了解、利用和管理种质资源的基础,在特定区域环境下,种质表型性状表达多数具有稳定性与优异性特点,这类种质可成为区域化育种的重要基础材料。利用表型性状研究群体的遗传多样性是种质资源研究的重要组成部分。兰科植物方面,铁皮石斛和虾脊兰有表型多样性研究的相关报道;孙海芹等通过濒危独花兰的表型变异揭示其适应性的意义;程浩等、周妍慧等、李宗艳等通过表型性状研究了硬叶兜兰和杏黄兜兰居群多样性;陈和明等利用表型性状对蝴蝶兰成功开展分类研究;李全健等揭示了扇脉杓兰居群表型变异与土壤养分因子的相关性。兰属植物中建兰、蕙兰、寒兰等野生资源及多种栽培品种都开展了表型多样性研究,并利用表型性状进行品种观赏性状综合评价,构建核心种质库。春兰在我国栽培历史悠久,其形态性状表现出了极高的多样性。罗涛对重庆地区野生春兰的营养器官8项形态性状指标和繁殖器官的25项形态性状指标进行方差分析和聚类分析表明:春兰在种群内和种群之间都存在较大的遗传变异,但主要存在于种群之内。陈君梅等研究了53份秦岭地区野生春兰和蕙兰表型性状遗传多样性,表明秦岭地区野生蕙兰比春兰具有更丰富的形态多样性。张韶伊等对10个春兰栽培品种表型性状进行分析。傅巧娟等对120个春兰栽培种表型性状进行分析,表明春兰种质表型多样性丰富,聚类分析将种质分出了素心和大花2类。
本研究对广西乐业收集的66份野生春兰种质资源遗传多样性进行分析,从表型性状上揭示了乐业野生春兰遗传结构和亲缘关系的远近,为野生春兰资源的保护、优异性状的发掘和开发利用提供重要理论依据。
材料与方法
材料
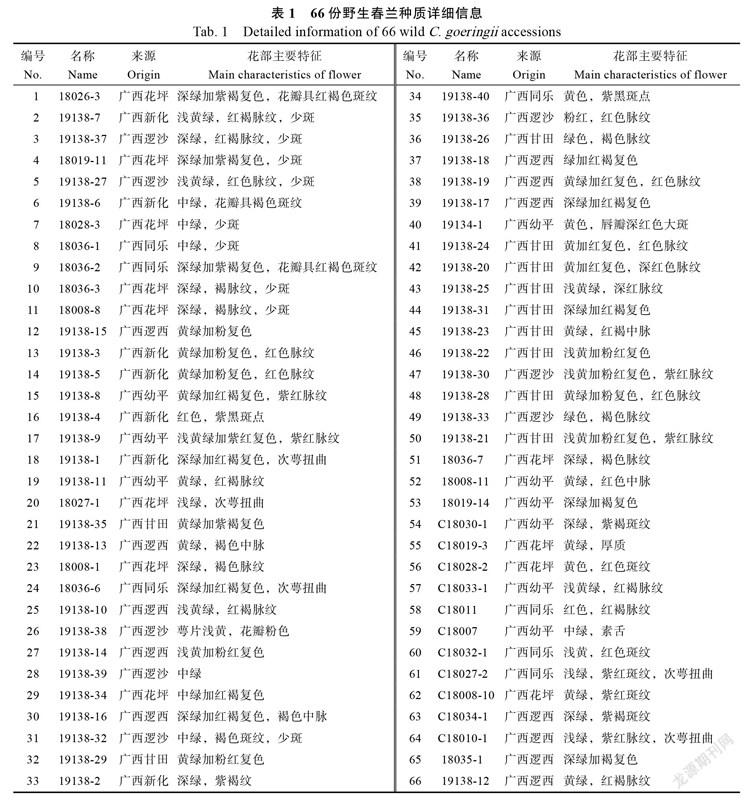
从广西乐业县采集了66份野生春兰样本(表1),包括花坪、同乐、新化、逻西、逻沙、幼平、甘田,共7个居群。采集样本全部置于广西农科院花卉研究所国兰种质资源圃迁地保存,2019—2021年对各样本开花性状进行观测和记载。
方法
1.2.1 数量性状数据处理方法 参考春兰DUS测试指南(NY/T 2437—2013)性状描述标准,选取春兰的13个数量性状,用直尺和游标卡纸测量数据,包括叶长(leaf length,LL)、叶宽(leaf width,LW)、花梗粗(stem width,SW)、花梗长(stem length,SL)、花中萼片长(medium sepal length,MSL)、花中萼片宽(medium sepal width,MSW)、花萼片厚(sepal thickness,ST)、花侧萼片长(side sepal length,SSL)、花侧萼片宽(side sepal width,SSW)、花瓣长(petal length,PL)、花瓣宽(petal width,PW)、唇瓣长(lip length,LIL)、唇瓣宽(lip width,LIW),每份种质选3株健康植株盛花期进行测量。运用Microsoft Excel 2003软件计算各个数量性状的平均值(Mean)、最大值(Max)、最小值(Min)、极差()、标准差()及变异系数(),对数量性状进行变异分析。运用SPSS 16.0软件,采用Pearson简单相关系数对春兰的数量性状进行相关性分析。Pearson简单相关系数是在变量的相关分析过程中常用的相关系数之一,常用来衡量定距变量间的线性关系。运用SPSS 16.0软件对国兰的数量性状进行系统聚类,使具有共同特征或关系密切的性状聚在一起,以便从多个数量性状中选出具有代表性的性状作分析,减少分析性状的个数。
1.2.2 质量性状数据处理方法 参考春兰DUS测试指南(NY/T 2437—2013)性状描述标准,选取30个质量有序多态性状目测描述,包括叶色(leaf color,LC)、叶形状(leaf shape,LS)、叶横截面形状(leaf cross-section shape,LCSS)、花苞片颜色(bracts color,BC)、开花习性(flowering habit,FH)、花姿态(flower posture,FP)、花中萼片形状(medium sepal shape,MSS)、花中萼片纵切面形状(medium sepal vertical shape,MSVS)、花中萼片姿态(medium sepal posture,MSP)、花中萼片先端形状(medium sepal preend shape,MSPS)、花侧萼片形状(side sepal shape,SSS)、侧萼片姿态(side sepal posture,SSP)、花侧萼片先端形状(side sepal preend shape,SSPS)、花侧萼片夹角(side sepal angle,SSA)、花萼片颜色(sepal color,SC)、花萼片主色(sepal main colour,SMC)、花萼片条纹颜色(sepal stripe color,SSC )、花瓣形状(petal shape,PS)、花瓣纵切面形状(petal vertical shape,PVS)、花瓣内侧主色(petal inner main color,PIMC )、唇瓣形状(lip shape,LS)、唇瓣纵切面形状(lip vertical shape,LVS)、唇瓣主色(lip main color,LMC)、唇瓣色斑颜色(lip Spot color,LSC)、唇瓣色斑数量(lip spot number,LSN )、唇瓣色斑图案(lip spot pattern,LSP)、花香味(Fragrance,FG)、花瓣先端形状(petal preend shape,PPS)、花瓣内侧条纹颜色(petal inner stripe color,PISC)、唇瓣色斑大小(lip spot size,LSS)。数据处理方法同数量性状。选取6个质量二元性状描述统计,利用Microsoft Excel 2003软件对二元性状进行概率分布分析。
数据处理
运用SPSS 16.0软件计算欧氏距离,采用最短距离法对春兰品种资源进行聚类,使具有共同特征或关系密切的国兰种质聚在一起,从形态学角度探讨春兰的亲缘关系。
结果与分析
数量性状描述性统计和相关分析
66份春兰种质的13个数量性状数据描述统计见表2。结果显示,春兰具有广泛的形态学变异,部分性状有较高的变异系数,变异度最大的为花梗长(41.56%),其次是叶长(27.21%)、花萼片厚度(26.83%)、花瓣宽(22.74%)、花侧萼片宽(19.67%)、花中萼片宽(19.67%)叶宽(19.07%),其余性状的变异系数相对较低,但基本都在15%以上,变异最小的是花中萼片长(11.62%)。花梗最长的是C18019-3(21.33 cm),其次是19138-16(19.27 cm)和19138-39(18.13 cm),最短的是18008-1(2.83 cm)。葉长最长的是19138-17(77.87 cm),其次是19138-2(72.17 cm)和19138-16(71.67 cm),最短的为C18019-3(15 cm)。花萼片厚度最厚的是C18019-3(0.88 mm),其次是C19138-40(0.83 mm)和C19138-33(0.81 mm),花萼片最薄的是C18008-8(0.22 mm)。花瓣最宽的是C18030-1(1.92 cm),其次是C19134-1(1.53 cm)和C19138-33(1.50 cm),最小的是C19138-28(0.45 cm)。花侧萼片宽最大的是C19134-1(1.25 cm),其次是C18030-1(1.23 cm)和C19138-39(1.22 cm),最小的是C18036-3(0.51 cm)。花中萼片最宽是的C19134-1(1.53 cm),其次是C19138-39和C18030-1(均是1.40 cm),最小的是C18007(0.57 cm)。叶宽最大的是C19138-28(1.21 cm),其次是C19138-31(1.13 cm),最小的是C18019-3(0.53 cm)。
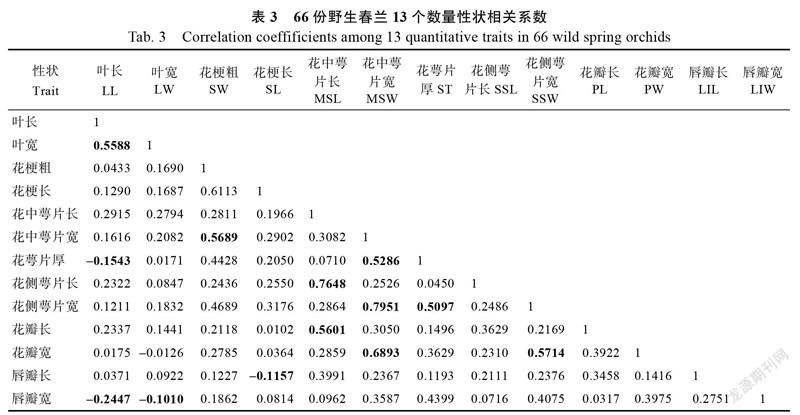
相关分析结果显示,13个数量性状多数存在较强的正相关(表3)。相关系数最高的是花侧萼片宽和花中萼片宽(0.7951),其次是花侧萼片长和花中萼片长(0.7648)。在花中萼片宽和花瓣宽(0.6893)、花瓣宽和花侧萼片宽(0.5714)、花中萼片宽和花梗粗(0.5689)、花瓣长和花中萼片长(0.5601)、叶宽和叶长(0.5588)之间也有较高的相关系数。花萼片厚和花中萼片宽(0.5286)、花侧萼片宽和花萼片厚(0.5097)之间也存在较高的正相关。相反,唇瓣宽和叶长之间存在最高的负相关(–0.2447),叶长和花萼片厚(–0.1543)、花梗长和唇瓣长(–0.1157)、叶宽和唇瓣宽(–0.101)之间也存在相对较强的负相关。
2.2 数量性状变量主成分分析
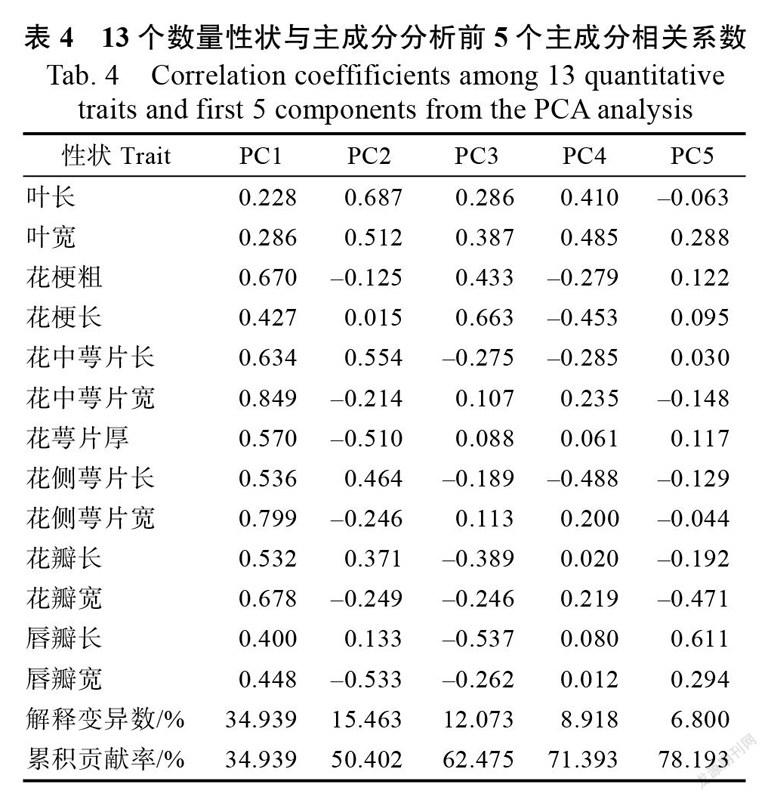
按照特征根大于1,提取前5个主成分,解释总变异数的78.193%(表4)。PC1解释总变异数的34.939%,花中萼片宽、花侧萼片宽、花瓣宽、花中萼片长、花梗粗5个性状与PC1相关系数较高。PC2解释总变异数的15.463%,叶长、花中萼片长、叶宽与PC2有较高相关系数,唇瓣宽和花萼片厚与PC2相关系数绝对值较高,且呈负相关。与PC3相关系数较高的性状包括花梗长、唇瓣长、花梗粗,解释12.073%的总变异数。与PC4相关系数较高的性状包括花侧萼片长、叶宽、花梗长和叶长,解释8.918%的总变异数。唇瓣长和花瓣宽与PC5相关系数较高,解释6.800%的总变异数。
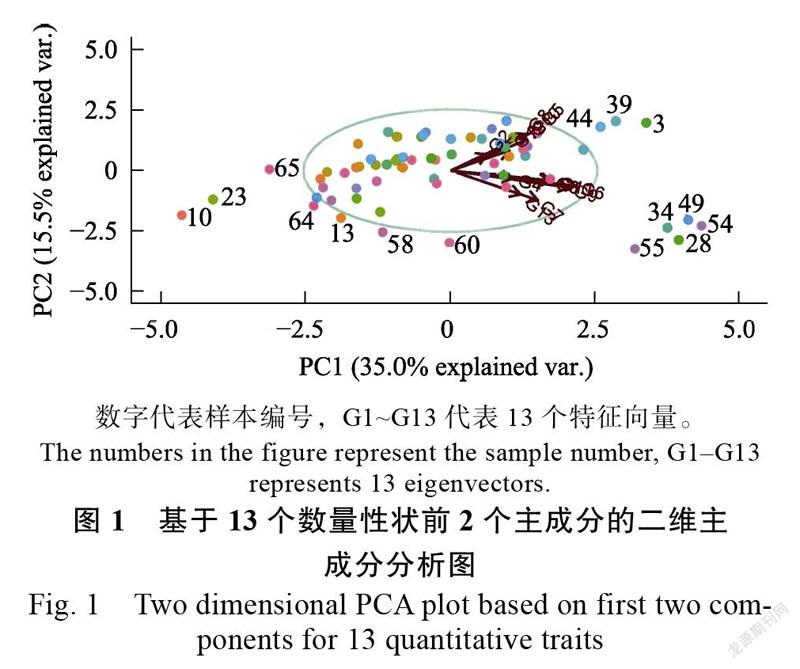
用前2个主成分PC1、PC2作PCA二维点图(图1),根据样本的表型相似性和形态学特征对
样本进行分组。如侧萼片宽最小、花梗最短的18036-3、18008-1、18035位于图最左边,对应编号10、23、65;叶宽最大、叶长最长的19138-31、19138-17、19138-37分布于图的右上角,对应编号44、39、3;而萼片较厚、侧萼片较宽、中萼片较宽、花瓣较宽的18019-3、19138-40、19138-33、19138-39、18030-1分布于图右下角,对应编号55、34、49、28、54。结果印证了花中萼片宽、花侧萼片宽、花瓣宽几个性状之间存在显著正相关,在PCA分析中有较高的载荷。
质量有序多态性状描述性统计和相关分析
由表5可知,30个质量有序多态性状通常表现出超过2种表型,显示了广泛的质量性状形态学变异。87%的多态性状的变异系数都超过20%,变异系数最高的是开花习性(55.72%);其次是唇瓣主色(51.07%)、花瓣形状(50.83%)、唇瓣形状(50.61%),唇瓣色斑图案(48.02%)、花侧萼片形状(45.16%)、唇瓣色斑大小(43.2%)、叶色(42.39%)等也有较高的变异系数;变异系数最小的是花侧萼片夹角(9.26%),唇瓣纵切面形状(13.25%)、唇瓣色斑颜色(14.54%)也有较小的变异系数。
大多数春兰种质开花习性为1朵,有14份为1朵或2朵,有4份开花习性为2朵(19138-11、19138-10、18036-7、18019-14)。唇瓣主色多数表现为黄色(35份)和白色(29份),19138-3为绿色,19134-1为红色。花瓣形状大部分表现为披针形(21份)和菱形(22份),有15份表现为长椭圆形,有8份表现为椭圆形。唇瓣形状多数表现为匙形(31份),其次是三角形(19份)和近圆
形(14份),18026-3和19138-26表现为扁圆形。唇瓣色斑图案多数表现为圆点(20份)、V或U形(24份),12份表现为斑块状,7份表现为品字形(18036-1、18036-2、19138-4、19138-5、19138-14、19138-34、18008-10),还有3份为不规则条形(18033-1、19138-12和18008-8)。唇瓣色斑大小多数为小(30份),中(28份),7份为大的色斑(18036-2、19138-10、19138-40、19134-1、19138-33、18036-7、18030-1),18007为唯一一个素舌品种,色斑大小记为4。花侧萼片夹角几乎大于180,只有19138-10为180平肩,18027-1为略小于180。
30个质量性状相关性分析,在435个相关系数中,54%为正相关(表6)。花侧萼片形状和花中萼片形状相关系数最高(0.771),其次是花瓣内侧主色和花萼片主色(0.741)、花侧萼片形状和花中萼片先端形状(0.709),花中萼片形状和中萼片先端形状(0.634)、花侧萼片先端形状和花中萼片先端形状(0.633)之间也存在显著正相关。花侧萼片夹角和侧萼片先端形状之间存在极显著负相关(–0.46),其次是唇瓣形状和中萼片先端形状(–0.43)、唇瓣色斑数量和叶色(–0.42)之间也呈显著负相关。
质量有序多态性状变量主成分分析
按照特征根大于1,提取前11个主成分,解释总变异数的73.023%(表7)。PC1解释总变异数的13.989%,花中萼片纵切面形状、花侧萼片形状、花侧萼片先端形状、唇瓣形状、花瓣先端形状5个性状与PC1相关系数绝对值较高,其中花中萼片纵切面形状和唇瓣形状与主成分呈负相关。PC2解释总变异数的10.399%,花萼片颜色、花瓣内侧主色、唇瓣主色、唇瓣色斑数量与PC2相关系数绝对值较高,其中花萼片颜色与PC2呈显著负相关。与PC3相关系数较高的性状包括叶形状、花萼片条纹颜色和花瓣内侧条纹颜色均呈正相关,解释8.330%的总变异数。花瓣形状和花瓣先端形状与PC4呈显著正相关,解释7.253%的总变异数。花萼片主色、花瓣内侧主色和唇瓣色斑颜色与PC5正相关系数较高,解释6.111%的总变异数。PC6解释总变异数的5.239%,包括叶颜色、叶横截面形状、花苞片颜色、花中萼片姿态4个相关性强的性状,其中花中萼片姿态为负相关。PC7解释总变异数的5.110%,包括开花习性和花侧萼片夹角2个正相关强的性状,侧萼片姿态1个负相关强的性状。PC8解释总变异数的4.426%,与花侧萼片夹角呈显著负相关。PC9包括花萼片颜色1个负相关较强的性状和唇瓣色斑图案1个正相关强的性状,解释总变异数的4.324%。PC10包括负相关强的唇瓣色斑图案,解释总变异数的4.126%,PC11解释3.716%的总变异数,包括花萼片主色和唇瓣纵切面形状2个正相关较强的性状。
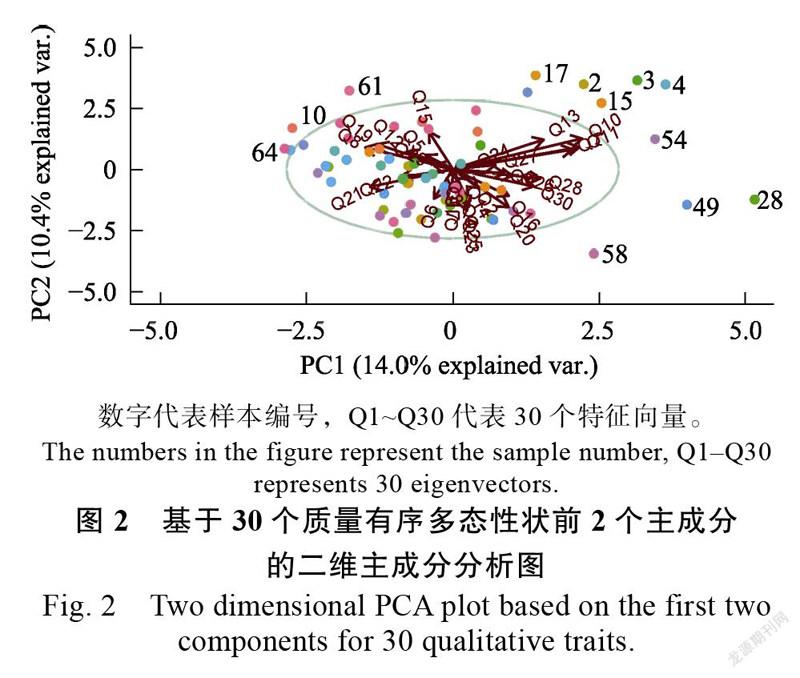
用前2个主成分PC1、PC2作PCA二维点图(图2),根据样本的表型相似性和形态学特征对样本进行分组,将相似表型的大部分种质聚为一组,具有表型离散程度比较大的样本则分散在周围。如比较分散的28、54、58,分别为19138-39、18030-1、18011,表现为花瓣椭圆形、唇斑大;61号18027-1侧萼片夹角略小于180°;19138-37、18019-11、19138-33唇瓣为斑块状色斑,对应编号3、4、49号,均散布在图的右边。花瓣形状在PC2和PC4主成分均有较高的载荷,唇瓣色斑大小在PC1中有较高的载荷,侧萼片夹角在PC7和PC8有较高的载荷,而唇瓣色斑图案在PC1、PC9和PC10均有较高的载荷。
质量二元性状概率分布分析
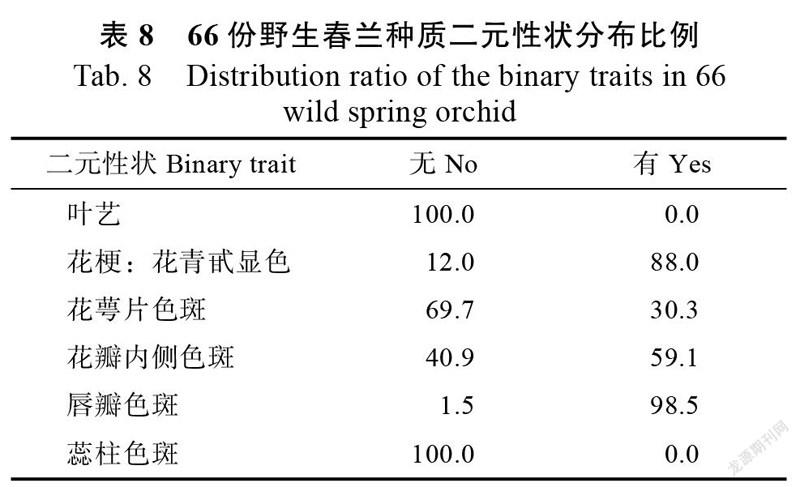
由表8可知,66个春兰种质的6个质量二元性状均有2种表现形式,即有或无。从这6个二元性状在不同春兰种质的分布比例来看,花青甙显色、花瓣内侧色斑和唇瓣色斑均以“有”占绝大多数;而花萼片色斑多数表现为“无”。供试种质全部无叶艺,具有蕊柱色斑。
系统聚类分析
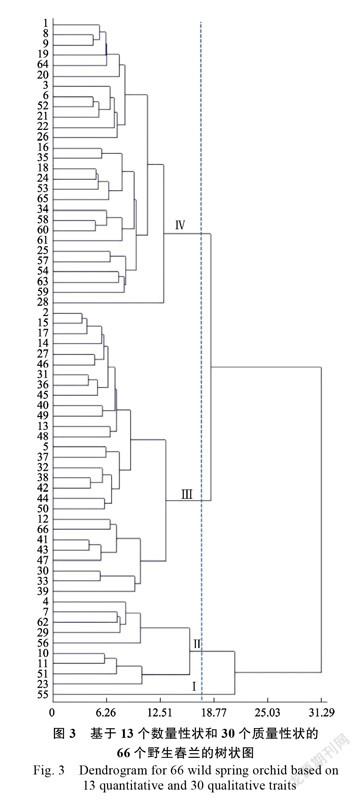
根据13个数量性状和30个质量性状数据,采用欧氏距离最短距离法对66个春兰种质作聚类分析。如图3所示,所有种质在欧氏距离为18.77的位置明显聚为4组。Ⅰ组和Ⅱ组在21.16处被分开,Ⅰ组只有55号1个品种资源,Ⅱ组包含4、7、62、29、56、10、11、51、23号共9个品种资源;Ⅲ组和Ⅳ组在18.77处被分开,均分别包含28个品种资源。遗传距离最远的是39号和55号(63.84),其次是10号和39号(60.89)及11号和39号(60.12)。
在66份春兰资源中,34、58、60、61号聚集在一起,来源地均是同乐;10、11、51、23号聚集在一起,来源地均是花坪,这是基于地域分布的特定聚集簇。部分有相似性状的种质聚在一起,形成基于表型性状的特定聚集簇,如1、8、9、19、64、20号种质聚在一起,均具有狭长而卷曲的侧萼片;2、15、17、14、27、46號聚成一小簇,属于花色偏粉的种质;5、37、32、38、42、44、50聚成一簇,属于花色偏红的种质。55号种质C18019-3单独成一组,与其他的种质遗传距离相对较远,具有最长的花梗和较厚的花瓣,与豆瓣兰相似,但基部又有春兰的关节,疑为春兰和豆瓣兰的天然杂交种。
讨论
本研究通过测量13个数量性状和30个质量性状,研究以66份野生春兰种质为代表的广西乐业野生春兰种质资源,揭示了广西乐业野生春兰种质资源具有广泛的形态学多样性。在数量性状中,花梗长的值最高,多样性更丰富,与傅巧娟等对春兰栽培种、陈君梅等对野生春兰的研究结果一致;花萼片长、花瓣长和唇瓣长变异相对稳定,说明乐业野生春兰在花的大小上变异程度不大。在质量性状中,开花习性的值最高,表现为1朵、2朵、1或2朵,其次花瓣和唇瓣的形状、颜色、唇瓣色斑图案等春兰主要观赏性状均有丰富的多样性,在花色、瓣型选择育种上有利用价值。
性状相关分析结果显示,有9对数量性状和5对质量性状间的关联较为紧密,根据这些表型性状进行选择育种时,会增加后代遗传的不可预知性及复杂性,可结合进一步的分子标记辅助选择减少育种的盲目性。
主成分分析结果显示,数量性状提取的前5个主成分、质量性状提取的前11个主成分综合反映了春兰的表型,但其累积贡献率不到80%(78.193%和73.023%),说明各性状贡献率不够集中,变异具有多向性。花中萼片宽、花侧萼片宽、花瓣宽、花中萼片长、花梗粗共5个数量性状和花中萼片纵切面形状、花侧萼片形状、花侧萼片先端形状、唇瓣形状、花瓣先端形状共5个质量性状与PC1相关系数绝对值较高,可作为野生春兰种质综合评价与杂交育种亲本选择的主要表型指标。
聚类分析中,大部分种质基于表型性状如花朵大小、花萼片形状、花色相近的聚集一起,有少量种质是基于地理分布相同聚集在一起。但与传统春兰品种按瓣形进行区分的结果不尽一致,这与傅巧娟等对120份春兰栽培品种的表型性状研究结果一致。牛田等和季祥彪等利用分子标记对野生春兰的遗传多样性研究结果也表明,聚类结果与春兰的瓣形分类不一致,原因可能与春兰种质的人为选择有关。在野外调查中了解到,野生春兰遭受到的破坏较严重,外形或颜色特异的春兰一经发现就被挖走,能在当地幸存下来的春兰多为外形普通的品种。此外,在本研究中,乐业春兰的地理分布和其表型多样性之间没有明确的相关性。季祥彪等研究表明野生春兰种质的聚类具有很强的地域性;牛田等研究认为来自相同地区的春兰品种的亲缘关系较近,与本研究结果不完全一致,可能与不同地区间春兰交流引种程度有关,或者与选择的分析方法不尽一致有关。对表型性状的选择和观测鉴定易受栽培条件及生态环境影响,具有一定的局限性,有待进一步研究。
广西乐业野生春兰花朵颜色基本为绿色和黄绿色,部分为桃红色,偶有枚红色和深绿色,花色多样性丰富。花瓣尖端多为渐尖型,其次为钝尖型,锐尖型最少。唇瓣上的紫红色斑点多呈“U”型,其次为“品”字型斑和不规则斑点,偶有红色大斑块,个别为素唇无色斑。本项目筛选出19138-38、19138-40、18011、18007、19134-1共5份种质综合观赏性状佳,可以直接筛选驯化加以利用。
参考文献
- 张振臣, 陈俊标, 马柱文, 李淑玲, 汤亚飞, 邱道寿, 谭铭喜. 铁皮石斛种质资源主要表型性状的差异与相关分析[J]. 广东农业科学, 2010, 37(8): 78-80.ZHANG Z C, CHEN J B, MA Z W, LI S L, TANG Y F, QIU D S, TAN M X. Discrepancy and correlation analysis on main phenotypic traits of [J]. Guangdong Agricultural Sciences, 2010, 37(8): 78-80. (in Chinese)
- 弓 莉, 罗 建, 林 玲. 西藏虾脊兰属植物表型多样性研究[J]. 高原农业, 2020, 4(4): 351-356.GONG L, LUO J, LIN L. Studies on phenotypic diversity of in Tibet[J]. Journal of Plateau Agriculture, 2020, 4(4): 351-356. (in Chinese)
- 孙海芹, 李 昂, 班 玮, 郑晓明, 葛 颂. 濒危植物独花兰的形态变异及其适应意义[J]. 生物多样性, 2005, 13(5): 376-386.SUN H Q, LI A, BAN W, ZHENG X M, GE S. Morphological variation and its adaptive significance for , an endangered orchid[J]. Biodiversity Science, 2005, 13(5): 376-386. (in Chinese)
- 程 浩, 徐玉凤, 王文晓, 朱 俊, 贾瑞冬, 杨树华, 赵 鑫, 葛 红. 中国硬叶兜兰主要分布区居群表型多样性分析[J]. 园艺学报, 2020, 47(6): 1098-1110.CHENG H, XU Y F, WANG W X, ZHU J, JIA R D, YANG S H, ZHAO X, GE H. Phenotypic diversity of populations from main distribution areas in China[J]. Acta Horticulturae Sinica, 2020, 47(6): 1098-1110. (in Chinese)
- 周妍慧, 贾瑞冬, 杨树华, 赵 鑫, 葛 红, 罗文美, 赵新梅, 桑益恒. 杏黄兜兰居群表型多样性分析[J]. 园艺学报, 2016, 43(7): 1337-1347.ZHOU Y H, JIA R D, YANG S H, ZHAO X, GE H, LUO W M, ZHAO X M, SANG Y H. Phenotypic diversity of populations[J]. Acta Horticulturae Sinica, 2016, 43(7): 1337-1347. (in Chinese)
- 李宗艳, 李名扬. 硬叶兜兰居群表型变异研究[J]. 植物遗传资源学报, 2015, 16(4): 765-771.LI Z Y, LI M Y. Study on phenotypic variation of population[J]. Journal of Plant Genetic Resources, 2015, 16(4): 765-771. (in Chinese)
- 陈和明, 吕复兵, 李 佐, 肖文芳. 基于表型性状的蝴蝶兰种质资源分类[J]. 中国园艺文摘, 2017, 33(2): 1-7.CHEN H M, LV F B, LI Z, XIAO W F. Classification of germplasm based on phenotypic traits[J]. Chinese Horticulture Abstracts, 2017, 33(2): 1-7. (in Chinese)
- 李全健, 王彩霞, 田 敏, 李翠新. 浙江扇脉杓兰野生居群的表型性状变异及其与地理-土壤养分因子的相关性[J]. 植物资源与环境学报, 2012, 21(2): 45-52.LI Q J, WANG C X, TIAN M, LI C X. Variation of phenotypic traits of wild populations of in Zhejiang Province and its correlation with geographical-soil nutrient actors[J]. Journal of Plant Resources and Environment, 2012, 21(2): 45-52. (in Chinese)
- 王宏利, 曾艳华, 卜朝阳. 30份建兰种质资源的表型性状遗传多样性研究[J]. 热带作物学报, 2021, 42(6): 1557-1565. WANG H L, ZENG Y H, BU Z Y. Genetic diversity of phenotypic traits of 30 germplasm resources[J]. Chinese Journal of Tropical Crops, 2021, 42(6): 1557-1565. (in Chinese)
- 郭 峰, 牛立新, 张延龙. 秦岭柞水地区野生蕙兰天然群体表型多样性[J]. 北方园艺, 2010(18): 91-93.GUO F, NIU L X, ZHANG Y L. Phenotypic variation of natural populations of in Zhashui[J]. Northern Horticulture, 2010(18): 91-93. (in Chinese)
- 傅巧娟, 李春楠, 赵福康, 孙 瑶. 7 种兰属种质表型性状遗传多样性分析[J]. 分子植物育种, 2018, 16(10): 3381-3394.FU Q J, LI C N, ZHAO F K, SUN Y. Genetic diversity analysis of phenotypic traits in 7 species of [J]. Molecular Plant Breeding, 2018, 16(10): 3381-3394. (in Chinese)
- 段艳皊, 范义荣, 敖素燕, 宁惠娟, 郭雨楠. 寒兰种质资源表型性状多样性分析[J]. 中国农学通报, 2014, 30(16): 143-147.DUAN Y L, FAN Y R, AO S Y, NING H J, GUO Y N. Analysis on phenotypic character diversity of germplasm resources[J]. Chinese Agricultural Science Bulletin, 2014, 30(16): 143-147. (in Chinese)
- 敖素燕, 范义荣, 段艳皊, 郭雨楠, 宁惠娟. 31个国兰品种的数量性状分析[J]. 江苏农业科学, 2014, 42(9): 131-134.AO S Y, FAN Y R, DUAN Y L, GUO Y N, NING H J. Quantitative trait analysis of 31 species[J]. Jiangsu Agricultural Sciences, 2014, 42(9): 131-134. (in Chinese)
- 黄晓玲. 部分国兰表型多样性研究[J]. 绿色科技, 2019(9): 84-86.HUANG X L. Phenotypic diversity studies for parts of [J]. Journal of Green Science and Technology. 2019(9): 84-86. (in Chinese)
- 陈宝玲, 王华新, 陈 尔, 杨舒婷, 龚建英. AHP法对广西引种的14个国兰品种观赏性状综合评价[J]. 广西林业科学, 2015, 44(3): 296-299.CHEN B L, WANG H X, CHEN E, YANG S T, GONG J Y. AHP in ornamental evaluation of 14 introduced in Guangxi[J]. Guangxi Forestry Science, 2015, 44(3): 296-299. (in Chinese)
- 艾 叶, 陈 璐, 兰思仁, 谢泰祥, 陈 娟, 彭东辉. 基于形态学性状构建建兰品种核心种质[J]. 分子植物育种, 2019, 17(23): 7924-7934.AI Y, CHEN L, LAN S R, XIE T X, CHEN J, PENG D H. Construction of core collection of varieties based on morphological traits[J]. Molecular Plant Breeding, 2019, 17(23): 7924-7934. (in Chinese)
- 何仁锋, 陈 喆, 姜梦莹, 周淑婷, 应奇才, 冯尚国, 王慧中. 兰属植物遗传资源核心种质构建探讨[J]. 杭州师范大学学报(自然科学版), 2015, 14(2): 202-209.HE R F, CHEN Z, JIANG M Y, ZHOU S T, YING Q C, FENG S G, WANG H Z. On the core collection establishment of genetic resources[J]. Journal of Hangzhou Normal University (Natural Science Edition), 2015, 14(2): 202-209. (in Chinese)
- 王晓英, 王长宪, 华芳霞. 春兰的遗传多样性研究进展[J]. 山东林业科技, 2013(3): 96-99. WANG X Y, WANG C X, HUA F X. Review on thg genetic diversity of Rchb. f[J]. Shandong Forestry Science and Technology, 2013(3): 96-99. (in Chinese)
- 罗 涛. 重庆地区野生春兰种群分布、群落特征及形态结构分化研究[D]. 重庆: 西南大学, 2007.LUO T. Studies on population distribution, community features and external morphological as well as anatomical differentiations of wildin Chongqing[D]. Chongqing: Southwest University, 2007. (in Chinese)
- 陳君梅, 宋军阳, 韩王亚, 陈博文, 张 显. 秦岭野生春兰和蕙兰的形态多样性研究[J]. 西北农林科技大学学报(自然科学版), 2017, 45(2): 143-150. CHEN J M, SONG J Y, HAN W Y, CHEN B W, ZHANG X. Morphological diversity of wild and in the Qinling mountains[J]. Journal of Northwest A&F University (Natural Science Edition), 2017, 45(2): 143-150. (in Chinese)
- 张韶伊, 范义荣, 褚 怡, 孙玉芬, 宁惠娟. 春兰属植物表型性状的多样性研究[J]. 北方园艺, 2013(12): 68-72.ZHANG S Y, FAN Y R, CHU Y, SUN Y F, NING H J. Study on the phenotypic traits of species diversity[J]. Northern Horticulture, 2013(12): 68-72. (in Chinese)
- 傅巧娟, 李春楠, 张晓莹, 赵福康. 基于表型性状的春兰种质遗传多样性分析[J]. 农学学报, 2020, 10(11): 65-71. FU Q J, LI C N, ZHANG X Y, ZHAO F K. Genetic diversity analysis of phenotypic traits of [J]. Journal of Agriculture, 2020, 10(11): 65-71. (in Chinese)
- 牛 田, 张 林, 王厚新, 李承秀, 聂 硕, 朱翠翠, 王长宪. 利用SRAP标记分析春兰种质资源遗传多样性[J]. 农学学报, 2014, 4(8): 53-58. NIU T, ZHANG L, WANG H X, LI C X, NIE S, ZHU C C, WANG C X. Genetic diversity analysis of 's germplasm resources based on SRAP markers[J]. Journal of Agriculture, 2014, 4(8): 53-58. (in Chinese)
- 季祥彪, 王国鼎, 乔 光, 康冀川, 宋常美, 文晓鹏. 贵州野生春兰遗传多样性的RAPD分析[J]. 华中农业大学学报, 2008, 27(2): 297-302.JI X B, WANG G D, QIAO G, KANG J C, SONG C M, WEN X P. Genetic diversity of the wild (Orchidaceae) populations from Guizhou as revealed by RAPD markers[J]. Journal of Huazhong Agricultural University, 2008, 27(2): 297-302. (in Chinese)
