河南济源人工引水渠隧道蝙蝠冬眠生态学特征
2022-04-25戴文涛尹香菊冷海霞江廷磊
刘 森, 戴文涛, 尹香菊, 冷海霞, 朱 越, 江廷磊,*
1 河南师范大学生命科学学院, 新乡 453007
2 河南理工大学资源环境学院, 焦作 454000
3 吉林省动物资源保护与利用重点实验室, 东北师范大学, 长春 130117
蝙蝠是翼手目(Chiroptera)动物,它们捕食昆虫或传播种子,对维护生态系统平衡具有重要作用。然而,受到全球变暖、生境破碎化、农药富集、疾病等因素影响,蝙蝠种群数量锐减[1]。由于蝙蝠越冬后繁育后代,深入了解蝙蝠冬眠生态学特征,特别是蝙蝠冬眠种群数量变化趋势及其冬眠场所环境情况,对准确评估蝙蝠物种濒危等级、种群繁衍和发展趋势至关重要,是量化种群变化响应生态扰动的基本前提。
国外研究者在蝙蝠冬眠种群监测、冬眠栖息地选择等方面做了大量工作。通过长期监测发现,一些冬眠物种种群数量出现年度增长趋势,比如,意大利西北部52个冬眠场所的6种蝙蝠种群数量在1992—2019年期间增长了3.5%—15%[2],而有些物种种群数量则由于疾病或者人类活动干扰等因素而锐减,如真菌诱导的白鼻综合症导致北美Myotislucifugus大量死亡[3];20世纪80年代美国西北部Shoshone Desert管理区洞穴旅游的兴起,致使Corynorhinustownsendii种群数量减少[4]。对众多冬眠栖息地蝙蝠数量和环境数据分析发现,蝙蝠倾向冬眠于微气候相对稳定的场所[5]。同时,冬眠场所结构,包括空间大小、地形、出口数量、气流等,也会影响到蝙蝠种群数量[6—7]。
在我国,有关蝙蝠冬眠的研究相对较少,包括绒山蝠体内贮脂在冬眠期消耗情况[8],中华山蝠的冬眠阵长度、觉醒后呼吸频率变化、冬眠环境温度等生理特性和生态习性[9],大菊头蝠的冬眠栖点特征[10],以及西南鼠耳蝠和皮氏菊头蝠的冬眠生态学特征(冬眠数量、冬眠姿势、栖点位置,以及栖点温度与冬眠体温相关性)[11]等。然而,目前鲜有对蝙蝠冬眠种群数量进行长期监测的研究报道。
2005年2月,赵黎明[12]发现河南省济源市布袋沟水库引水渠隧道中有大量蝙蝠冬眠(超过1600只)。该引水渠包含多个长短不一的隧道,且隧道内部有流水,可为蝙蝠提供众多适宜冬眠场所。然而,由于缺乏相关数据,尚不清楚在这种湿润环境下,蝙蝠冬眠种群数量是如何变化的。2017—2020年期间,每年1月份对该引水渠隧道进行调查,初步分析蝙蝠冬眠生态学特征,包括:1)冬眠蝙蝠种类组成、种群数量现状,以及变化趋势;2)冬眠方式、栖点位置,以及栖点温度与蝙蝠体温的关系;3)环境因子对蝙蝠冬眠场所选择的影响。研究工作旨在丰富我国蝙蝠冬眠种群生态学的基础数据,并为我国蝙蝠保护策略及其冬眠栖息地管理决策制定提供重要的科学依据。
1 研究地区与方法
1.1 研究地区概况
布袋沟水库引水渠位于华中北部地区、太行山南段东麓的太行山猕猴国家级自然保护区,行政区划隶属于河南省济源市,该区域地貌属流水作用的断块中山,区内峡谷、沟壑纵横,气候类型为大陆性季风气候,年平均气温13.28 ℃,年平均降水量为 695 mm,平均无霜期为 190 d[13]。森林植物类型以针叶林、针阔混交林、落叶阔叶林、半常绿阔叶林为主,多为天然次生林。动物主要包括太行山猕猴、金钱豹、林麝等国家重点保护动物[14]。
布袋沟水库引水渠隧道由长短不一的77个隧道构成,从上游至下游,依次编号C1—C77。总长约13 km,最短约10 m,最长约770 m。长度≤ 60 m的隧道有34个,占总数的44%,其中24个位于C1—C29之间。每个隧道首尾均有一个洞口,内部曲折,隧道高约2—5 m,宽约2—4 m (图1)。C56与C57衔接之处,设置有闸门,将水截流用于发电,因而从C57开始,水位开始降低(图1),隧道温度逐步升高。
部分明渠采用人工砌石封顶(图1),组成隧道的一部分,因而形成较多的长隧道,比如有16个长度超过300 m的隧道均存在多个人工砌石处。这种拱形顶厚约50—70 cm,该区域受外界环境影响较大,有极少数蝙蝠冬眠于此(图1)。

图1 引水渠隧道特征
1.2 研究方法
1.2.1蝙蝠种类、数量、冬眠方式、栖点位置调查
2017—2020年期间,每年的1月份调查了冬眠蝙蝠的种类、数量、冬眠方式和栖点位置。所有调查人员认真检查每个石缝和凹窝,对不确定的个体需轻轻摘下认真鉴定,之后原地放回,以减少漏查和鉴定错误的几率。对个体较多的聚集群体拍照,后续通过照片确定个数。参照龚小燕等[15]定义,将蝙蝠在某一隧道内的具体栖挂或栖卧位点称为“栖点”,测量栖点到隧道口的距离。调查过程中,尽量降低人为噪音,并避免非必要的直接碰触,以减少对蝙蝠的刺激。
1.2.2影响蝙蝠冬眠隧道选择的环境因子测量
考虑到长度、高度和水深影响冬眠空间,风可以改变微气候的稳定性,声强差异可能干扰蝙蝠(对声音敏感)冬眠,水温和环境温度(空气温度)可能会影响蝙蝠冬眠的适宜度,2020年1月,本次研究对历年均有蝙蝠冬眠的隧道,测量了上述环境因子数据。除隧道长度外,所有参数每30 m测一组数据,取平均值。风速由数字式风速计(TM856D)测定,声强由数字式声级计(TD1357)测定。
1.2.3隧道不同位置环境温度、相对湿度的测量
为评估隧道不同位置环境温度和相对湿度的差异性和稳定性,选择一个长约310 m、包含人工砌石区域且有较多蝙蝠分布的隧道(C41),对其不同位置进行为期3个月(2019年10月15日—2020年1月15日)的温度和相对湿度监测(华图USB温湿度记录仪HE173,温度精度± 0.5 ℃,相对湿度精度± 3%),每6 h获取一组数据,共获得372组数据。依据前期调查的蝙蝠分布数量情况,共设置了6个位置,包括距离洞口0 m、30 m、70 m、130 m、160 m (隧道正中间)、人工砌石处等位点。0 m、30 m和人工砌石处蝙蝠数量< 3只,其余位点均有较多蝙蝠冬眠(超过10只)。
1.2.4蝙蝠体温与栖点温度测量
将Fluke红外测温仪(62MAX,精度± 1.5 ℃)置于离蝙蝠背部约5 cm处测量体温,在距蝙蝠约2 cm处测定栖点岩壁温度,均测3次取平均值,用于评估栖点温度与蝙蝠体温的相关性。冬眠位置过高或苏醒的个体不进行测量。
1.3 内业数据处理
统计并计算距离洞口不同位置的环境温度、相对湿度的变异系数,以判断其波动程度。采用多元线性回归分析方法,评估环境因子与蝙蝠冬眠数量的相关性程度,并通过逐步回归获取显著影响蝙蝠冬眠场所选择的关键环境因子。回归分析用于评估栖点温度与蝙蝠体温的相关性,独立样本t检验判别组间差异显著性。所有统计分析通过SPSS 22和R 4.1.0完成。
2 结果
2.1 种类组成和种群数量的年度变化
通过外部形态鉴定,在该水渠冬眠的蝙蝠一共有2科5属7种(图2),包括菊头蝠科的马铁菊头蝠(Rhinolophusferrumequinum)和小菊头蝠(R.pusillus),蝙蝠科的华南水鼠耳蝠(Myotislaniger)、白腹管鼻蝠(Murinaleucogaster)、金管鼻蝠(Mu.aurata)、奥氏长耳蝠(Plecotusognevi)和亚洲宽耳蝠(Barbastellaleucomelas)。
冬眠蝙蝠总数呈历年增长趋势,即从2017年774只到2020年1092只(图3)。马铁菊头蝠数量占绝对优势(约52%—73%),2017年数量最低,有407只,逐年增至765只。小菊头蝠数量次于马铁菊头蝠(约占19%—37%),2017—2019年期间从288只降至183只,2020年又增至247只。华南水鼠耳蝠数量约为总数的5%—8%,其余物种数量总和不足总数的3%,其中2018年未发现奥氏长耳蝠,2020年未发现金管鼻蝠。除马铁菊头蝠数量逐年增加外,其余物种数量在这4年期间处于增、减波动状态。

图3 2017—2020历年每种蝙蝠冬眠个体数量的变化
2.2 影响蝙蝠冬眠场所选择的主要环境因子分析
2017—2020年期间均有蝙蝠冬眠的隧道共有42个,各隧道冬眠蝙蝠数量是年际变化的。每年马铁菊头蝠和华南水鼠耳蝠在这42个隧道中几乎均有分布,而63.1%—78.7%的小菊头蝠冬眠于C58隧道,其余个体主要分布在C58后的隧道。通过多元线性回归分析发现(表1),隧道平均高度、水位、风速、声强、水温、环境温度等因子与冬眠数量相关性均不显著(P均> 0.05),隧道长度与冬眠数量呈显著正相关(P=0.001),经逐步回归分析获得的最佳模型仅保留了隧道长度这一个环境因子(Fst=11.76, AdjustedR2=0.208,P=0.001),表明隧道长度是影响蝙蝠冬眠场所选择的主要环境因子。

表1 环境因子对冬眠蝙蝠数量影响的多元线性回归分析
此外,有些隧道的固定位置,每年有比较稳定的蝠群冬眠,比如C41隧道距洞口约20 m处有一个马铁菊头蝠聚集群体,C58隧道距洞口100—110 m处有超过100只小菊头蝠独栖。
2.3 蝙蝠在隧道内分布特征及其冬眠方式
蝙蝠在各隧道分布存在空间差异性。在长度≤ 60 m的隧道中(占隧道总数的44%),冬眠个体数不及总数的5% (表2),超过95%的个体冬眠于长度> 60 m的隧道。至少有3/4的蝙蝠个体冬眠于长度超过200 m的隧道(占隧道总数的36%),特别是最长的C58隧道(长约770 m)拥有至少1/5的冬眠个体(历年最低197只)。此外,选择在人工砌石处以及距洞口≤ 30 m处进行冬眠的个体数量不及总数的20% (表2),包括约13.5%—22.9%的马铁菊头蝠、1.6%—4.5%的小菊头蝠、24.7%—36.8%的华南水鼠耳蝠个体;约4/5的个体选择在隧道深处冬眠。

表2 历年在短隧道(≤60 m)、人工砌石处以及距洞口≤ 30 m处冬眠蝙蝠的数量和比例
表3表明,随着距洞口距离的增加,环境温度逐步上升,相应的变异系数也逐步降低,相对湿度呈现相似变化。人工砌石处的温度和湿度及其变异系数与距洞口30 m处相当,这些区域易受外界环境影响。上述数据表明,多数个体选择相对温暖且稳定的隧道深处而非环境波动较大的地方作为冬眠栖点。

表3 距离洞口不同位置的温度和相对湿度及其变异系数
蝙蝠冬眠方式是多样的。绝大多数个体(> 90%)以独栖(solitary)方式进行冬眠,但也存在聚集现象(huddling),比如最大的一个马铁菊头蝠聚集群体包括了43只个体。不同物种冬眠的姿势并不相同,菊头蝠科蝙蝠一般垂直倒挂,独栖的马铁菊头蝠采用翼膜包裹身体,聚集的马铁菊头蝠则不包裹身体,而小菊头蝠一般不展开翼膜包裹身体;蝙蝠科蝙蝠一般是腹部贴壁倒挂或呈匍匐状卧栖姿势,不用翼膜包裹身体。菊头蝠一般选择较空旷的区域冬眠,而蝙蝠科蝙蝠则会选择石缝或凹窝处冬眠。
2.4 栖点温度与冬眠蝙蝠体温
栖点温度与蝙蝠体温之间呈显著正相关(R2= 0.98,P< 0.001) (图4),表明栖点温度直接影响了蝙蝠体温。二者的差异值主要分布在-0.5 ℃至0.2 ℃范围内(88.7%),大约64.1%的蝙蝠体温低于栖点温度。
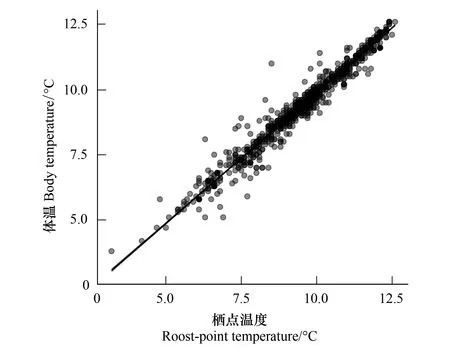
图4 冬眠蝙蝠体温与栖点温度之间的关系
距洞口≤ 30 m以及人工砌石处冬眠栖点温度((8.2±1.6) ℃,n=183),显著低于隧道深处栖点温度((9.6±1.3) ℃,n=671) (P< 0.001)。不同物种栖点温度有所不同,马铁菊头蝠((9.2±1.4) ℃,n=658)和华南水鼠耳蝠((8.8±1.1) ℃,n=60)之间无显著差异(P> 0.05),约9 ℃,但均显著低于小菊头蝠栖点温度((11.2±0.8) ℃,n=126) (P均< 0.001)。上述结果表明,蝙蝠冬眠所需的栖点温度不仅具有种内差异性,也具有种间差异性。
3 讨论
3.1 冬眠数量呈增长趋势
2017—2020年期间,河南省济源市邵原镇人工引水渠的隧道共有2科5属7种蝙蝠物种冬眠,其中马铁菊头蝠数量占绝对优势,且逐年增加,其他物种的数量则处于增、减波动状态。虽然调查时间有限,但一些长期的野外调查研究也发现了冬眠优势物种数量增加的现象[2, 16],这可能由以下原因造成:第一,优势物种更加适应当前的冬眠环境,且基数大,在一定程度上促进其种群的恢复和增长。第二,隧道内部寒冷黑暗,积水较深,人为干扰程度低,是较为安全和适宜的冬眠地,这可能促进蝙蝠种群繁衍和增长,如瑞典南部的废弃矿洞冬眠蝙蝠数量,在安装防护门后出现了增长和稳定[17]。然而,由于本次调查区域有限,存在临近区域的冬眠地(如废弃矿洞)不再适合冬眠时,蝙蝠可能迁徙到隧道内进行冬眠,从而造成数量增加的情况,但这有待进一步调查。
虽然冬眠蝙蝠数量呈逐年增长趋势,数量已超过了1000只,但仍明显少于2005年2月的数量[12]。华中北部地区的一些极端寒冷事件带来的地表热变化(如2007年为冰冻日数高值年[18],2009年为极端降温事件多发年[19]),可通过基岩的热传导引起冬眠场所低频热变化[20],进而增加蝙蝠越冬风险,这可能是引水渠隧道冬眠蝙蝠数量锐减的一个主要原因。但由于缺乏2006—2016年期间冬眠蝙蝠状况、隧道环境温度等相关数据,尚无法将数量锐减与极端气候变化建立关联。
3.2 隧道长度可能是蝙蝠选择冬眠场所的主要影响因素
在外界气候几乎一致的背景下,引水渠为蝙蝠提供了长短不一的77个冬眠隧道,但仅有42个隧道每年均有蝙蝠冬眠,且各隧道冬眠蝙蝠数量在4年间是不同的,因而隧道的微生境环境差异可能直接影响蝙蝠冬眠场所的选择。一般认为,微气候是决定蝙蝠选择冬眠场所的主要原因[5,21]。然而,当考虑到栖息地内部其他环境因子时,微气候的影响力会降低。例如,荷兰“New Dutch Waterline”防御地堡的内部空间大小和隐蔽空间的数量对冬眠蝙蝠数量的影响力比环境温湿度更大[7]。
在影响蝙蝠冬眠隧道选择的环境因子中,仅有隧道长度与冬眠蝙蝠数量显著正相关。隧道越长,内部微气候相对稳定的空间越大,能有效减少蝙蝠种内、种间的竞争,容纳更多的蝙蝠冬眠;同时隧道越长,其温度梯度越大(如C41,温度范围为3.4—10.0 ℃),能够使蝙蝠依据自身生理状态,在同一隧道内选择适宜温度的区域进行冬眠[22],进而降低因更换冬眠场所导致的能量过多消耗及被捕食风险。蝙蝠一般会选择较高的地方作为栖点,可有效避免天敌,而引水渠隧道相对安全,因而决定垂直空间大小的隧道高度和水深,可能不是影响冬眠场所选择的主要因素。风可以引起隧道内部温湿度波动,然而蝙蝠可以选择微气候相对稳定的石缝和凹窝冬眠[23],以减少风带来的不利影响。蝙蝠长期在流水环境中冬眠,各隧道的背景声强差异对冬眠的影响作用可能甚微。由于每个隧道内部存在一定的温度梯度,蝙蝠可根据自身生理状态选择更适宜的温度区域冬眠,因而隧道的平均水温和平均环境温度可能不会显著影响到隧道内冬眠蝙蝠数量。
3.3 不同的栖点位置和冬眠方式可能有利于冬眠成本优化
每个隧道内,蝙蝠冬眠栖点位置是不同的。大部分蝙蝠冬眠于相对温暖且稳定的隧道深处(距洞口>30 m),而并非集中在最冷的区域。在相对温暖环境下冬眠,可能会降低蝙蝠蛰伏持续时间,并增加能量消耗[24],似乎并不利于蝙蝠越冬。一些研究发现,脂肪储备多的蝙蝠个体更倾向于温暖环境冬眠,而脂肪储备少的个体则倾向冬眠于较冷区域[22, 25],这种策略既可保证越冬存活率,又可降低长时间蛰伏的代价(如增加生理和被捕食风险,减少交配机会等)[26]。考虑到栖点环境的热稳定性与冬眠哺乳动物的代谢支出成反比[27],在隧道深处温暖且稳定的环境中冬眠,蝙蝠的能量使用效率可能会更高[28]。
蝙蝠具有多样的冬眠方式。绝大多数蝙蝠是独栖的,这种方式能够降低被捕食和一些依赖于密度传播的疾病感染几率,比如白鼻综合症[29]。不同物种可能通过不同方法降低独栖冬眠个体的体热或体能损耗,比如马铁菊头蝠采用翼膜包裹身体,蝙蝠科物种则选择环境相对稳定的岩缝内冬眠。少量的蝙蝠采用聚集方式冬眠,该方式可减少暴露在外的体表面积,提高周边环境温度,降低体内热量损失和水分的蒸失[30]。遇到环境异常时,聚集群体任何个体的觉醒都可能促使其他个体被动觉醒,节约觉醒所需能量[31]。由此可见,不同的冬眠栖点和多样的冬眠方式,可能是蝙蝠对冬眠成本优化的策略。
3.4 栖点温度显著影响蝙蝠体温
要保持节能的蛰伏状态,蝙蝠需要将体温降至周边环境温度,以接近蛰伏代谢率(torpid metabolic rate)。因而,栖点温度往往显著影响冬眠蝙蝠体温。小菊头蝠栖点温度显著高于马铁菊头蝠和华南水鼠耳蝠,表明不同蝙蝠物种的栖点温度有所差别。这种栖点温度的种间差异性现象,可能普遍存在于冬眠蝙蝠中,比如湘西及武陵山地区冬眠的大菊头蝠((11.3±2.4) ℃,n=14)、中华菊头蝠((12.7±1.0) ℃,n=11)、西南鼠耳蝠((10.4±2.4) ℃,n=49)等具有不同的栖点温度[10, 32]。这种现象可能是由以下原因造成:第一,不同物种的体型差异较大,其蛰伏代谢率可能存在种间差异[33],从而造成物种的蛰伏适宜温度范围有所不同。第二,不同地区的冬季气候差异,导致冬眠场所之间存在温度差异,进而影响到蝙蝠栖点温度,比如在美国德克萨斯州北部较冷洞穴中冬眠的Perimyotissubflavus和Myotisvelifer的栖点温度,要高于南部温暖洞穴冬眠的同种个体栖点温度[34]。
值得注意的是,多数蝙蝠体温是略低于栖点温度,这与一些已有的报道不一致[11, 15]。一方面,可能与冬眠状态有关系,比如当蝙蝠处于冬眠阵的深眠阶段,接近冬眠过程中最低体温,会略低于栖点温度[35]。另一方面,调查时间的不同,以及物种差异都可能影响到这个结果。
4 问题及展望
第一,调查时间仅限于每年的1月(冬眠中期),事实上,在不同的冬季时期,蝙蝠数量和冬眠栖点位置可能是动态变化的[22],因而相关结果可能具有时限性。第二,由于客观条件的限制(相关设备的缺乏、仪器供电困难等),用于分析的微生境环境参数数值不是动态的,可能与真实情况存在偏差。
未来工作可在上述调查的基础上,关注冬眠不同时期的蝙蝠数量和冬眠位置动态变化趋势,调查引水渠附近区域潜在的冬眠地及蝙蝠冬眠情况,还要排查当地蝙蝠种群受到威胁的因素,这是野生种群保护和冬眠栖息地管理的基本前提。
致谢:感谢袁树信、陈传毓、韩鹏举、刘询、李子昊、李奥强、李仲乐等在野外调查中给予的帮助。
