基于FORCCHN模型的中国典型森林生态系统碳通量模拟
2022-04-25吕富成马建勇延晓冬
吕富成, 马建勇, 曹 云, 延晓冬,*
1 北京师范大学地理科学学部地表过程与资源生态国家重点实验室,北京 100875
2 华中农业大学植物科学技术学院,武汉 430070
3 国家气象中心,北京 100081
森林作为陆地上面积最大、生产力最高、结构和功能最为复杂的生态系统,在调节全球碳平衡、吸收温室气体等方面具有重要作用[1]。据测算,森林生态系统植被碳库(包括地上和地下部分)约为363 Pg C,占全球陆地植被碳库的70%;总初级生产力约为 90 Pg C/a,超过全球陆地生态系统的一半[2—3];全球森林生态系统固碳量约为2.4 Pg C/a,其固碳能力高于海洋和其他生态系统[4]。中国森林生态系统是陆地生态圈的巨大碳汇,尤其是西南林区和东北林区在固碳中扮演了重要角色[5]。因此,精确量化森林生态系统碳通量,有助于理解陆地生态系统碳循环对全球变化的响应与适应,寻求最合理的碳中和措施,具有重要的科学意义[6]。
森林生态系统碳循环模型是研究和预测森林碳通量及其循环过程机制的有效工具[7—8]。按照研究的空间尺度,森林生态系统碳循环模型可以分为斑块尺度和区域尺度的碳循环模型[9—11],以往对中国森林生态系统碳收支研究主要集中在区域尺度碳循环模型[12—15],且不同模型验证工作比较简单粗糙,缺乏不同时间尺度碳通量的验证工作。以个体为基础的斑块模型模拟所有时间内每株树木的出生、生长和死亡的全过程,每株树木的生长是通过对光照、水分及土壤的养分竞争获得,模拟的森林动态演替过程及其碳收支变化相对合理。FORCCHN模型( FORest ecosystem Carbon budget model for CHiNa,FORCCHN)是一个由我国自主研发的,基于个体生长过程和气象资料驱动的斑块模型[16],具有4个主要特征:(1)有机结合了碳、氮、水在大气—植物—土壤不同界面的循环与转化过程;(2)森林生态系统碳收支的驱动因素并非由当前气候和生态系统简单的统计关系预先确定,而是基于个体生理生态过程机制耦合;(3)森林的碳通量是由林分个体生长来确定的,进而使碳收支进行更为科学合理的模拟;(4)模型可用于预测未来气候变化下的森林生态系统碳收支的动态波动和未来平衡态。目前,该模型已在一些区域对森林碳收支进行模拟[17—19],但主要集中在单一森林生态系统和短时间序列的模拟,缺乏对不同类型森林生态系统长时间序列模拟验证。
本文旨在加深斑块模型对中国典型森林生态系统碳通量的模拟研究工作,通过对局部森林生态系统的模拟,检验模型在站点不同时间尺度的模拟性能,为研究大范围森林生态系统碳收支提供依据。以长白山温带阔叶红松林(CBS)、鼎湖山南亚热带常绿阔叶林(DHS)、千烟洲中亚热带常绿针叶林(QYZ)和西双版纳热带雨林(XSBN)为研究对象,着重评估模型对森林生态系统呼吸(Ecosystem Respiration: ER),总初级生产力(Gross Primary Productivity: GPP),净生态系统生产力(Net Ecosystem Productivity: NEP)的模拟效果,以期为未来气候变化下我国森林生态系统碳收支提供参考。
1 数据与方法
1.1 基本过程和模拟策略
模型由每日气象资料驱动,通过求和固定斑块面积上每株树木的碳收支,再耦合土壤碳收支,进而得到森林生态系统斑块面积上的碳收支,其涉及的主要特征如图1。
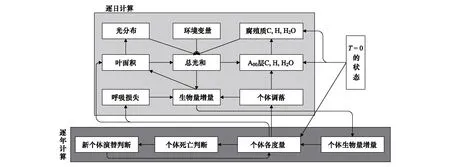
图1 模型的主要生态过程及时间步长
模型通过两种时间步长进行积分,对于每株树木而言,每天计算一次光合作用,维持呼吸,氮吸收和部分凋落物通量,土壤水,有机质的分解和氮的矿化都是以日为步长发生的。模型假定每株树木每日不同时间内维持呼吸,部分叶,细根凋落,土壤有机质分解和氮矿化速率恒定不变,从而避免在较短的步长上计算有关通量。模型所计算的树木个体碳氮的年净增加量,需要加上前一年储存的碳量,在每年最后一天分配到不同的树木成分内。当树木没有足够的碳用于光合作用产生叶片时,认为树木已经死亡并停止生长,从而树木的碳储存下降。死亡的树木所包含的碳氮含量,以及活的木质部分,花,果,叶和根的凋落物通量在每年最后一天加入到土壤枯枝落叶库中。由于本研究模拟时间较短,小于50年,因此假定树木不发生更新。
1.2 模型输入与输出
FORCCHN模型的输入参数主要包括气象数据、土壤数据、遥感数据及其它数据。模型所需每日气象数据包括:太阳总辐射(W/m2)、平均气温(℃)、最高气温(℃)、最低气温(℃)、降水量(mm)、空气相对湿度(%)、平均风速(m/s)、平均气压(hPa);土壤数据包括:土壤碳密度(g C/cm2)、土壤氮密度(g N/cm2)、田间持水量(cm)、萎蔫系数(cm)、土壤容重(g/cm3)、砂粒含量(%)、粉粒含量(%)、黏粒含量(%);遥感数据包括:不同森林植被类型分布图、模拟初始年的最大叶面积和最小叶面积指数(m2/m2);其它数据还有:站点的平均大气CO2浓度(μL/L),经纬度及海拔高度(m)。FORCCHN模型可输岀日尺度和年尺度的碳通量数据,主要参数包括:地上生物量(g C m-2d-1或 g C m-2a-1)、地下生物量(g C m-2d-1或g C m-2a-1)、总初级生产力(g C m-2d-1或g C m-2a-1)、植被地上呼吸(g C m-2d-1或g C m-2a-1)、植被地下呼吸(g C m-2d-1或g C m-2a-1)、土壤异养呼吸(g C m-2d-1或g C m-2a-1) 、凋落通量(g C m-2d-1或g C m-2a-1)、土壤碳库量(g C/m2)。
1.3 数据来源
涡度相关技术是测定大气与生态系统之间CO2交换,水分和能量通量最直接的方法,所提供的通量观测数据已广泛应用于陆地生态系统的碳循环研究[20]。本文使用的观测数据是利用涡度相关技术和基于ChinaFLUX 数据处理体系形成标准化的生态系统碳水通量和关键气象要素数据集[21],主要包括2003—2012年日尺度,月尺度,年尺度的气象数据和通量数据。通量数据主要包括生态系统呼吸(ER)和生态系统净CO2交换量(NEE)。生态系统总初级生产力(GPP)和生态系统净初级生产力(NEP)计算方法[22]如下:
NEE=-NEP=ER-GPP
模型所需的土壤数据、遥感数据及其它数据主要来源于中国通量数据网(http://www.cnern.org.cn)和IGBP-DIS发布的Global Gridded Surfaces of Selected Soil Characteristics数据集。通量站基本信息参见表1。

表1 4个通量站基本信息
1.4 分析方法
为评估模型对森林生态系统碳通量的模拟性能,本文分别采用模拟结果与观测值的相关系数(Corr),均方根误差(RMSE),以及偏差(Bias)等指标。具体计算公式如下:
(1)
(2)
(3)

2 结果
2.1 碳通量日尺度模拟
总体而言,模型较好的模拟了4种典型森林生态系统的逐日碳通量。图2和表2展示了以年为周期森林生态系统的逐日碳通量观测值与模拟值的相关系数,均方根误差及偏差。森林生态系统3个碳通量指标的相关系数从高到低依次为ER、GPP、NEP。4种森林生态系统ER的相关系数均超过0.85,且对应四分位距(IQR)最小,表明其模拟效果最好。不同森林生态系统GPP的相关系数略有差异:落叶阔叶林和常绿针叶林相关系数分别为0.86和0.73;常绿阔叶林和热带雨林相关系数在0.50左右。NEP相关系数较低,4种森林生态系统NEP的相关系数均小于0.5,从大到小其次为:落叶阔叶林,常绿阔叶林,常绿针叶林和热带雨林。
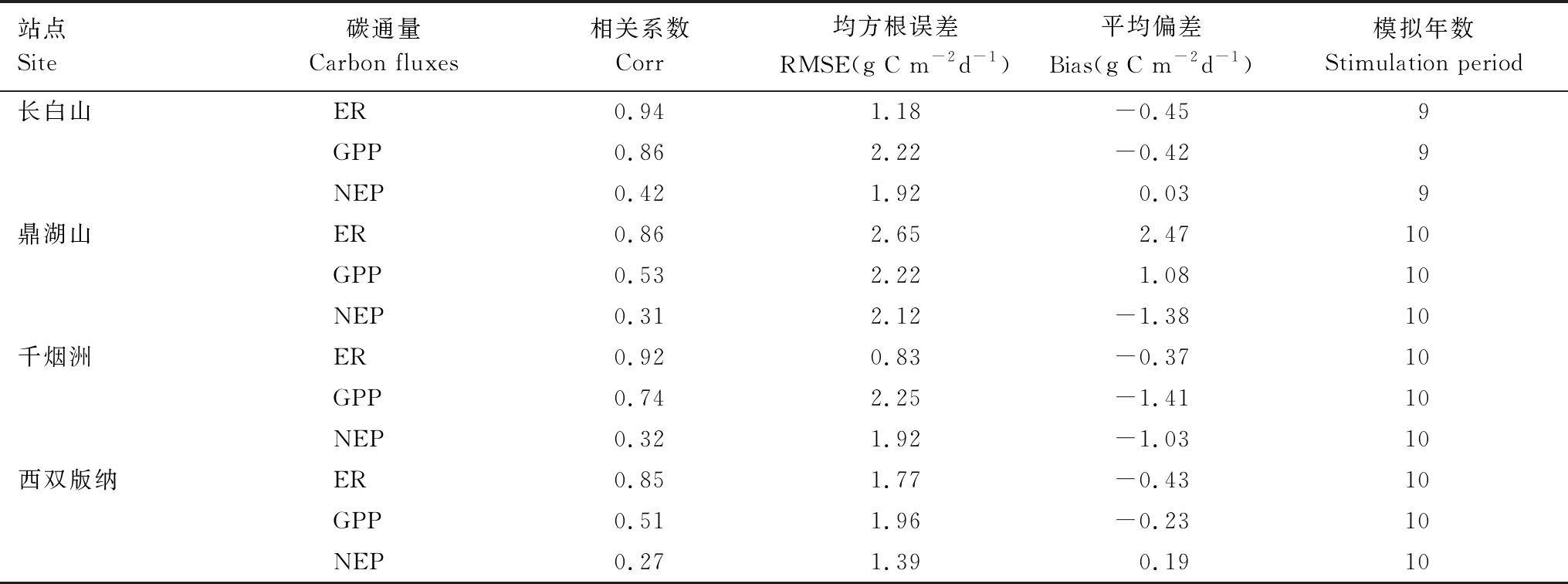
表2 不同森林生态系统碳通量以年为周期日尺度模拟结果
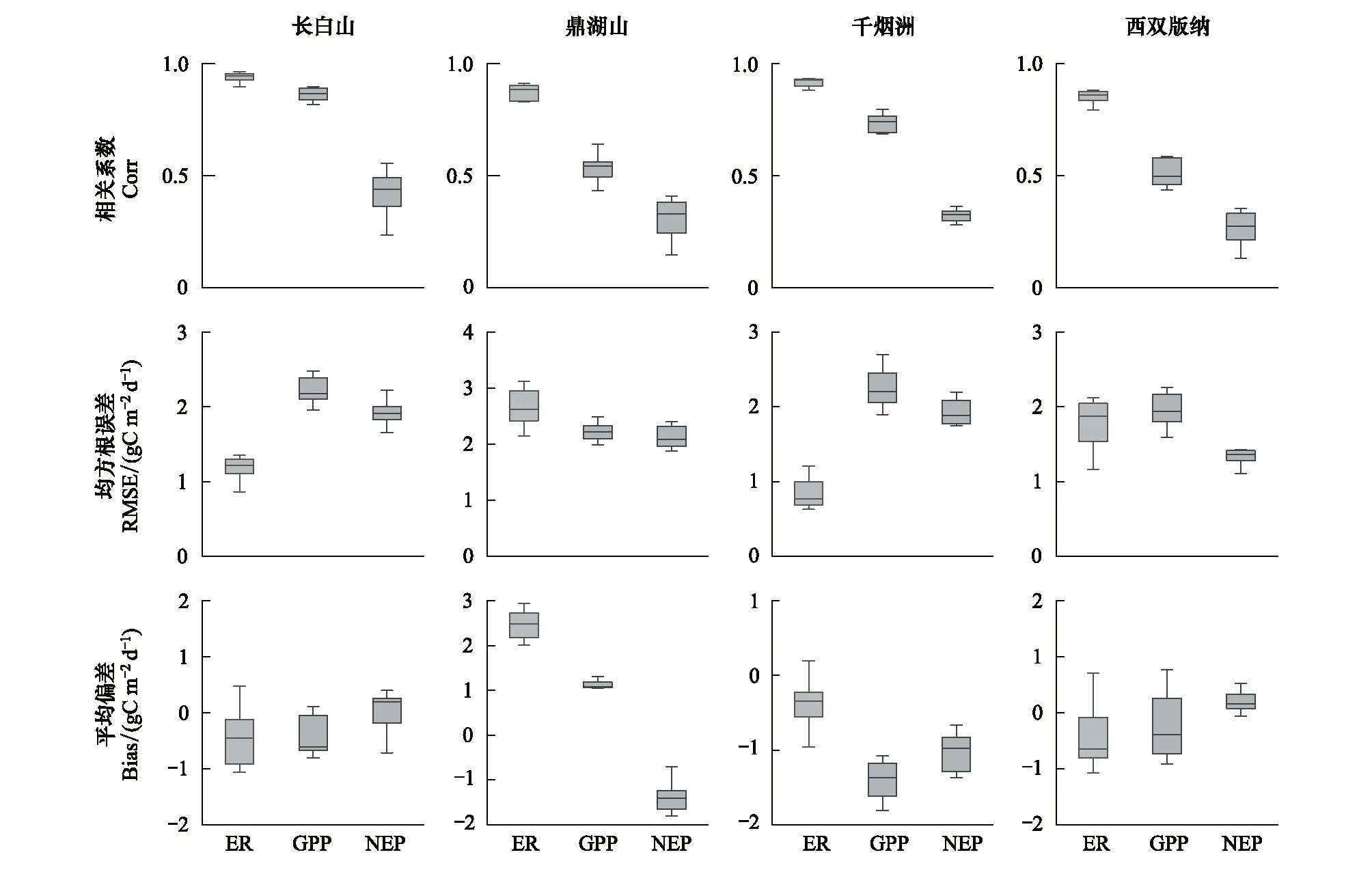
图2 每日碳通量模拟值与观测值的关系
不同森林生态系统均方根误差(RMSE)分析得出,3个碳通量的均方根误差GPP>NEP>ER。其中,常绿阔叶林的3个碳通量均方根误差最大(RMSE>2 g C m-2d-1),表明模型对该森林生态系统模拟误差较大,对其他3种森林生态系统碳通量模拟的均方根误差在1.67—1.77 g C m-2d-1之间,其中常绿针叶林ER均方根误差最小,RMSE=0.83 g C m-2d-1。
对偏差(Bias)分析发现,除常绿阔叶林外,其他3种森林生态系统ER和GPP偏差在-1.41—-0.23 g C m-2d-1之间,表明模拟值略低于观测值,常绿阔叶林ER和GPP偏差分别为2.47 g C m-2d-1和1.08 g C m-2d-1,模型高估了该生态系统碳通量。不同森林生态系统NEP偏差略有不同,落叶阔叶林和热带雨林的NEP偏差略大于0,模拟值基本与观测值持平;常绿阔叶林和常绿针叶林的NEP偏差均小于-1 g C m-2d-1,Bias分别为-1.38 g C m-2d-1和-1.03 g C m-2d-1,说明模型低估了NEP。
基于以上分析,可以得到以下简要结论:FORCCHN模型基本可以再现不同森林类型日尺度的碳通量,但大部分森林生态系统ER和GPP存在不同程度的低估,而高估了常绿阔叶林的ER和GPP。
2.2 碳通量季节变化模拟
总体而言,模型能够较好地模拟森林生态系统碳通量的季节动态。对不同森林生态系统逐月碳通量观测值和模拟值进行了相关分析(图3),结果表明,落叶阔叶林,常绿针叶林,常绿阔叶林和热带雨林ER,GPP和NEP的观测值和模拟值呈显著相关(P<0.01),ER观测值和模拟值的R2分别为0.93,0.91,0.84,0.77;GPP观测值和模拟值的R2分别为0.88,0.82,0.61,0.54;NEP观测值和模拟值R2最低,均小于0.4。碳通量观测值和模拟值对比发现,落叶阔叶林和常绿针叶林3个碳通量指标模拟值低于观测值,回归方程斜率略低于1;常绿阔叶林和热带雨林ER和GPP模拟值大于观测值,回归方程斜率大于1,两者的NEP模拟值小于观测值,回归方程斜率小于1。
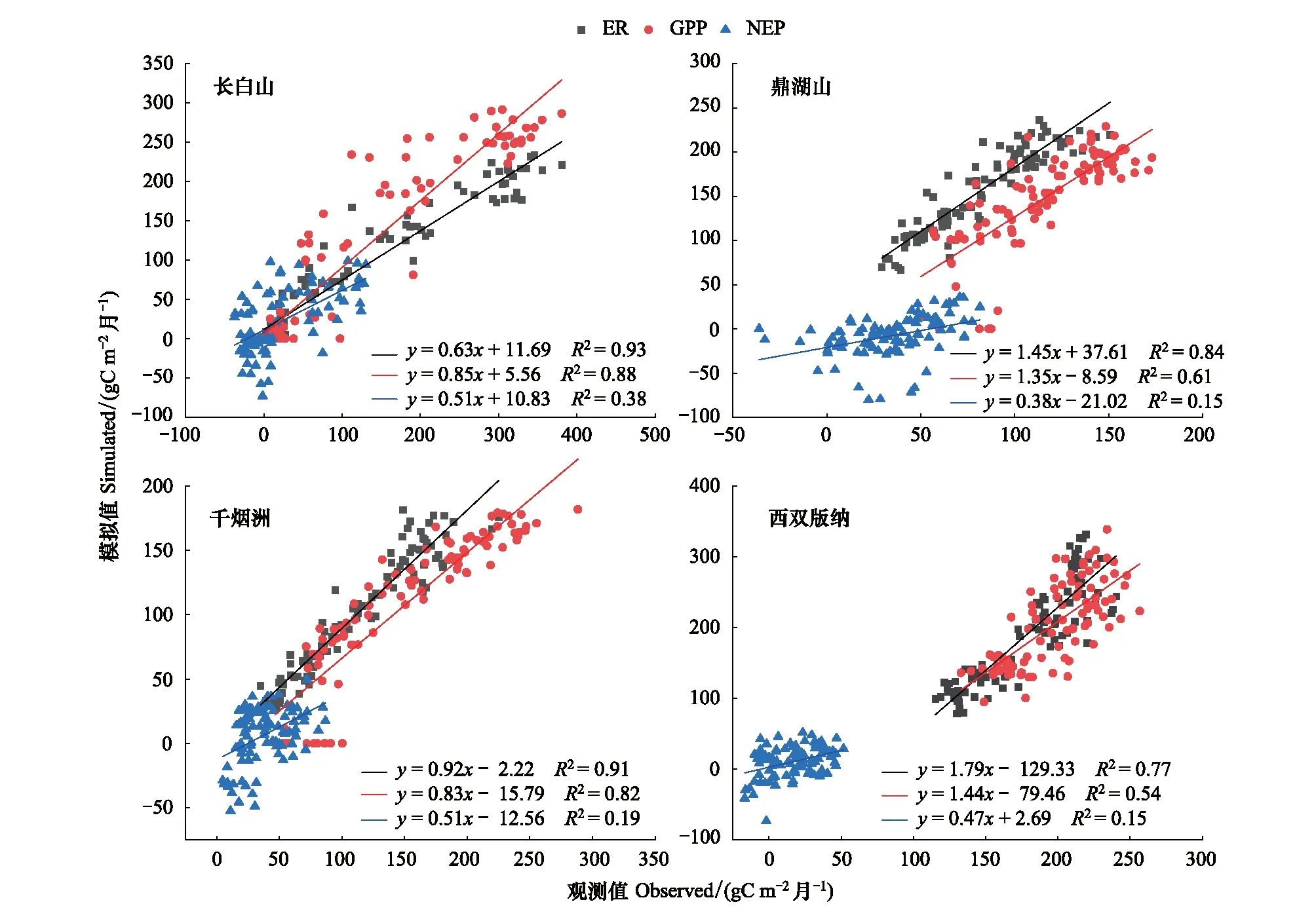
图3 每月碳通量观测值和模拟值的关系
为进一步探究FORCCHN模型对不同森林类型碳通量季节变化的模拟能力,图4给出了4种森林生态系统典型年份的ER,GPP及NEP的月变化。结果表明FORCCHN对不同森林生态系统碳通量的季节变化具备良好的模拟能力,能够再现中国森林生态系统碳通量由南向北递减、生长期高于非生长期的季节特征,但在少数月份模拟值与观测值存在偏差。
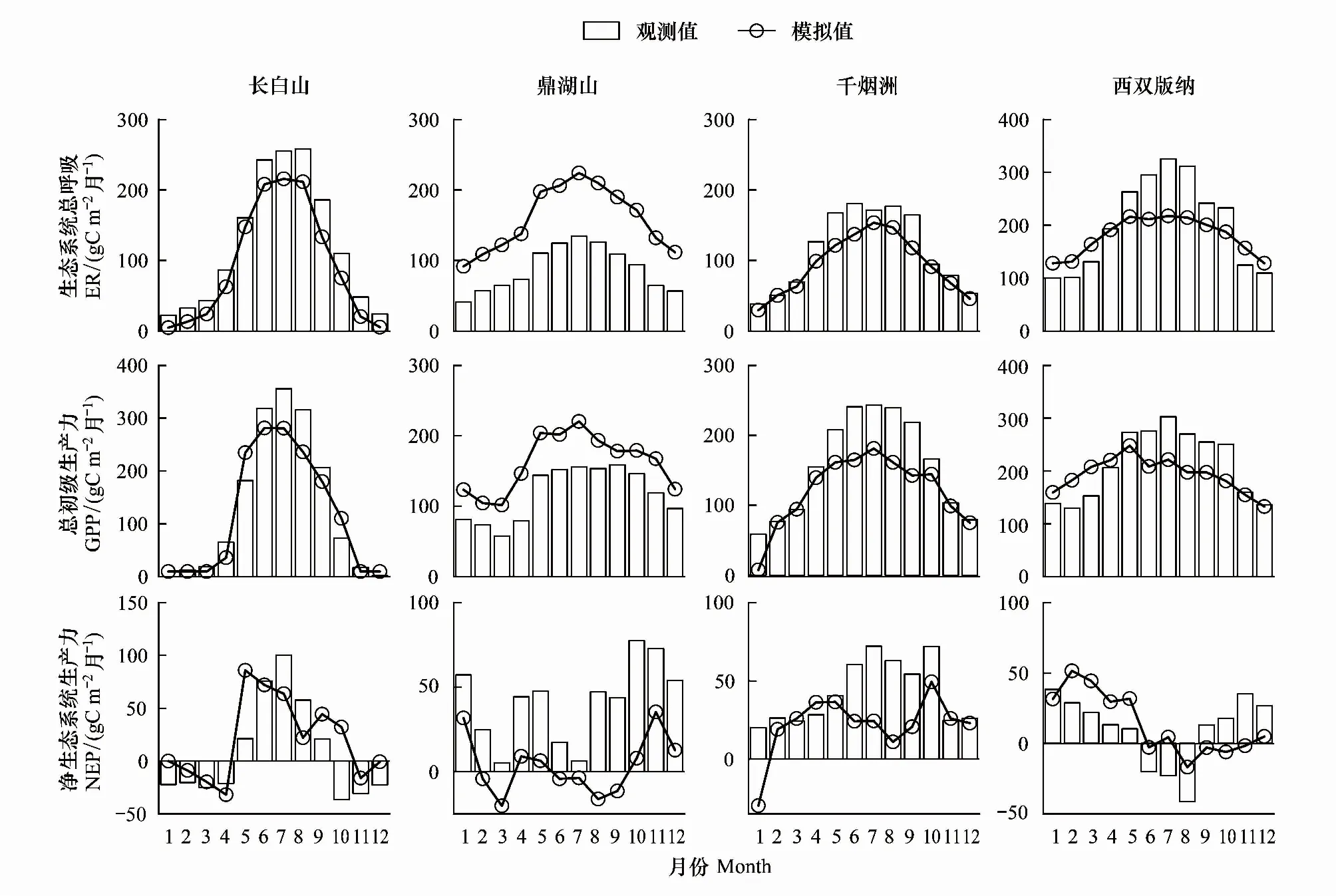
图4 碳通量观测值和模拟值的季节动态
在ER模拟方面,模拟与观测曲线波动非常相似,表明模型可以再现不同森林生态系统的季节变化。然而,模型对于4种森林生态系统ER模拟有一定的差异:模型对于落叶阔叶林,常绿针叶林和热带雨林的ER存在小幅度的低估,模拟值分别低于观测值13.43%,10.19%,6.83%;但模拟明显高估了常绿阔叶混交林的ER,模拟值高于观测值达90.06%。
同时,模型对不同森林生态系统的GPP和NEP均展现良好的季节模拟能力。落叶阔叶林生产期短,GPP季节差异显著,夏季6—8月GPP达到最高值,其他季节GPP小于100g C m-2月-1;随着年均温和降水的增加,常绿阔叶林和常绿针叶林的生长期较长,每年5—10月份GPP最大;热带雨林一年四季均为生长期,每月GPP均大于100g C m-2月-1。尤为可贵的是,模拟再现常绿阔叶林3月份GPP和NEP为一年最低的生长特征,与观测数据季节变化趋势高度一致。模型能够再现不同森林生态系统在一年四季碳源(NEP<0)、碳汇(NEP>0)的变化规律,特别是对于热带雨林,雨季(5—10月)为碳源,干季(11月到翌年3月)为碳汇[23],但遗憾的是,模型在数值上存在低估。
2.3 碳通量年尺度模拟
分析森林生态系统2003—2012年碳通量年际变化及偏差(图5—图7),结果表明,FORCCHN模型对不同森林生态系统碳通量模拟能力存在差异。在ER年通量模拟中(图5),落叶阔叶林、常绿针叶林、热带雨林模拟值与观测值的年通量差异较小,热带雨林观测ER年通量为2052.59—2629.68gC m-2a-1之间,模拟ER年通量为2062.20—2339.3 gC m-2a-1之间,年均偏差为-7.29%,仅在2009年和2010年模拟的ER通量高于观测值;常绿针叶林观测ER年通量为1212.79—1571.19 gC m-2a-1之间,模拟ER年通量为1122.90—1306.71 gC m-2a-1之间,两者呈显著相关(P<0.05),年均偏差为-10.11%,仅在2005年模拟值略高于观测值;落叶阔叶林观测ER年通量为842.99—1546.51 gC m-2a-1之间,模拟ER年通量为945.70—1146.01 gC m-2a-1之间,年均偏差率为-12.75%,仅在2005年模拟值高于观测值,同时落叶阔叶林ER年通量波动较大,模拟的年通量年际变化相对稳定。表明模型对3种森林生态系统ER年通量存在不同程度的低估,常绿阔叶林年均偏差率达93.87%,说明模拟值明显高于观测值。GPP年际动态模拟结果(图6)与ER模拟结果相对一致,即模型低估了热带雨林,常绿针叶林和落叶阔叶林GPP,平均偏差分别为-5.50%,-29.78%,-12.97%。主要原因在于夏季模拟的碳通量低于观测值,导致模拟的年通量低于观测值。同时高估了常绿阔叶林的GPP,平均偏差为25.71%。对比模拟和观测NEP发现(图7),落叶阔叶林观测年均NEP为252.51 gC m-2a-1,模拟年均NEP为224.85 gC m-2a-1,模拟值低于观测值10.95%;常绿阔叶林和常绿针叶林观测和模拟NEP年际波动趋势相对一致,但具体数值存在较大差异,模拟值均低于观测值。热带雨林观测和模拟NEP显著相关(P<0.05),观测年均NEP为91.70 gC m-2a-1,模拟年均NEP为128.93 gC m-2a-1,模拟值高于观测值40.60%。

图5 生态系统呼吸观测值和模拟值的年际变化及偏差

图6 总初级生产力观测值和模拟值的年际变化及偏差

图7 净生态系统生产力观测值和模拟值的年际变化及偏差
3 讨论
本文以长白山温带阔叶红松林、千烟洲亚热带常绿针叶林、鼎湖山亚热带常绿阔叶林和西双版纳热带雨林等4种中国典型森林生态系统为研究对象,利用涡度相关2003—2012年观测数据,评估FORCCHN模型对不同森林生态系统生态系统呼吸(ER),总初级生产力(GPP),净生态系统生产力(NEP)不同时间尺度的模拟效果。
不同时间尺度碳通量模拟效果对比,FORCCHN模型可以很好的捕捉森林生态系统碳通量逐日变化和季节性动态,但在每年和更长的时间尺度上表现一般,这与前人的研究结论一致[24—26]。已有研究表明,碳循环模型通常无法充分模拟观察到的落叶冠层物候的年际变化[27],并且落叶阔叶林的GPP和常绿阔叶林的ER春季的差异很大,同时GPP与NEP的年际变化存在协同性[28—29],这是导致碳通量模拟值和观测值年际变化一致性低的重要原因,也有人把年际模拟的不确定性归因于建模过程的偏差和碳分配机制的不明确[30]。为此,需要提高模型与物候的耦合程度和植被对热量的敏感性[27,31]。
从森林类型上看,FORCCHN模型在落叶阔叶林模拟效果好于常绿阔叶林,这个模拟结论得到了一些模型评估的支持[32—33]。落叶阔叶林具有明显的物候季节动态,模型可以通过明确不同时期植被生长的主导因素,模拟植被的生理生态过程,从而再现生态系统的碳通量动态。相反,常绿阔叶林受制于季节性叶片物候的细微变化,多种环境因素共同调控植物的光合作用,常绿阔叶林夏季的碳通量通常较高,并且存在较大的不确定性[31],这增加了模型模拟的难度。
ER在逐日尺度和季节尺度的精度高于GPP和NEP,关键在于模型改进了植物生长过程中呼吸量计算方法。生态系统呼吸由植被维持呼吸,植被生长呼吸和土壤异养呼吸3部分构成[34]。多数植物生长模型将维持呼吸与生物量建立线性关系,通过生物量去计算植物的维持呼吸量[35—36]。然而,有研究表明,对树木而言,呼吸作用主要用以维持边材软组织细胞的生长,这些细胞在所有活的树木内都存在[37],因此,在FORCCHN模型中,将维持呼吸表达为一个与边材量成正相关的函数,这有利于提高维持呼吸模拟精度。相对于ER和GPP,模型对森林生态系统NEP模拟误差相对偏大。有研究表明,ER和GPP的模拟误差可以影响NEP模拟的不确定性[38—40],因此,造成了NEP模拟精度低于ER和GPP。
模型对一些森林生态系统碳通量存在一定的偏差。原因可能在于:(1)模型参数不能完全解释个体树木的生理和生态特征,因为统一参数无法表征单株之间的生长差异[30]。比如,在12月至3月,落叶阔叶林模拟ER和GPP接近零,与观测结果存在一定差异。(2)ER和GPP受多种环境因素(比如氮沉降)影响,模型只能利用有限的参数和方程模拟植被生理生态过程,未考虑氮沉降对植物生长的影响。比如,Luyssaert 等[41]对欧洲森林研究发现,与不考虑氮沉降相比,考虑氮沉降影响的森林生产力平均增加11%。(3)空气温度作为驱动变量模拟不同类型森林生态系统呼吸精度存在差异,这可能是由树龄不同造成的。比如千烟洲人工林种植于1985年前后,其植物呼吸在生态系统呼吸中起着更为重要的作用,空气温度与生态系统呼吸相关性最强,因此本模型对其模拟效果较好。长白山落叶阔叶林为成熟林,树龄大约200年,异养呼吸可能是影响生态系统呼吸的重要成分,土壤温度作为驱动变量应该更为合理[42]。同时,由于受仪器故障和天气影响等原因,本研究中使用的常绿阔叶林碳通量数据存在部分误差,通量数据受随机误差的影响与通量的大小大致成正比[43—44],这可能是导致常绿阔叶林碳通量模拟与观测偏差较大的原因。
本研究基于观测数据,验证了FORCCHN模型在中国典型森林生态系统碳通量模拟的适用性,对研究碳循环具有重要意义。一方面模型可以利用气象、土壤和植被等历史数据,探究历史时期森林生态系统的碳收支状况,分析更大尺度生态系统的变化特征;另一方面模型可提供对于区域和全球尺度的森林生态系统时空连续分析,有利于对未来气候情景的模拟预测,为制定碳中和政策提供科学支撑。
4 结论
FORCCHN模型能够较好地模拟中国4种典型森林生态系统不同时间尺度的碳通量。模型对落叶阔叶林和常绿针叶林的ER和GPP逐日变化模拟效果较好,ER的相关系数分别为0.94和0.92,GPP的相关系数分别为0.86和0.74。模型对森林生态系统碳通量的季节动态具有良好的模拟能力,不同季节森林生态系统碳通量模拟值和观测值显著相关,ER模拟值和观测值的R2为0.77—0.93,GPP模拟值和观测值的R2为0.54—0.88,NEP模拟值和观测值的R2小于0.4,模型可以模拟森林不同季节碳汇、碳源变化规律。在年尺度模拟中,热带雨林、常绿针叶林、常绿阔叶林碳通量模拟值与观测值的年通量有很好的吻合度,但在绝对数值上存在差异。
致谢:感谢中国通量观测研究联盟(ChinaFLUX)在通量数据方面提供的帮助。
