北京东灵山土壤动物-植物互作关系对海拔格局的响应
2022-04-25丁彰琦徐国瑞张育新马克明
丁彰琦,徐国瑞,张 霜,张育新,马克明,*
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085
2 中国科学院大学,北京 100049
3 中国科学院西双版纳热带植物园热带森林生态学重点实验室,勐腊 666303
不同生物类群彼此并非孤立存在,而是往往互相依赖,形成特殊的互作网络系统,可地上-地下生物类群间的相互作用研究仍处于初始阶段[1]。与环境的过滤作用相似,种间相互作用在解释和预测生物多样性分布与探索群落形成机制的研究中也尤为重要[2—3]。在森林生态系统中,木本植物可以通过改变资源输入、林下光照强度以及土壤理化特性对草本和无脊椎动物群落产生一系列影响[4]。草本植物虽然仅占森林总生物量的0.2%,提供了4%的森林净初级生产和16%的叶凋落物[5],但其凋落叶营养成分较木本植物更高,分解速度更快[6],也同样影响着土壤动物群落的组成和结构。而土壤动物可以通过调控凋落物的分解(如腐食性)、摄食根系直接影响植物生长(如植食性),或通过改变土壤物理结构、养分可用性以及土壤细菌与真菌群落的组成和活性来间接作用于植物生长(如蚯蚓和线虫),对推进整个生态系统的养分循环至关重要[7—8]。地上-地下之间有着复杂的反馈和调节机制[9],不同种木本植物对林下物种作用的特异性,类群间的正关系、无关系甚至负关系均有过报道[10]。然而,大多数研究集中于特定类群(如植物和微生物的关系),对于土壤动物与植物类群之间的关系如何响应环境变化的研究还比较缺乏[11—12],限制了对生物多样性维持机制的全面评估[13]。
首先,环境梯度的参与让种间相互关系的分析变得非常复杂,类群之间相互作用的海拔格局目前就不清楚。相比其他的自然地理梯度,海拔在较短空间内展现出跨度很强的环境变化,为生物多样性对环境变化的响应研究提供了良好的平台[14]。因此,鉴于土壤动物对不同资源类型做出的适应性选择,以及由于自身扩散能力和对微生境响应机制的差异,原则上将导致相互作用强度随海拔梯度发生转变,影响其各类群的空间分布,以致群落结构的改变。因此,地上植物(木本和草本)和地下土壤动物的相互作用与联系如何响应海拔的改变引起了众多研究者的注意。针对α多样性的大量研究已表明,木本、草本植物和土壤动物有着多元化的海拔分布格局[15—16],越来越多的研究者将关注点又放在能够反映物种空间周转或嵌套的β多样性[17],但依旧难以体现多类群的系统性变化[18]。近年来,基于拓扑研究的网络分析已被证明是理解物种共现格局特征的有力工具[19]。其中,非随机共现模式在大型生物体中很常见[20],但最近对大型复杂土壤微生物群落数据集的研究中也同样发现了强非随机关联与生态位的特化,以及一些确定性过程[21]。因此,结合群落中各物种的α、β多样性、共现格局以及对各拓扑指数的变化,将有助于揭示种间关系与生态位分化,为潜在的群落组装规则提供新见解[22]。
其次,尽管木本与草本植物对土壤动物群落的影响都不可忽视,但二者的作用机制之间可能存在差异[23]。一方面,木本与草本植物群落自身的物种组成与结构对海拔的响应模式不同;另一方面,对土壤动物群落的资源输入模式也不同(如凋落物的适口性、频度与总量,以及根系分泌物)。将不同生活型地表植被与土壤动物之间互作关系的细化剖析将有助于对地上-地下耦合关系响应环境变化格局和机制的清楚认识与精准预测[24]。另外,在森林生态系统中,凋落物和土壤表层虽然相近,但二者为较小体型的土壤动物提供了截然不同的生境[25]。内部的孔隙大小会影响活动空间,造成扩散限制,营养物质的组成,水分、温度等其他非生物因子也存在差异。因此地上植物对两层中土壤动物的影响机制也可能不同:影响凋落物层的是输入资源的多度与多样性,而影响土壤层的则可能是植物的根系分泌物,对微生物群落或凋落物分解的间接调控。鉴于物种对逆境的耐受能力往往在不同的背景环境条件下会发生改变[26],将两层对比研究或许可以更全面地揭示土壤动物与地上植物的关系。
虽然近年来复杂网络在传粉、种子传播、昆虫-植物和菌根等互惠关系研究中应用较多[27],然而对于土壤动物-植物互作关系研究中的应用还较为缺乏。由此,本文以设立在东灵山森林生态系统中的海拔梯度样带为基础,通过对比不同生活型地上植被与凋落物层和土壤层中各取食功能群土壤动物,以期回答下列科学问题:沿海拔梯度,(1)各类群多样性变化是否存在差异?(2)地上植物与地下土壤动物种间相互作用是否发生改变?(3)相互作用关系在两层之间是否存在差异?
1 研究地区与方法
1.1 样地设置与样品收集
本研究区位于北京市西郊门头沟区,在东灵山北京森林生态系统定位研究站附近进行(115°26′—115°30′E,40°00′—40°02′N)。东灵山属典型的暖温带半湿润大陆性季风气候区,土壤属于山地棕壤,年平均降水量约550 mm(集中于7—8月),年平均气温5—10 ℃。山地森林带状植被高度异质性,主要有辽东栎林,混交林(如椴树、胡桃、白蜡等),桦树林(黑桦、白桦、棘皮桦),林下有次生灌木(如绣线菊属,胡枝子属等)。在研究区内选择十个坡度相近、并以暖温带落叶阔叶树种辽东栎(Quercuswutaishansea)为优势种的山体,再于各山体的西坡设置范围不等(80—180 m)、宽度相等(10 m)的样带,从而衔接形成1020—1770 m的海拔梯度。
然后,将此十条样带按照10 m × 10 m大小划分为119个基础调查样方。调查每个样方内的木本植物种类和数目,并沿每个样方的对角线设置3个1 m × 1 m的样点,调查样点内草本植物的种类和数目。在每个样方内随机选取3个0.6 m × 0.6 m的小样方,收集内部的凋落物(期间用手捡法采集大型土壤动物),然后用内径为8 cm的土钻随机采集6个10 cm深的土柱混合作为一个空间采样点采集土壤层中土壤动物的样品。将收集的凋落物和土壤样品装入布袋后,于实验室内采用改良的Berlese-Tullgren干漏斗法(烘48 h)进行中小型土壤动物的收集。分离结束后于显微镜下鉴定其种类并计数,除前气门亚目和中气门亚目外,其他类群均鉴定至科或形态种水平[25]。其中在凋落物层中以弹尾目、寄螨目和真螨目为主,分别占总多度的37.95%、22.47%和22.98%;土壤层主要有弹尾目、寄螨目、真螨目和双翅目,分别占总多度的19.99%、24.68%、34.5%和11.27%。所有野外工作都于2013年7月—8月间完成。
1.2 数据分析方法

与以往网络分析使用的数据集不同,本研究中土壤动物与植物样品之间并不存在原始的可观测到的相互对应关系。而目前大多数计算种间相互作用的方法却并没有充分地考虑间接作用,且将协方差系数作为相互作用也因为其成对相互作用从相关残差推断出来的,所得的效应值无法直接与协变量相对应。因此,本文首先使用马尔可夫随机场(MRF),通过控制间接相互作用,获得具有权重的类群间直接相互作用数值矩阵。期间,利用记录的GPS坐标,建立了平滑高斯过程空间回归来去除样方间的空间自相关[30]。然后,用bipartite包对获得的3个海拔段(低-中-高段分别含有35、38、46个样方)所对应的植物-土壤动物二分网络矩阵结构进行了拓扑分析[31],本文选取的网络结构评价指标有:连接度和嵌套性(NODF),土壤动物的生态位重叠度(文中采用“horn”距离)、灭绝曲线系数和普遍性。
鉴于各网络中土壤动物与植物各自的物种数量和组成都发生了变化,为了在不同海拔段的网络结构之间进行对比,本研究采用建立零模型的方法获得观测值对应的零模型的标准化效应值(SES=(Xobs-μnull)/σnull)。其中,虽然可以利用马尔可夫随机场计算得到土壤动物与植物之间的有权无向型网络矩阵,但由于是通过统计间接获得的,因此为了使所得结果更加保守,本文将原始矩阵去除权重而转化为0—1型二元数值矩阵后,选择“r00”的零模型算法[32]。通过计算1000个随机网络矩阵所对应的参数,并根据观测值在零模型结果中的分布得到对应的P值(取95%的置信区间)。
上述分析都在R- 3.5.3中完成。
2 结果与分析
2.1 土壤动物和植物的多样性海拔格局
凋落物层和土壤层的土壤动物群落在低-中-高三个海拔段的物种累积曲线(图1)表明,样品收集充分,能够较为全面地反映两层中土壤动物在三个海拔段的多样性。而且在采样点数量相同时,土壤动物丰富度在中海拔段较高。
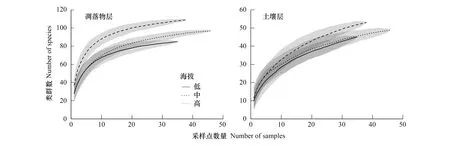
图1 三个海拔段土壤动物物种累积曲线
图2表明,凋落物层中土壤动物的稀有类群在中海拔段多样性最高(如q=-2时,低115.7,中192.6,高126.7),优势类群在高海拔段多样性最低(如q=2时,低8.0,中7.6,高6.6);而土壤层中土壤动物的稀有类群在中海拔段多样性较高(如q=-2时,低22.4,中31.6,高26.6)。木本植物无论是稀有还是优势类群,都是在低海拔段多样性最高。而草本植物却无论是稀有还是优势类群,都是在高海拔段多样性最高。
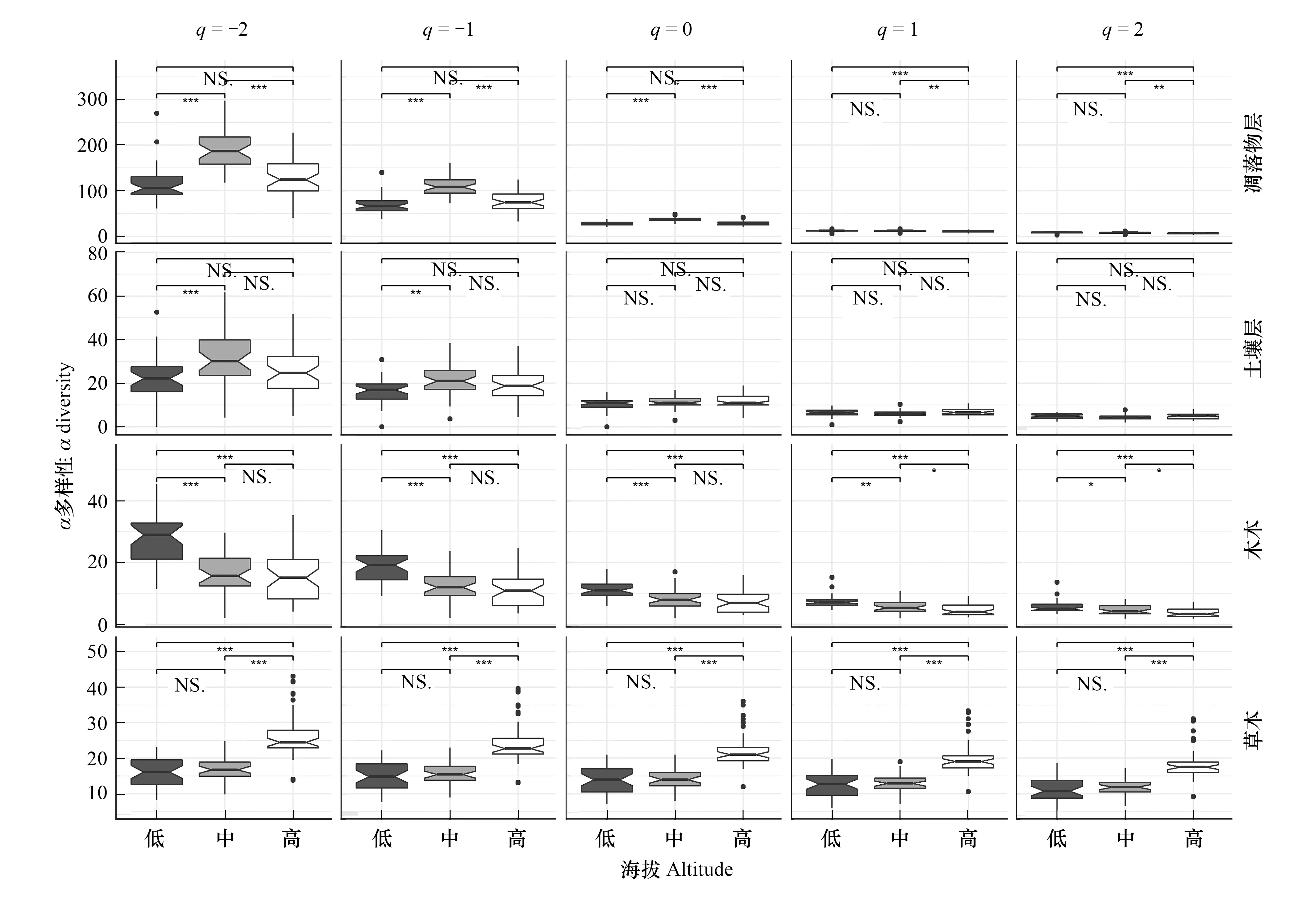
图2 各生物类群在三个海拔段的α多样性
在海拔梯度上,4个生物类群的β多样性都随海拔距离线性增加(除q=-2时土壤层中的稀有类群),即各类群(无论稀有还是优势类群)的组成差异都随海拔距离而增加,且其中稀有类群的β多样性截距要高于优势类群(相邻样方间的物种组成差异较大)。而且从图3中可以明显看出,与土壤动物和木本植物不同,随海拔距离的增加,草本植物的优势类群物种组成差异的变化速率要更大(图中斜率较大)。
2.2 土壤动物-植物互作网络分析
为了确定森林生态系统中的土壤和植物(木本-草本)类群对海拔的响应模式,本研究通过结合三个海拔梯度3种生物类群构建了互作网络,通过分别与建立的零模型对比发现,土壤动物-植物间的互作网络基本都表现出了显著的非随机关系格局,且两层中的土壤动物在各自网络中的连接度累积分布均符合幂律分布(拟合系数的P<0.05)。
图4表明,除草本与土壤层中土壤动物互作网络,其他三类网络的连接度都是在中海拔段最高,其中木本植物参与网络的连接度都是中海拔段>低海拔>高海拔段。网络的嵌套性指数NODF在木本植物与土壤动物(凋落物和土壤层)互作网络的中海拔段最高,尤其是与土壤层土壤动物的互作网络中明显高于低-高海拔段(低6.0,中11.1,高3.1);而在草本植物与土壤动物(凋落物和土壤层)互作网络中的嵌套性指数NODF在高海拔部分最大,尤其是和凋落物层中土壤动物的互作网络显著高于零模型预测值,且嵌套性指数随海拔升高而增大(低14.1,中17.9,高32.7)。

图4 土壤动物-植物互作网络的标准化拓扑特征比较
两层中土壤动物的生态位重叠度(niche overlap)均显著高于零模型的模拟值,其中凋落物层中土壤动物与草本植物、土壤层中土壤动物与木本植物互作网络都是在中海拔段较高,而凋落物层中土壤动物与木本植物、土壤层中土壤动物与草本植物互作网络都是在高海拔段较低。而在四类土壤动物-植物的网络中,加权后土壤动物的连接普遍性(generality)都随海拔升高而变大,且都高于零模型的模拟值。另外,本研究还利用灭绝曲线系数(extinction slop)作为评估群落稳定性的参数,结果表明,凋落物层土壤动物与植物(木本和草本植物)网络的灭绝曲线系数都是负值,且其绝对值大小趋势一致,都是在中海拔段最小。而土壤层中土壤动物与木本植物网络的灭绝曲线系数在低-中海拔段都为正值且相近,但在高海拔段相对于零模型并不显著;与草本植物网络的系数的绝对值则随海拔增大。
3 讨论
物种间相互作用是群落空间分布格局与构建机制的重要影响因子。本研究对比分析了东灵山地区地下土壤动物(按照生境分为凋落物层和土壤层)与地上植物(按照生活型分为木本和草本)互作网络的结构参数和各生物类群的多样性海拔分布,将有助于深入了解土壤动物的空间聚集机制及其多样性海拔分布机制。
3.1 多样性的海拔变化
沿海拔梯度,植物和土壤动物等各生物类群不仅物种组成会发生变化,各物种的数量也同样会存在差异。因此,对各类群生物多样性的海拔分布格局的了解是研究地上-地下生物之间的相互作用关系的基础,也是对预测生物间相互作用关系对未来环境变化的必要前提[1,19]。本研究表明,无论是稀有类群还是优势类群,相比土壤层,凋落物层中土壤动物有着较高多样性,且两层中土壤动物的优势类群随海拔梯度有着不同的变化格局。但无论木本还是草本植物,其稀有和优势类群的海拔变化模式基本相同。其中木本植物多样性随海拔降低,而草本植物多样性变化则恰好相反。另外,本研究发现,植物和土壤动物中各类群在海拔梯度上存在显著的聚集性分布,且稀有类群的格局特征比优势类群更大。其中,草本植物的物种组成对海拔梯度的响应模式与其他生物类群存在差异,其优势类群随海拔距离的变化速率也很高,聚集性分布格局最明显。本文的研究区域从低海拔段一直延伸至树线附近,因此受温度、光照和水分等因子的综合作用,形成林下草本植物到亚高山草甸群落的转变[33]。而对于木本植物,由于本研究样带一直都是以辽东栎为优势种,其优势类群的组成变化必然不会很大。这或许也是两层中土壤动物优势类群差异为什么不大的原因,鉴于其受资源多样性的影响可能较小[34—35],将来可以结合其他环境因子随海拔梯度的变化情况,深入探讨土壤动物群落多样性海拔格局的驱动机制。
3.2 网络的海拔变化
已有大量文献通过网络分析来探讨生态系统结构和功能,对于了解生物间互作关系在群落空间分布格局的作用以及多样性的维持机制具有重要的作用[19,36—37]。本研究结果表明,土壤动物(凋落物层与土壤层)与植物(木本和草本植物)在三个海拔段上互作网络的各项拓扑参数都与建立的零模型存在显著差异。这也表明,在土壤动物海拔分布与群落的装配过程中,除外界环境因子的驱动外,生物间相互作用也同样产生着重要影响。
由于选择性灭绝、生境异质性差异、扩散能力以及物种空间分布的幂律特性,自然界中许多互惠网络都存在一定的嵌套特性,这也一直是群落生态学的关注热点,对认识群落组成和结构层次以及生物多样性的了解具有重要价值[38—39],在网络分析中也经常通过嵌套性NODF来表征物种相互关系组合中的的冗余特征[40]。本研究发现,木本植物-土壤动物互作网络在中海拔部分的冗余度高,而随着海拔的升高,草本植物-土壤动物网络在高海拔段有着较高的嵌套性指数。木本植物-土壤动物网络的连接度和嵌套性可能主要与土壤动物的多样性有关,而草本植物-土壤动物网络的连接度和嵌套性则可能与草本植物的多样性有关[23]。鉴于木本植物多样性随海拔梯度的下降和草本植物多样性的升高,这也可以说明,为了能够抵抗海拔升高带来的环境胁迫,两层中的土壤动物与植物的互作关系都在木本和草本植物之间发生了转变,与草本植物建立了更多的联系,为从森林生态系统过渡到草甸而做出了适应性调整。
本研究还关注了用来衡量物种对相同资源的利用程度的生态位重叠度,用来展示类群之间相互作用方式的相似程度,从侧面反映各类群间竞争的强度。在凋落物层土壤动物-草本植物网络和土壤层土壤动物-木本植物网络中,土壤动物生态位重叠度在中海拔段较高,说明在中海拔部分土壤动物类群的竞争强度高,生态位分化程度较低。在四类网络中土壤动物的生态位重叠度都是在高海拔段较低,说明当外部环境胁迫较强时,土壤动物类群间的竞争强度减弱[41]。而且在四类网络中土壤动物的连接普遍性都随海拔而增大,也从侧面说明了,相比低-中海拔段,在高海拔段土壤动物与植物的互作关系变得更密切。
沿海拔梯度,凋落物层中土壤动物-植物网络的稳定性(灭绝曲线系数)SES值均为负值,且根据其变化趋势推断可能主要与土壤动物的稀有类群有关,而土壤中土壤动物-植物网络的稳定性则可能与植物的多样性有关。土壤层土壤动物-木本植物网络在低-中海拔段的灭绝曲线系数较大,表明随着木本植物的周转和消失,土壤动物还可以选择其他木本建立新的联系。而与草本植物组成的网络则不同,主要是在高海拔段有着较高的稳定性。结合植物的α多样性格局可推断,在高海拔段,土壤动物主要是扩大了与各类草本植物的联系,可能也同样增强了草本植物对环境胁迫的耐受性[42—43]。
综合以上结果,凋落物层与土壤层中的土壤动物类群与地表植物可能有着不同的相互作用机制,而且不同生活型植物对地下土壤动物的影响也不同,不同类型土壤动物-植物互作网络随海拔梯度的变化存在差异。
3.3 研究分析与展望
土壤动物是生态系统中重要的组成因子,推动着营养元素的循环和能量的流动。然而,尽管地上-地下生物的多样性以及二者的相互作用一直都是群落生态学研究的热点[9,12,44],但由于其较小的体型和复杂的生境异质性,其与植物的互作关系研究依然较为缺乏,通过网络分析来系统地反映土壤动物与其他类群的相互作用关系是目前研究的趋势[45]。与其他的生物网络,如昆虫-植物的传粉、动物-植物的种子传播、菌根-植物互作网络等研究相比[36,46],土壤动物-植物相互作用关系的研究目前还较为落后[47]。一部分原因是由于样品收集和鉴定中的不足,现在可利用高通量测序技术的使用,利用分子生物学手段,得到分类更加精细并且功能类型更多样化的土壤动物的类群。其次,与其他类型的互作网络分析相比,本研究是基于同一样方中植物与土壤动物的共现格局[48]。因此,对相互关系的鉴定推断方式主要依赖于统计分析,而不是基于真实观察到的两个生物类群之间的对应互作关系。虽然可能会揭示一些潜在的无法直接观测到的关系,但同时也可能会包括一些关系并不紧密的类群,对结果造成较大的干扰。因此,从实验设计的角度来看,直接的野外调查可能存在一定的缺陷,与控制实验和部分人为干预结合的方式可能对揭示真实的土壤动物-植物相互作用关系更为有效。例如,土壤动物主要与植物根系或其凋落物存在直接和间接的互作关系[49—50],如果能集中对根际土和植物附近的凋落物中进行样品收集,将可能更有助于对植物-土壤动物间互作网络的分析。
4 结论
地上植物一方面可以通过生物性作用,如根、茎、叶、分泌物与凋落物;或者化学性,如植物的化感作用;以及物理性,巢穴、遮蔽造成的微生境对地下土壤动物多样性造成直接或间接的影响。因此,地上-地下两个系统间的耦合关系如何对环境变化做出动态响应也一直是生态学研究的重点。本研究对比分析了北京东灵山森林生态系统中木本和草本植物与凋落物与土壤层内土壤动物的互作网络关系变化,同时展示了各类群多样性沿海拔梯度的变化格局。结果表明,沿海拔梯度,(1)土壤动物、植物各类群的多样性变化存在差异:土壤动物的稀有类群在中海拔段多样性较高,木本植物多样性在低海拔段较高,而草本植物多样性则在高海拔段较高,且土壤动物与木本植物各自的优势和稀有类群的多样性海拔变化也不相同;(2)土壤动物、植物各类群的群落组成差异都随海拔距离而增大;(3)地上植物与地下土壤动物之间的互作关系随海拔发生改变,且在两层之间、木本和土壤动物与草本和土壤动物的互作关系存在明显的差异。
