自噬对噪声性耳聋急性损伤期毛细胞凋亡的作用机制研究*
2022-04-24于佳雪黄宏明盛晓丽
李 林,于佳雪,黄宏明,崔 勇,盛晓丽△
(1.广东省人民医院珠海医院/广东省珠海市金湾中心医院耳鼻喉头颈外科 519040;2.广东省医学科学院/广东省人民医院耳鼻咽喉头颈外科 510080)
噪声性耳聋是感音神经性耳聋里最常见的原因之一,目前其发病机制及治疗是全世界共同难题之一。导致噪声性耳聋的一个关键因素是感觉毛细胞的氧化损伤。研究表明活性氧(ROS)的过度产生已成为各种内耳损伤的常见病理机制,如噪音暴露和耳毒性药物所致的耳蜗损伤[1-2]。既往研究发现在噪声暴露前后给予抗氧化剂治疗能够有效减少噪声引起的毛细胞死亡从而有效改善噪声性耳聋的发生[3-4]。越来越多的证据表明ROS会通过诱导自噬发生来保护细胞,这一过程是通过形成自噬小体来介导的,自噬小体与溶酶体融合,对吞噬的成分进行酶解[4]。通过程序性地处理这些可能受损的细胞成分,包括受损的细胞器和错误折叠的蛋白,起到了限制病理进展的作用[4-5]。
自噬与ROS表达水平的复杂关系表明自噬在噪声性耳聋的发生发展中起着潜在关键作用,然而其具体的调控机制仍不明确。越来越多的证据表明自噬诱导的氧化应激(OS)与耳聋密切相关[6-8]。脂质过氧化、线粒体DNA氧化损伤和谷胱甘肽结合蛋白的过表达都被证明与小鼠耳蜗老化密切相关[6,9-10]。因此,探寻自噬导致的耳蜗细胞变性的确切机制及可能导致耳聋的关键分子靶标,无疑对噪声性耳聋患者的治疗有着极为重要的意义。本研究拟观察自噬在噪声性耳聋中的确切表达情况,以及其对ROS水平的调控机制,进一步明确自噬和ROS与听觉阈值之间的相关性,为临床治疗噪声性耳聋奠定理论基础。
1 材料与方法
1.1 实验动物及主要试剂、仪器
健康成年 SD 大鼠 48只,雌雄不限,体重220~240 g,由华南理工大学动物实验中心提供,分为3组:对照组,自噬抑制剂3-甲基嘌呤(3MA)组和自噬激动剂雷帕霉素(RAP)组。RAP、3MA购自上海宝曼生物科技有限公司;TUNEL 试剂盒购自瑞士Roche 公司;酶标仪购自美国Thermo Fisher公司;LE2202S 型电子天平购自德国Sartorius 公司;Image-Pro Plus 6.0 图像分析软件(美国Media Cybernetics 公司);LeicaDC500 病理图像分析系统(德国Leica 公司);H-7650 透射电镜(日本Hitachi 公司)。
1.2 噪声暴露
噪声性耳聋模型按既往文献进行造模[11],即将大鼠放入暴露舱(0.5 m×1.5 m×0.5 m),用UZ-3型噪声发生器发声,平均声强为106 dB。动物暴露范围内声扬不均匀度为±2 dB,噪声频率为4 kHz,持续暴露6 h。
1.3 腹腔注射给药
RAP以75 mg/mL溶于DMSO并在-20 ℃下储存;3-MA以30 mg/mL溶于0.9%的生理盐水并在-20 ℃下储存。根据文献报道使用剂量[11],即每只大鼠共接受3次腹腔内注射,RAP的剂量为每次7.5 mg/kg,3-MA的剂量为每次30 mg/kg。使用时间为在噪声暴露前24 h、前2 h和暴露后即刻进行腹腔注射。
1.4 听觉脑干反应(auditory brainstem response,ABR)
按以往文献描述对大鼠行ABR检测[11]。即分别以10%水合氯醛(30 mg/kg)腹腔麻醉,在静电屏蔽室内采用Nicolet Compass系统测试ABR。ABR阈值即为观察到有刺激反应的最低刺激水平和无刺激反应的最高水平。所有的ABR阈值测量都由同一个实验者进行。
1.5 末端DNA转移酶dUTP缺口末端标记(TUNEL)法检测凋亡
耳蜗基底膜组织切片常规脱蜡至水,按照试剂盒说明书完成余下操作后,镜下观察耳蜗毛细胞凋亡情况,光镜下400倍视野观察拍照。细胞核呈绿色颗粒伴核形态不规则或固缩的即为凋亡细胞。
1.6 免疫荧光定量分析
按免疫荧光染色按试剂盒说明书操作,使用的一抗分别为稀释浓度1∶50的多克隆兔抗LC3B(美国丹佛Cell Signaling Technology公司),1∶50的多克隆兔抗3-硝基酪氨酸(3-NT,美国比尔里卡EMD Millipore公司)。Alexa Fluor 594标记二抗的稀释浓度为1∶200,Alexa Fluor 488标记的二抗的稀释浓度为1∶100(美国Life Technologies公司)。所有切片均置于电子显微镜下观察摄片,并使用ImageJ软件量化处理。
1.7 Western blot
耳蜗基底膜组织切片经PBS清洗后按蛋白提取试剂盒说明书提取组织总蛋白,100 ℃变性5 min。然后各泳道等量蛋白进行十二烷基硫酸钠-聚丙烯酰胺凝胶(SDS-PAGE)电泳分离蛋白并转至聚偏氟乙烯(PVDF)膜。经5%的牛血清清蛋白(BSA)封闭1 h后加入相应的一抗(LC3-Ⅱ/LC3-Ⅰ、Beclin-1蛋白一抗),4 ℃过夜孵育。第2天加入相应二抗,室温孵育1.5 h。最后加入发光液,以GAPDH为内参于凝胶成像仪进行曝光拍照并进行条带灰度值分析。
1.8 统计学处理

2 结 果
2.1 抑制自噬能够降低ABR阈值
噪声暴露后1、12、24 h检测各组ABR阈值后发现,各组ABR阈值较暴露前升高。RAP组在噪声暴露后1、12、24 h各时间点的ABR阈值较对照组均明显增加;而3MA组在以上各时间点的ABR阈值较对照组则明显减少(均P<0.05)。见图1。
2.2 抑制自噬能够抑制耳蜗毛细胞中自噬标志蛋白和ROS表达水平
在噪声暴露后1、12、24 h分别检测各组耳蜗毛细胞自噬标记蛋白LC3-Ⅱ/LC3-Ⅰ和Beclin-1发现,各组噪声暴露后耳蜗毛细胞自噬标记蛋白表达水平明显升高。RAP组噪声暴露1、12、24 h耳蜗毛细胞自噬标记蛋白表达水平明显高于对照组,而3MA组以上各时间点耳蜗毛细胞自噬标记蛋白表达水平则明显低于对照组(均P<0.05),见图2。为进一步验证自噬表达与细胞程序性死亡的终末产物ROS水平的相关性,本研究检测了噪声暴露后1、12、24 h 3-NT水平后发现,噪声暴露后耳蜗毛细胞3-NT水平明显较暴露前增高,RAP组暴露1、12、24 h 3-NT水平较对照组增加;而3MA组以上各时间点3-NT水平较对照组减少(均P<0.05),见图3。
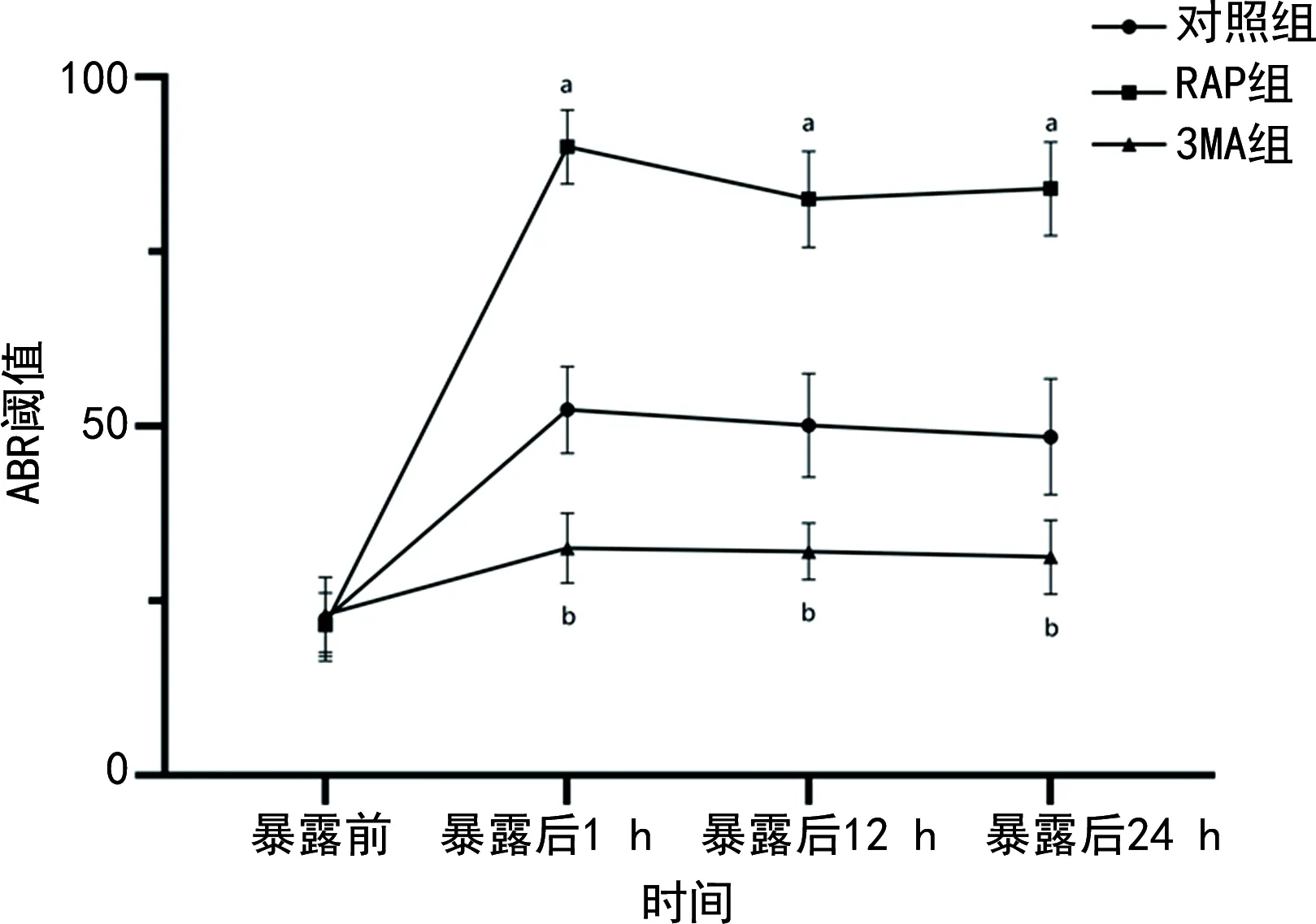
a:P<0.05,b:P<0.05,与对照组比较。
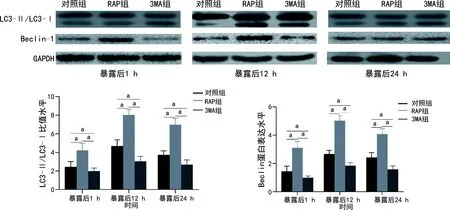
A:Western blot;B:Western blot定量分析图;a:P<0.05。
2.3 抑制自噬能够抑制毛细胞凋亡发生
为验证自噬是否会导致细胞程序性死亡,本研究检测了噪声暴露后24 h的细胞凋亡情况。结果发现噪声暴露后各组耳蜗毛细胞凋亡较暴露前明显增加,RAP组耳蜗毛细胞凋亡较对照组增加,而3MA组耳蜗毛细胞凋亡较对照组减少(均P<0.05),见图4。
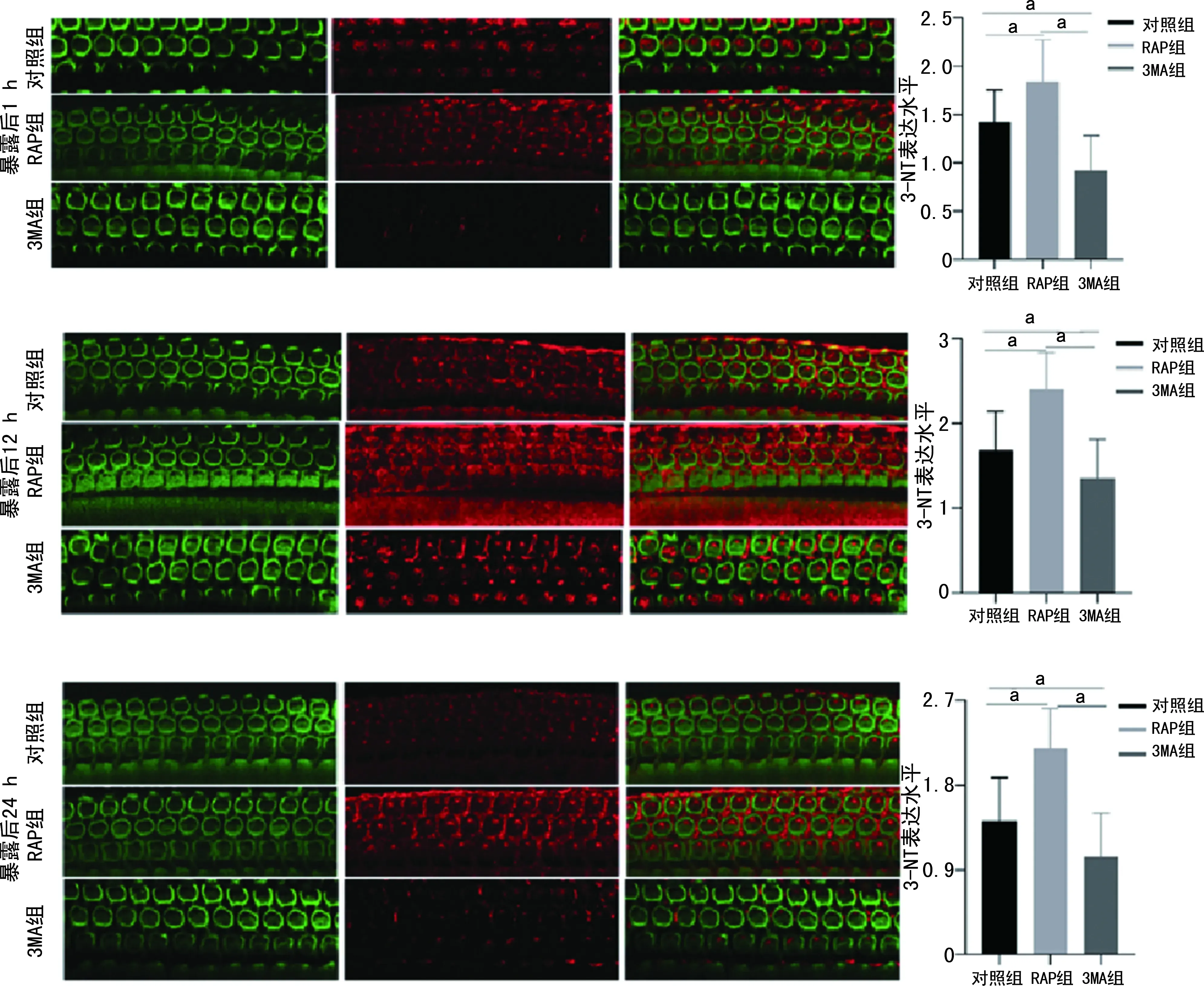
a:P<0.05。
3 讨 论
耳蜗毛细胞属高度分化的终末细胞,毛细胞作为听觉传入的重要神经信号传导细胞,它的损伤程度直接反映了听力的受损程度。但毛细胞一旦损伤凋亡,则无法再生,进而出现感音神经性耳聋。因此如何保护毛细胞避免噪声性损伤导致毛细胞死亡成为防治噪声性耳聋的首选策略。本研究发现,在噪声性耳聋发生时,自噬表达明显上调,OS标记物如3-NT水平明显增加,进而导致毛细胞的程序性凋亡。抑制自噬能够有效改善毛细胞的OS损伤,同时明显抑制毛细胞凋亡的发生。
耳蜗毛细胞极易受到噪声的损伤,既往研究证明噪声引起OS,耳蜗组织蛋白3-NT水平明显增加[10,12-13]。本研究进一步证明了这一结论,即噪声暴露会导致OS,耳蜗毛细胞中3-NT的表达水平升高。值得注意的是尽管既往文献同时发现OS导致细胞凋亡的发生,但其具体途径仍未明确阐述[14-16]。OS的形成,如超氧化物、羟基自由基和一氧化氮,可能会根据压力的不同机制而发生。噪声暴露后耳蜗中OS系统激活的确切信号机制十分复杂。目前公认有以下几种可能:超氧自由基的增加是由于其他自由基的连锁反应形成,这一过程可能是由于线粒体氧化过程加强所致;组织末梢血管收缩引起的长时间组织缺氧也可能导致OS表达增加,或组织局部的缺血再灌注损伤[14,16]。但在噪声性耳聋中针对毛细胞的损伤凋亡以上理论均有待商榷。
自噬也称为Ⅱ型程序性细胞死亡,参与了许多退变性疾病的发生发展,如帕金森病、阿尔茨海默病等神经退行性疾病,骨性关节炎、风湿性关节炎等[17-19]。但在噪声性耳聋中,自噬的具体作用机制仍存在争议。自噬和OS的相关性仍缺乏系统研究。本研究揭示噪声性耳聋急性期的自噬和OS的确切作用机制,结果表明,急性损伤期毛细胞的自噬表达明显增加,诱导细胞通过OS系统进入程序性凋亡状态。在抑制自噬表达后OS标记蛋白3-NT随之下降,同时细胞凋亡数量也明显降低。
综上所述,强噪声暴露后耳蜗毛细胞在急性损伤期通过自噬途径诱导OS系统激活,进而诱导细胞程序性死亡。这一结论为噪声性耳聋特别是急性期的防治提供了新的思路和研究方向。通过深入研究自噬和噪声性耳聋的具体作用机制可以为防治噪声性耳聋提供新的药物作用靶点及基因治疗策略。
