黄淮麦区41 个小麦品种(系)品质相关基因的分子检测
2022-04-23李春鑫赵明忠韩留鹏李正玲昝香存
李春鑫,赵明忠,韩留鹏,高 崇,李正玲,王 艳,昝香存,胡 琳
(1. 河南省农业科学院 经济作物研究所,河南 郑州 450002;2. 河南省作物分子育种研究院,河南 郑州 450002)
面制食品的制作、口感、色泽及外形受小麦蛋白质含量或面筋强度、淀粉特性、面粉色泽、籽粒硬度等品质性状的综合影响[1]。高分子质量麦谷蛋白亚基(HMW-GS)是决定小麦面团流变学特性和面筋特性的重要因素[2],由位于小麦第一同源染色体上的Glu-A1、Glu-B1 和Glu-D1 位点的基因编码[3],每个位点都含有2 个紧密连锁基因,分别编码分子质量较大的x 型和分子质量较小的y 型亚基[3]。其中,Ax1、Ax2*、Bx7+By8 和Dx5+Dy10 等亚基是公认的优质亚基[2,4]。
淀粉特性影响面条的质地,直链淀粉含量较低,煮熟后面条口感较好[5]。颗粒结合淀粉合成酶(Granule‑bound starch synthase,GBSS)又称Waxy 蛋白(简称Wx 蛋白),与直链淀粉合成密切相关。普通小麦含有Wx-A1、Wx-B1 和Wx-D1 三种Waxy 蛋白亚基,分别由位于7AS、4AL 和7DS 染色体上的基因位点编码[6]。其中,Wx-B1 蛋白亚基缺失类型直链淀粉含量较低,具有优良的面条品质[6]。
面粉色泽直接影响面制品的外观,是评价小麦品质的重要指标。面粉的色泽主要由小麦籽粒中的色素决定。其中,黄色素是小麦籽粒中的主要色素,受多基因控制。八氢番茄红素合成酶(Phytoene synthase,Psy)、ζ-胡 萝 卜 素 脱 氢 酶(ζ‑carotene desaturase,Zds)是类胡萝卜素生物合成过程中的关键酶,对小麦籽粒或面粉的黄色素含量起决定作用[7‑8]。HE 等[9‑10]先 后 克 隆 了 与 黄 色 素 含 量 相 关 的Psy-A1 和Psy-B1 基因。其中,位于Psy-A1 位点的Psy-A1a、Psy-A1b等位基因分别与高、低黄色素含量相关,位于Psy-B1 位点上的等位基因Psy-B1a、Psy-B1b、Psy-B1c分别与高、低、高黄色素含量相关。Zds 活性影响黄色素含量,目前在小麦上克隆了位于2D 和2A 染色体上的基因Zds-D1 和Zds-A1,其中Zds-A1a和Zds-D1b基因与低黄色素含量相关[11‑12]。多酚氧化酶(Polyphenol oxidase,Ppo)是引起面粉制品颜色褐变的主要因素,其主效基因Ppo-A1 和Ppo-D1 位 于 染 色 体2AL 和2DL 上[13]。SUN 等[13]开发了位于2AL 的PPO 基因标记Ppo18,能有效区分控制高、低Ppo 活性的等位基因Ppo-A1a和Ppo-A1b;HE 等[14]开发了互补标记Ppo16 和Ppo29,能有效区分Ppo-D1 位点等位基因Ppo-D1a(低Ppo 活性)和Ppo-D1b(高Ppo 活性)。脂肪氧化酶(Lipoxygenase,Lox)活性影响面粉白度,是一个受多基因控制的数量性状。目前,已经分别在小麦1AS、4A、4B、4D、5A 和5D 染色体上定位到可解释Lox活性表型变异的QTL[15]。GENG等[16]开发了互补显性标记Lox16和Lox18,能准确有效地鉴定小麦品种Lox 等位变异基因Lox-B1a(高Lox 活性)和Lox-B1b(低Lox 活性),可基本满足小麦Lox 活性辅助选择的需要。
籽粒硬度对小麦面粉颗粒大小、出粉率、淀粉破损率和加工品质有重要影响。研究表明,籽粒硬度遗传力较高,由位于5DS 上的籽粒硬度基因(Puroindoline,Pin)Pina和Pinb共同控制[17]。当Pina和Pinb同为野生型(Pina-D1a/Pinb-D1a)时,籽粒为柔软表型,两者任何一个基因发生突变或缺失都会使籽粒变硬[18]。CHEN 等[19]还发现与Pinb高度同源的基因Pinb-2v,可能对籽粒硬度具有微效的调节作用。研究表明,拥有Pinb-2v3类型的小麦品种产量相关性状表现优于Pinb-2v2类型;且与Pinb-2v2类型相比,Pinb-2v3类型小麦品种有更高的籽粒硬度和更优的籽粒性状[20]。
随着生活水平的提高和膳食习惯的改变,人们对优质小麦的需求量日益增加。黄淮麦区是我国小麦主产区,也是小麦品质改良和小麦选育的重要贡献区,但由于品种数量多、品种来源广,小麦品质涉及的位点和基因数量多。而现有相关报道对该地区小麦品质相关基因的研究大多集中于1 个或2个品质性状方面,缺乏对多个品质性状的系统检测。为此,选用41 个黄淮麦区小麦品种(系),利用23 个分子标记对面粉的面筋强度、淀粉特性、色泽及籽粒硬度等品质性状相关基因进行检测,明确相关品质基因在这些品种中的分布情况,为我国小麦品质育种提供材料和数据支持。
1 材料和方法
1.1 供试材料
供试材料为41个黄淮麦区小麦品种(系),均来自河南省农业科学院小麦研究所资源研究室种质库,所有品种(系)在试验前均经过连续2 a 的田间严格去杂和性状一致性鉴定。
1.2 基因组DNA的提取
每份材料挑选饱满种子5 粒,放置于玻璃培养皿中20 ℃培养,7~10 d 后,剪取生长健康叶片提取基因组DNA,DNA 提取方法参照LAGUDAH 等[21]的方法,用1%琼脂糖凝胶检测DNA 提取的质量和完整 性,所 有 样 品DNA 均 稀 释 为100 ng/μL,置于-20 ℃备用。
1.3 PCR扩增及分子标记检测
根据已发表的与小麦各品质特性相关的分子标记合成23 对引物,引物全部由生工生物工程(上海)股份有限公司合成。相应的引物名称、序列及其他相关信息见表1。PCR 扩增在Eppendorf 的Mastercycler Pro S (Eppendorf AG,Hamburg,Germany)上 进 行,PCR 反 应 体 系 为10 μL,包 括2.0 μL DNA 模 板、1.0 μL 10×buffer(Mg2+)、0.2 μL dNTPs(10 mmol/L)、0.2 μL 上游引物(10 pmol/μL)、0.2 μL 下 游 引 物(10 pmol/μL)、0.06 UTaqDNA polymerase(5 U/μL)、6.34 μL 无菌水。各引物PCR扩增程序对照参考文献[9‑14,16,19‑20,22‑24]进行设置。
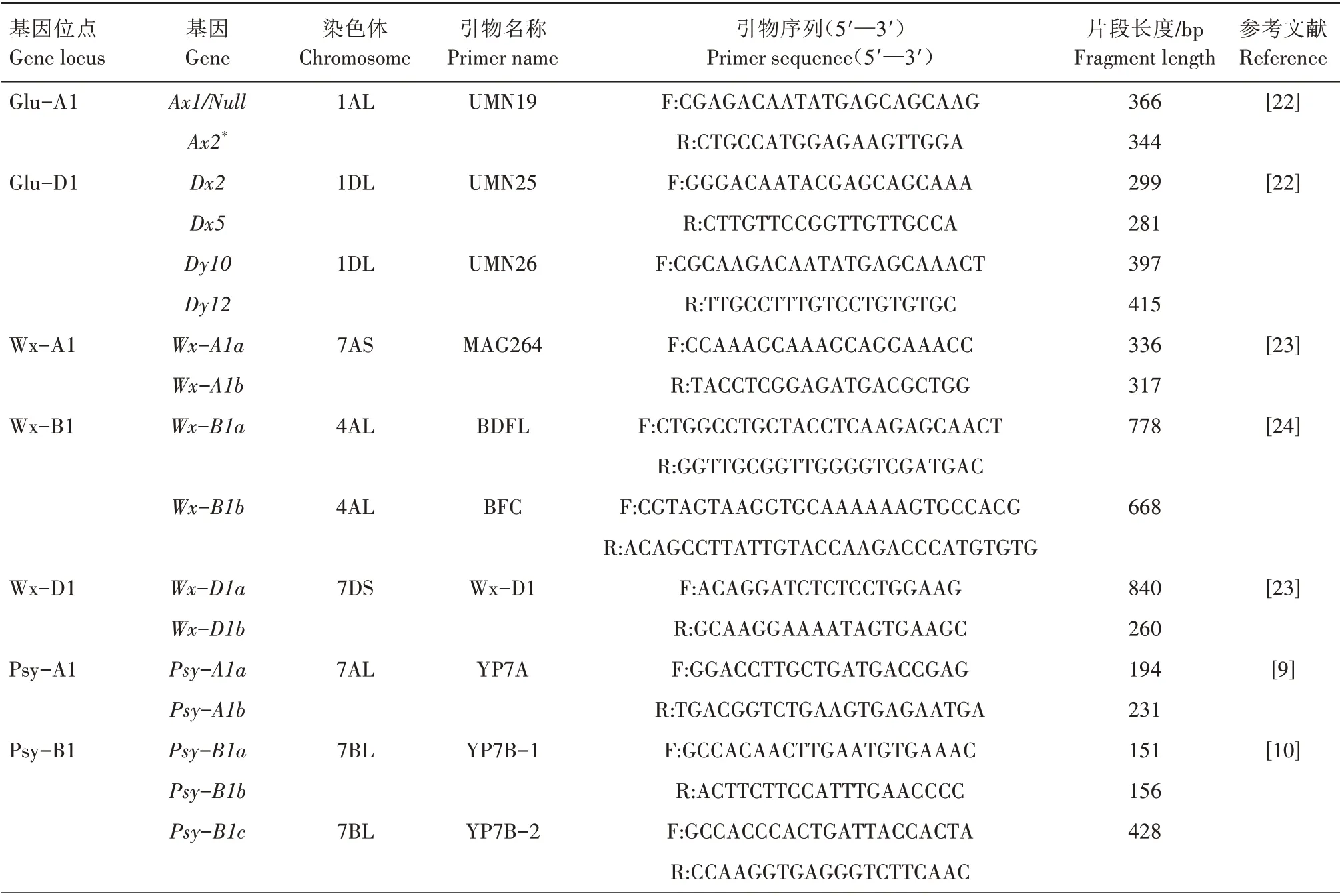
表1 小麦品质相关基因及其引物信息Tab.1 Wheat quality related genes and their primers information
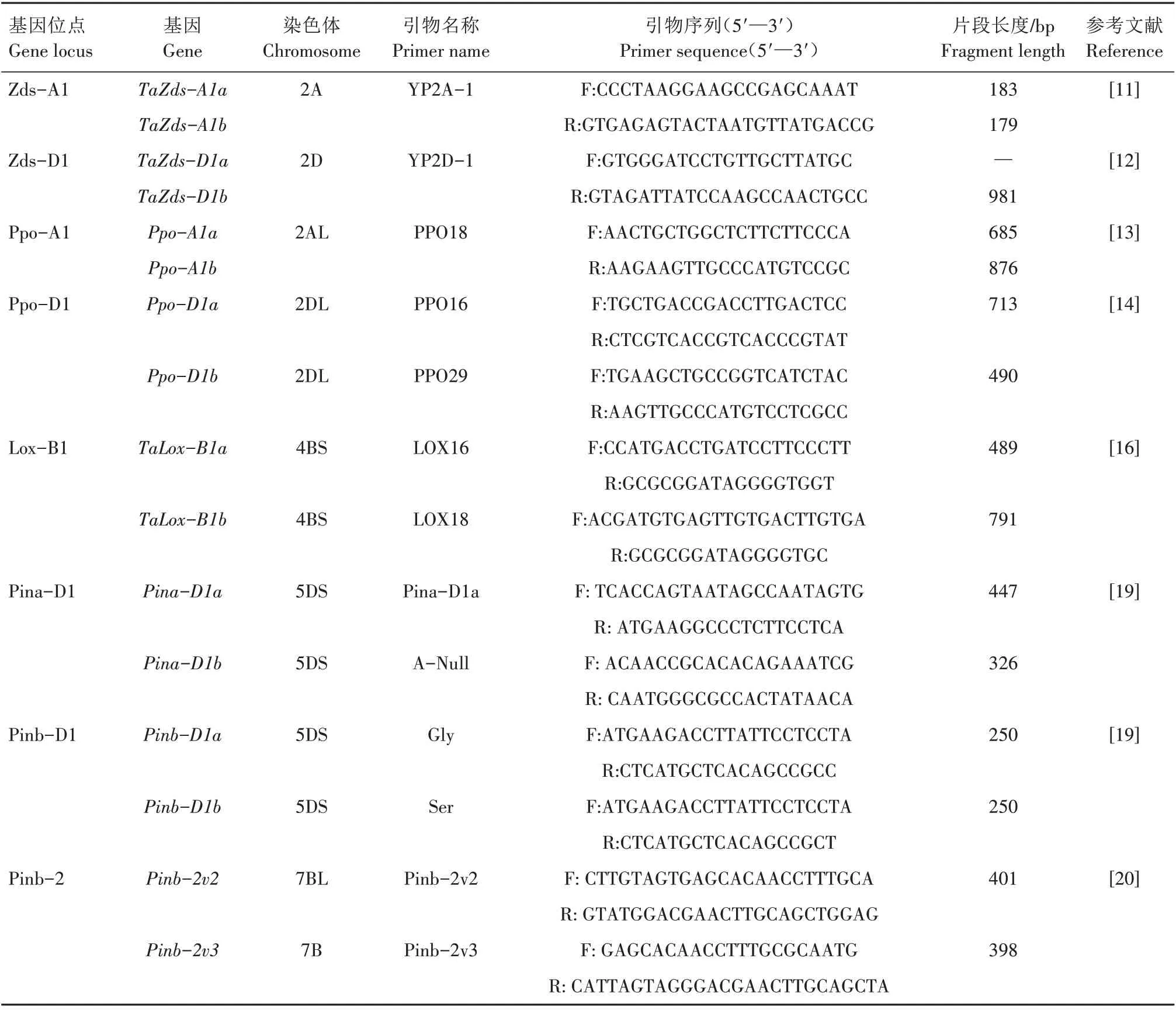
续表1 小麦品质相关基因及其引物信息Tab.1(Continued) Wheat quality related genes and their primers information
2 结果与分析
2.1 黄淮麦区41个小麦品种(系)面筋强度相关基因分子检测
41 份黄淮麦区小麦材料在Glu-A1 和Glu-D1位点共检测出5 种类型蛋白亚基基因(表2)。其中,3 份材料Glu-A1 位点为稀有亚基基因Ax2*,频率为7.32%,38 份材料Glu-A1 位点为Ax1/null;Glu-D1 位点检测出Dx5+Dy10、Dx2+Dy12基因组合类型,分别在18、23 份材料中出现,频率分别为43.90%、56.10%。
2.2 黄淮麦区41个小麦品种(系)淀粉特性相关基因分子检测
由表2 可知,在Wx-A1 位点,41 份黄淮麦区小麦材料全部表现为Wx-A1 正常表达(Wx-A1a);在Wx-B1 位 点,1 份 材 料 表 现 为Wx-B1 缺 失(Wx-B1b),频率为2.44%;在Wx-D1位点,41份材料全部表现为Wx-D1正常表达(Wx-D1a)。表明供试小麦品种(系)Wx 基因组成以野生型为主,等位变异类型不够丰富。
2.3 黄淮麦区41个小麦品种(系)面粉色泽相关基因分子检测
2.3.1 Psy-A1、Psy-B1 位点 由表2 可知,41 份黄淮麦区小麦材料中有25 份材料含Psy-A1a基因,频率为60.98%;16 份材料含Psy-A1b基因,频率为39.02%。对41 份小麦材料中Psy-B1 位点进行检测,发现Psy-B1a、Psy-B1b、Psy-B1c基因分别在16、18、7 份材料中出现,频率分别为39.03%、43.90%、17.07%,其中与低黄色素含量相关的Psy-B1b基因出现频率最高。

表2 黄淮麦区41个小麦品种(系)品质性状基因型Tab.2 Genotypes of quality traits of 41 wheat varieties(lines)in Huang-Huai wheat region
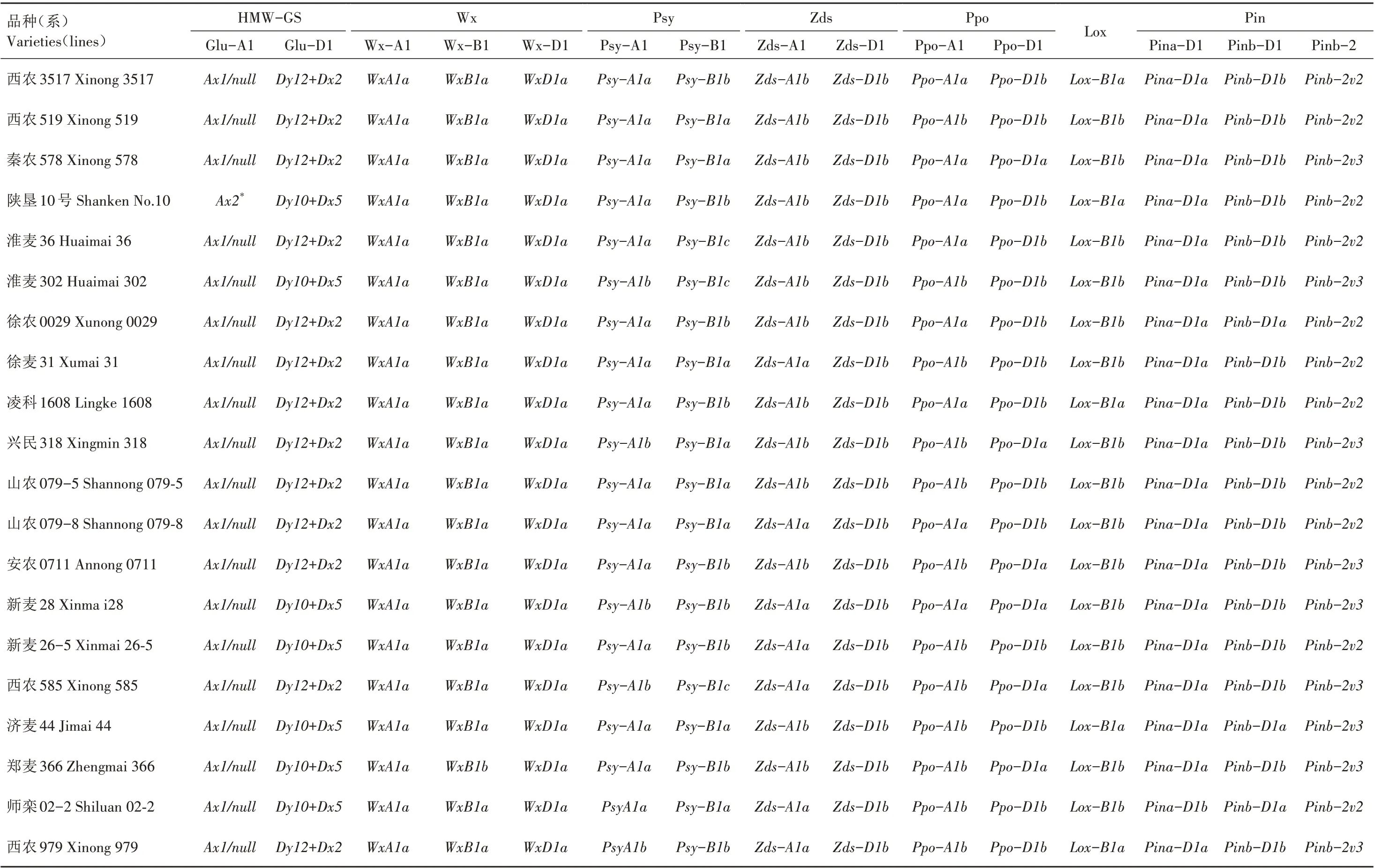
续表2 黄淮麦区41个小麦品种(系)品质性状基因型Tab.2(Continued) Genotypes of quality traits of 41 wheat varieties(lines)in Huang-Huai wheat region
2.3.2 Zds-A1、Zds-D1 位点 对41 份黄淮麦区小麦材料进行Zds-A1、Zds-D1 位点等位变异类型分析(表2),发现Zds-A1 位点存在Zds-A1a、Zds-A1b2种基因类型,频率分别为51.22%、48.78%;而在Zds-D1位点,所有材料均为Zds-D1b基因类型。
2.3.3 Ppo 基因 由表2 可知,在41 份黄淮麦区小麦材料中,Ppo-A1a、Ppo-A1b基因分别在20、21 份材料中出现,频率分别为48.78%、51.22%;Ppo-D1a、Ppo-D1b基因分别在25、16 份材料中出现,频率分别为60.98%、39.02%。
2.3.4 Lox 基因 表2 显示,在41 份黄淮麦区小麦材料中,6 份材料中出现Lox-B1a基因,频率为14.63%;35 份材料中出现Lox-B1b基因,频率为85.37%。
2.4 黄淮麦区41个小麦品种(系)籽粒硬度基因分子检测
由表2 可知,在41 份黄淮麦区小麦材料中,8 份材料表现为Pina-D1b类型,频率为19.52%;33 份材料表现为Pina-D1a类型,频率为80.48%。对Pinb位点的等位变异类型进行检测,发现Pinb-D1a类型在11 个材料中出现,频率为26.83%;Pinb-D1b类型在30 个材料中出现,频率为73.17%。对Pinb-2 位点的等位变异类型进行检测,发现表现为Pinb-2v2类型的材料有14 份,频率为34.15%;表现为Pinb-2v3类型的材料有27分,频率为65.85%。
2.5 黄淮麦区41个小麦品种(系)品质性状优质基因组合
在41 份黄淮麦区小麦材料中共检测出14 个品质相关优质基因(表2),优质基因组合共16 个(表3),对于HWM-GS 和Wx 基因来说,优质基因组合Ax2*/Dy10+Dx5和Dy10+Dx5/Wx-B1b频率均为2.44%。对于Psy、Zds 基因,优质基因组合Psy-A1b/Psy-B1b、Zds-A1a/Zds-D1b、Psy-A1b/Zds-A1a/Zds-D1b、Psy-B1b/Zds-A1a/Zds-D1b、Psy-A1b/Psy-B1b/Zds-A1a/Zds-D1b频 率 分 别 为21.95%、51.22%、4.88%、7.32%、17.07%。同时存在4 个低黄色素优质基因的品种(系)共7 份,分别是郑0856、郑1325、郑9188、郑9062-5、郑9062-9、新麦28 和西农979(表2)。对于Ppo、Lox 基因,Ppo-A1b、Ppo-D1a及Lox-B1b为优质基因,优质基因组合为Ppo-A1b/Ppo-D1a、Ppo-A1b/Lox-B1b、Ppo-A1b/Ppo-D1a/Lox-B1b,频率分别为29.27%、4.88%、2.44%。对于硬度基因Pin,Pina-D1a/Pinb-D1a基因组合表现为软质,其频率为7.32%;Pina-D1a/Pinb-D1b和Pina-D1b/Pinb-D1a基因组合表现为硬质,频率分别为73.17% 和19.51%;Pina-D1a/Pinb-D1a/Pinb-2v3、Pina-D1a/Pinb-D1b/Pinb-2v3和Pina-D1b/Pinb-D1a/Pinb-2v3基因组合出现频率分别为4.88%、43.90%和17.07%。

表3 黄淮麦区41个小麦品种(系)品质性状优异基因组合Tab.3 Excellent gene combinations for quality traits of 41 wheat varieties in Yellow-Huai wheat region
3 结论与讨论
近年来,品质育种已成为小麦育种的主要目标之一。小麦的品质性状是基因和环境共同作用的结果,受基因型、外界环境、酸碱度、热敏感性等多种因素影响[25]。在育种过程中,可以通过对高遗传力性状的分子辅助选择,提高后代选择的精准性。本研究利用23 对分子标记对黄淮麦区41 份小麦材料中的33个品质性状相关基因进行检测,研究结果可为今后小麦品质育种亲本选择和品种布局提供参考。
HMW-GS 的类别和数量是影响小麦面团特性和面筋强度的关键因素[2]。Glu-A1位点的Ax1、Ax2*基因类型优于null;Glu-D1 位点Dx5+Dy10基因组合优于普通亚基Dx2+Dy12基因组合[4‑5]。本研究中的41 份小麦材料在Glu-A1 和Glu-D1 位点共检测出5 种变异类型,类型比较丰富,其中Ax2*和Dx5+Dy10频率分别为7.32%和43.90%。Ax2*亚基是稀有优质亚基,在我国普通小麦品种中分布较少。董永梅等[26]2007年在200份我国小麦代表性地方品种中检测到的Ax2*基因频率为0.5%;郝浩楠[27]2019 年发现Ax2*基因在294份我国核心种质小麦材料中的频率为3.68%。本研究中Ax2*基因频率为7.32%,这是我国育种单位近年来重视强筋品种选育的结果。品种陕垦10 号中出现Ax2*/Dx5+Dy10优质亚基基因组合,可优先考虑作为小麦强筋育种的亲本。
小麦直链淀粉含量是影响小麦面粉品质的重要因素。Wx 蛋白作为小麦直链淀粉合成过程中的重要蛋白,包含3 种亚基,任一种Wx 蛋白亚基的缺失或失活可导致直链淀粉含量降低[6]。本研究中41份小麦材料中Wx-A1、Wx-D1 蛋白全表现为正常,仅有1 份材料郑麦366 表现为Wx-B1 蛋白缺失,远低于李式昭等[28]从四川省小麦品种中检测出的Wx-B1蛋白缺失水平(32.4%)。供试的黄淮麦区小麦材料中有关直链淀粉含量的优质种质资源类型十分稀缺,下一步应加强黄淮麦区Wx 蛋白缺失类型品种的引进及新种质资源的创制。
Psy 和Zds 作为小麦籽粒黄色素合成过程中的关键酶,其活性影响面粉的色泽[12]。本研究共检测出4 个高黄色素含量相关基因和4 个低黄色素含量相关基因,基因类型十分丰富。Psy-A1位点上高黄色素含量相关基因Psy-A1a频率为60.98%,Psy-B1位点上低黄色素含量相关基因Psy-B1b频率为43.90%,这与陈杰等[29]的研究结果一致。而相吉山等[30]研究发现,新疆维吾尔自治区小麦中Psy-A1a频率超过90%,这可能与研究时间、材料来源和地域有一定的关系。在41 份小麦材料中,来自Zds-A1 位点的高、低黄色素含量基因频率接近,Zds-D1位点只检出了低黄色素含量基因,低黄色素含量基因频率比张钰玉等[31]研究结果有大幅度提高,表明为提高面条、馒头和饺子等传统面食的亮度和白度,低黄色素含量基因已被大量用于黄淮麦区品种选育。此外,低黄色素含量相关基因最优组合Psy-A1b/Psy-B1b/Zds-A1a/Zds-D1b频率为17.07%,表明优质低黄色素含量基因已在部分品种中得到聚合,为后续选育优良品质的小麦品种奠定了较好的基础。
低Ppo活性可降低面粉在加工和贮存过程中的褐变程度,具有低Ppo 活性的小麦品种更适合加工传统面食[18,30]。本研究发现,低Ppo 活性Ppo-A1b基因在41 个小麦材料中出现频率基本与全国水平持平(52%),Ppo-D1a基因频率略高于全国水平(59.4%)[32];Ppo-A1b和Ppo-D1a基因频率高于杨子博等[33]的研究结果。Lox 活性同样影响小麦的品质,高Lox 活性可以提高面粉及其制品的白度[16]。41 份小麦材料中Lox-B1a和Lox-B1b基因频率分别为14.63%和85.37%。本研究中高Lox 活性相关基因频率偏低,与杨芳萍等[34]和张福彦等[35]的研究结果一致,这可能是因为高Lox 活性的小麦籽粒不耐长时间储藏,使携带此类基因的材料在选育过程中被淘汰。郑麦7698 是41 份小麦材料中唯一携带优质基因组合Ppo-A1b/Ppo-D1a/Lox-B1a的品种,为今后小麦籽粒色泽改良和育种亲本选择提供参考。
本研究在Pina-D1、Pinb-D1 和Pinb-2 三个位点共检测出6 种基因型,41 份小麦材料的籽粒以硬质为主,与刘红美等[36]研究发现的黄淮麦区小麦核心种质中的软质麦占比较低、硬质麦占比较高的结果相一致;而胡文静等[37]研究发现,扬州市育成小麦品种(系)以软质类型基因组合为主,这可能与不同小麦品种生态类型以及生产需求不同有关。Pina-D1 和Pinb-D1 位点产生的基因组合类型有3 种,其中Pina-D1a/Pinb-D1b基因频率最高,这与桑伟等[38]、CHEN 等[20]研究结果相符。Pinb-2 位点的Pinb-2v3基因具有很高的分布频率,与前人的研究相一致[21,39]。这可能是由于Pinb-2v3与高产性状相关基因协同效应更好,使育种家选择高产的同时保留了这一基因。籽粒硬度基因中优势基因组合为Pina-D1a/Pinb-D1a/Pinb-2v3、Pina-D1a/Pinb-D1b/Pinb-2v3和Pina-D1b/Pinb-D1a/Pinb-2v3,以上3 种基因组合的频率共达65.85%,表明黄淮麦区小麦品质改良和育种工作取得了很大的进步。
