茶树诱导抗虫性的研究进展
2022-04-19邢玉娴于广威孙晓玲
张 瑾, 邢玉娴, 韩 涛, 于广威,4,*, 孙晓玲,*
(1. 中国农业科学院茶叶研究所, 杭州 310008; 2. 农业农村部茶树生物学与资源利用重点实验室, 杭州 310008;3. 海南阿罗多甘农业科技有限公司, 海南白沙 572800; 4. 潍坊科技学院, 山东潍坊 261000)
作为世界的三大无酒精饮料之一,茶不仅具有层次丰富的口感,而且具有著名的保健功效。茶主要由茶树Camelliasinensis的芽和嫩叶制成。茶树属于山茶科山茶属,是多年生常绿灌木或乔木,生长在热带和亚热带环境中,主要分布在中国南部、印度阿萨姆和东南亚(共占世界种植总量的82%)(Zhaoetal., 2020)。目前,我国已有记载的为害茶树的害虫和害螨种类已超过800种,常见种类也有400余种。害虫为害除了显著降低茶叶产量以外,还会降低茶叶品质,干扰农事活动(Lietal., 2018; 周孝贵和肖强, 2020)。据统计,小贯松村叶蝉Matsumurascaonukii大发生年份常导致高达50%以上的茶叶产量损失,年防治费用占茶树病虫害总防治费用的60%以上(濮小英和冯明光, 2004; Chen LLetal., 2019)。目前,对茶树害虫的有效防治尚主要依赖于化学农药,化学农药的频繁使用除了导致茶树害虫产生抗药性、生态系统污染和茶叶农药残留超标外,还会大量杀伤天敌,导致茶园生态系统抗性持续降低。因此,茶树害虫频繁暴发成灾已严重制约了我国茶产业的持续健康发展,开发茶树害虫绿色防控新途径是保障茶叶安全生产的重要措施。
植物诱导抗性是指植物在植食性昆虫为害后产生的具有防御性的生理生化反应,可影响植食者的生长发育及其天敌的寄生行为等(Wu and Baldwin, 2010)。深入剖析植食性昆虫诱导植物产生的防御反应及其内在机理,可为害虫治理新途径(如植物诱导剂、害虫和天敌行为调控剂)的开发提供重要的理论基础。茶树受体感知来自植食性昆虫的相关信号后,启动早期信号通路如钙离子、活性氧和MAPK激酶,进而激活植物激素、转录因子和防御基因等信号网络,最终以局部和系统的方式启动防御代谢物、蛋白、物理防御和耐受性等防御应答保护自身免受昆虫攻击(桂连友, 2004; Sunetal., 2014; Yeetal., 2021)。茶树诱导抗虫性的研究始于20世纪90年代(许宁等, 1999)。近10年以来,随着茶树基因组注释的完成,茶树抗虫研究领域取得了突飞猛进的进展。在茶树抗虫防御系统建立、茶树抗虫基因克隆、抗虫代谢物分离鉴定以及诱导抗虫分子机制的解析等方面均取得重大突破。上述研究结果不仅在理论上可加深对害虫与茶树互作关系、茶树诱导抗虫机理等的认识,而且在实践上可为开发茶树害虫治理新途径,如茶树诱导抗虫剂、天敌引诱剂、害虫驱避剂等提供重要的理论与技术指导。至今,茶尺蠖Ectropisobliqua、灰茶尺蠖E.grisescens、小贯松村叶蝉M.onukii和茶丽纹象甲Myllocerinusaurolineatus等主要害虫为害诱导的茶树防御反应、相关调控机理及诱导抗虫性的利用等方面已经取得一些进展。本文主要对上述进展进行梳理、归纳和总结,以期为深入研究茶树诱导抗虫性的分子机理和进一步利用茶树诱导抗虫性开发害虫绿色防控技术提供科学参考。
1 害虫为害诱导茶树释放的挥发物及其生态功能
在遭受植食性昆虫为害后,植物通过改变自身挥发物组成相、提高挥发物释放量,以及释放虫害诱导特异性挥发物等方式与周围有机体进行化学信号交流,从而发挥其生态功能 (Dudarevaetal., 2006)。一方面,虫害诱导的植物挥发物(herbivore-induced plant volatiles, HIPVs)通过吸引害虫的天敌前来捕食或寄生,从而实现虫害诱导植物产生的间接防御反应(Heil, 2004);另一方面,HIPVs还可通过激活植物的钙离子信号通路和茉莉酸(jasmonic acid, JA)信号通路引起临近植物的防御警备效应或直接激活临近植物的防御反应(Erbetal., 2015; Jingetal., 2021; Yeetal., 2021)。更为有趣的是,害虫还能利用HIPVs定位交配或产卵场所,识别其嗜好寄主,从而提高后代的生长适合度和繁殖效率(Shiojirietal., 2002, 2003; Delphiaetal., 2007; Sunetal., 2014)。
茶树是多年生的木本经济作物,茶树叶片中的芳香类物质,如芳樟醇(linalool)、吲哚(indole)和水杨酸甲酯(methyl salicylate, MeSA)等是茶叶香气的重要组成成分(蔡晓明等, 2009)。被茶树害虫为害后,茶树挥发物的组成相会发生相应变化,进而影响茶叶品质。大量研究结果表明,不同害虫为害诱导茶树释放的挥发物中有许多相同的成分,如茶尺蠖、茶丽纹象甲和小贯松村叶蝉为害均可诱导茶树释放(Z)-3-己烯基醋酸酯[(Z)-3-hexenyl acetate]、(E)-β-罗勒烯[(E)-β-ocimene]、芳樟醇、(E,E)-α-法尼烯 [(E,E)-α-farnesene]、(E)-橙花叔醇 [(E)-nerolidol]、吲哚和苯乙醇(phenylethyl alcohol)等17种共有的挥发物成分;另外,由于害虫的取食策略、为害程度和虫害来源激发子的不同,不同种类害虫为害又会诱导茶树释放特异性成分,如小贯松村叶蝉为害可特异性诱导茶树释放水杨酸甲酯、4,8,12-三甲基-1,3,7,11-十三碳四烯[(E,E)-4,8,12-trimethy ltrideca-1,3,7,11-tetraene, TMTT]和香叶醇(geraniol),茶尺蠖为害可特异性诱导茶树释放(Z)-β-罗勒烯、苯甲醇(benzyl alcohol)、(Z)-3-己烯基丁酸酯 [(Z)-3-hexenyl butyrate],以及(E)-β-法尼烯[(E)-β-farnesene]等8种挥发性化合物,茶丽纹象甲为害可特异性诱导茶树释放γ-依兰油烯(γ-muurolene)和1,3,8-p-menthatriene(Sunetal., 2010, 2014; 王国昌, 2010; Caietal., 2014)。不同害虫诱导茶树释放挥发物的具体情况详见表1。
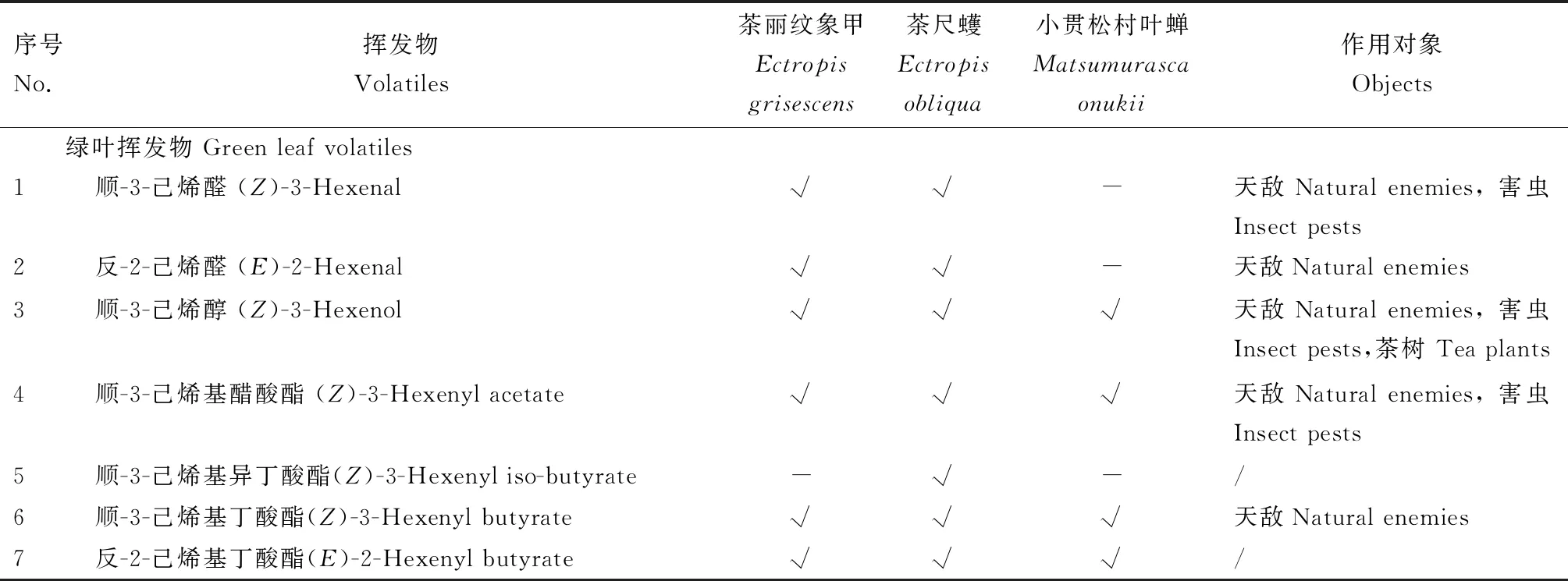
表1 3种害虫诱导茶树释放挥发物的种类及生态功能
根据茶树害虫的生物学习性,基于HIPVs开发的害虫引诱剂,与诱捕器或粘板结合使用直接诱杀害虫是HIPVs利用的重要途径之一。茶丽纹象甲是茶园中的重要食叶害虫种类之一,具有聚集分布的习性,科研工作者从茶丽纹象甲为害诱导茶树释放的挥发物中筛选出成虫引诱剂配方。有研究发现,茶丽纹象甲为害诱导茶树释放的40余种挥发物中,苯甲醇、(Z)-3-己烯基醋酸酯和香叶烯(β-myrcene)等13种挥发物能够引起象甲雌雄成虫强烈的触角电生理反应;其中,γ-萜品烯(γ-terpinene)、苯甲醇、顺-3-己烯基醋酸酯、香叶烯、苯甲醛(benzaldehyde)和顺-3-己烯醛 [(Z)-3-hexenal]对象甲的雌雄两性均具引诱活性,(E/Z)-β-罗勒烯和(E,E)-α-法尼烯仅对象甲雄成虫具有引诱作用,芳樟醇、顺-3-己烯醇[(Z)-3-hexenol]、苯乙醇和(E)-4,8,-二甲基-1,3,7-壬三烯[(E)-4,8-dimethyl-1,3,7-nonatriene, DMNT]仅对象甲雌成虫具有引诱作用(Sunetal., 2010; Caietal., 2014)。在此结果基础上,科研工作者采用正交法对具有引诱活性的化合物进行组合配比,筛选出与象甲为害苗引诱活力相当的两个配方组合,田间试验结果表明(E/Z)-β-罗勒烯和(Z)-3-己烯基醋酸酯两种物质的组合对茶丽纹象甲具有显著的引诱活性(Sunetal., 2012)。茶尺蠖隶属于鳞翅目,其幼虫暴发成灾常给茶叶生产带来严重损失,其成虫利用寄主植物挥发物进行交配和产卵场所的定位。在茶尺蠖诱导茶树释放的众多挥发物中,苯甲醇、顺-3-己烯基丁酸酯[(Z)-3-hexenyl butyrate]和顺-3-己烯醛对未交配的茶尺蠖雌雄成虫均具有显著的引诱效果,而芳樟醇和苯乙腈(phenylacetonitrile)对二者均有驱避作用;交配后的茶尺蠖雌雄成虫可以被顺-3-己烯基丁酸酯、顺-3-己烯基醋酸酯和顺-3-己烯醛所引诱,由此推测茶尺蠖可能利用上述3种物质定位产卵场所(Sunetal., 2014)。田间生测试验结果表明,顺-3-己烯基醋酸酯、苯甲醇、顺-3-己烯基丁酸酯和顺-3-己烯醛4种物质按照一定比例混合对茶尺蠖成虫具有显著的诱集效果,可作为茶尺蠖成虫引诱剂在田间使用(Sunetal., 2016)。小贯松村叶蝉是茶园中的头号害虫,以成若虫刺吸茶树汁液为害,世代重叠严重,难于防治。研究发现小贯松村叶蝉利用顺-3-己烯基醋酸酯定位产卵场所,其释放量在叶蝉抗、感茶树品种间存在明显差异(Xinetal., 2016; 任倩倩, 2020),小贯松村叶蝉会根据顺-3-己烯基醋酸酯的释放量为后代选择更为合适的寄主(Xinetal., 2016)。目前,利用寄主植物挥发物,已研制出多个具有田间引诱活性的小贯松村叶蝉引诱剂(Caietal., 2020);利用芳香植物挥发物,研制出1种小贯松村叶蝉的驱避剂(Chenetal., 2020)。
利用临近茶树对HIPVs的感知作用,开发茶树诱导抗虫剂是HIPVs利用的另外一个重要方面。Dong等(2011)发现茶小卷叶蛾Adoxophyeshonmai和神泽氏叶螨Tetranychuskanzawai为害可诱导茶树释放法尼烯、芳樟醇和顺-3-己烯醇等挥发物,并且这些物质可从为害部位通过维管束传导至健康部位,但是由此带来的对健康部位次生代谢物积累的影响显著低于挥发物暴露处理。进而,有研究发现茶尺蠖幼虫为害诱导茶树释放的挥发物可作为化学信号物质在茶树间传递,并具有激活临近植株防御警备/直接防御反应的作用(雷舒等, 2016; Jingetal., 2021)。至今,已发现顺-3-己烯醇、DMNT、芳樟醇、α-法尼烯、吲哚和橙花叔醇(nerolidol)等化合物是茶树间化学信息交流的关键物质,可引起临近茶树防御警备或激活临近茶树的防御反应(Xinetal., 2016, 2019; Jingetal., 2019, 2021; Chenetal., 2020; Yeetal., 2021)。例如,有研究发现茶尺蠖幼虫取食诱导茶树释放的挥发物(顺)-3-己烯醇、芳樟醇、α-法尼烯和萜烯同系物DMNT可诱导邻近茶树β-罗勒烯的释放,进而提高邻近健康茶树对茶尺蠖成虫的驱避能力(Jingetal., 2021);外用顺-3-己烯醇处理可激活茶树茉莉酸和乙烯(ethane, ET)信号通路,并由此提高茶树对茶尺蠖的直接和间接防御反应,进一步研究发现顺-3-己烯醇是在糖基转移酶的作用下生成顺-3-己烯醇糖苷而发挥其调控功能的(Xinetal., 2016, 2019; Jingetal., 2019);DMNT和吲哚暴露处理可以通过激活茶树茉莉酸通路和钙离子信号通路,通过改变下游次生代谢物的积累从而提高茶树的防御反应(Jingetal., 2021; Yeetal., 2021);橙花叔醇暴露处理可激活茶树JA、过氧化氢和脱落酸信号途径,通过提高下游多酚氧化酶、胼胝质和几丁质酶的活性,而提高茶树对小贯松村叶蝉和茶炭疽病的抗性(Chenetal., 2020)。以上研究结果为茶树诱导抗虫剂的开发提供了理论依据和技术支持。

续表1 Table 1 continued
2 调控茶树诱导抗虫性的重要防御信号网络
感知植食性昆虫攻击后,植物会启动早期信号事件(数秒和数分钟内)。主要包括胞内Ca2+含量升高、活性氧(reactive oxygen species, ROS)产生、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)活性增强等,这一现象已在拟南芥和水稻等模式植物中得到证实(Liuetal., 2018; Erb and Reymond, 2019),而茶树仅在近年来才有少量报道。通过对茶尺蠖幼虫为害后茶树转录组学结果的分析,Yang等(2019)发现识别虫害相关分子模式的受体、Ca2+信号通路、MAPK级联、ROS和众多转录因子参与了茶树抗虫防御的早期信号事件。挥发性吲哚暴露处理可提高随后茶尺蠖幼虫为害诱导的茶树早期信号元件基因在转录水平的上调,如钙调蛋白(calmodulin, CAM)基因、类钙调蛋白(calmodulin-like protein, CML)基因、Ca2+/钙调素依赖性蛋白激酶(Ca2+/calmodulin-dependent protein kinase, CDPK)基因、MPK基因CsMPK2和CsMPK3, WRKY转录因子CsWRKY7和CsWRKY75, 以及MYC基因CsMYC2a和CsMYC2b等(Yeetal., 2021)。
在植物应答虫害胁迫过程中,激素信号通路在早期信号到下游的转录水平、蛋白水平和代谢水平的重组间发挥着重要的调控作用(Schuman and Baldwin, 2016; Howeetal., 2018)。植物识别到害虫为害后,通过激活植物体内复杂的植物激素信号网络,调控下游相应防御代谢物的积累而产生抗虫性。茉莉酸和水杨酸(salicylic acid, SA)信号转导通路是其中的核心通路,植物生长素、脱落酸、乙烯和赤霉素等其他植物激素通路则通过与茉莉酸和水杨酸信号转导通路的相互作用而发挥其调控功能(Shigenaga and Argueso, 2016)。以上植物激素信号通路在模式植物抗虫性机理的解析和调控方面已得到较为深入研究(Zhouetal., 2009; Lietal., 2015; Erb and Reymond, 2019)。由于茶树还未建立成熟的遗传转化体系,目前害虫诱导茶树抗虫机理的研究仅从以下几个方面进行了粗浅的解析:(1)害虫为害/模拟为害对茶树体内信号分子含量的影响;(2)害虫为害/模拟为害对茶树信号通路重要合成酶基因和响应基因转录表达,以及下游次生代谢物质含量的影响;(3)植物激素及相关信号通路抑制子处理后对茶树抗虫性、相关基因表达量和防御化合物含量的影响。已证明,JA和SA可参与茶树对多种害虫的防御应答,其中,JA通路在调控茶树防御反应过程中发挥重要作用;此外,植物生长素信号通路、ABA通路和赤霉素通路同时参与了茶树的诱导防御反应(Liaoetal., 2019; Zhang Jetal., 2020; 邢玉娴, 2021)。 但是,上述植物激素信号网络对茶树诱导抗虫性的调控机理尚需深入解析。
3 茶树茉莉酸信号通路的研究进展及利用
如前所述,茉莉酸信号转导通路在茶树抵御害虫为害过程中发挥着重要作用,也是目前茶树应答害虫胁迫机制研究得最为详尽的一条通路。脂氧合酶(lipoxygenase, LOX)参与植物抗虫防御反应,是JA和绿叶挥发物合成所需的关键酶。Zhu等(2018)深入研究了CsLOX家族基因系统发育、基因结构及其蛋白特征,分析了CsLOX家族基因成员在茶树应答生物和非生物胁迫时的表达特征,以及6个CsLOX基因的可变剪切情况,为研究LOX在茶树诱导防御反应中的调控机理提供了分子基础。此外,12-氧-二烯酸还原酶3(12-oxophytodienoate reductase, OPR3)和丙二烯氧化物环化酶(allene oxide cyclase, AOC)等茉莉酸信号通路合成相关基因已经被克隆和鉴定,它们的组织表达特征及其在生物胁迫和激素处理后的时空表达谱已经被较为详尽地研究,并据此对它们的功能进行了预测(Wangetal., 2016; Xinetal., 2017)。两个酰基辅酶A氧化酶(acyl-CoA oxidase)基因CsACX1和CsACX3是已分离鉴定的茶树酰基辅酶A氧化酶家族成员基因,能被生物和非生物压力诱导,并参与JA合成。另有研究报道茶树CsJAZs家族基因差异应答生物因子(茶尺蠖、叶蝉和炭疽病菌诱导)、非生物因子(机械损伤、干旱)和植物激素(乙烯、赤霉素和脱落酸),而且在蛋白水平上证明了CsJAZs与CsMYC2的互作关系。因此,CsJAZ家族基因的鉴定和分析为研究JA通路介导的生物和非生物胁迫提供了重要的分子基础(Zhang Xetal., 2020; Zhengetal., 2020)。转录组学分析结果表明,茶尺蠖取食显著上调了9个CsLOX基因、5个CsAOS基因、4个CsAOC基因、3个CsOPR基因、2个CsJMT基因和 9个CsJAZ基因等的转录表达(Yangetal., 2019),提示了这些基因在茶树抵御茶尺蠖为害过程中发挥着重要作用,但是它们的确切生物学功能尚待遗传转化体系建立后进一步验证。
大量研究结果表明,茶树茉莉酸信号通路正调控茶树对害虫的直接与间接抗性。 多酚氧化酶(polyphenol oxidase, PPO)和丝氨酸蛋白酶抑制剂(serine protease inhibitor, SPI)是茶树体内重要的抗虫化学靶标(桂连友等, 2005; Yangetal., 2013; Zhang Jetal., 2020)。外用JA或茉莉酸甲酯处理,可显著提高茶苗叶片中PPO和SPI的活性,并发现二者活性与JA或茉莉酸甲酯的浓度呈显著正相关,但与取食不同浓度激素处理苗的茶尺蠖幼虫生长速率呈显著负相关(桂连友等, 2005; Yangetal., 2013);另有研究结果表明,外用MeJA处理茶树可显著提高茶尺蠖单白绵绒茧蜂Apantelessp.对茶尺蠖幼虫的寄生率,进一步的研究发现这是由于MeJA处理诱导茶树释放了与茶尺蠖为害相似的挥发物组成相所致(桂连友等, 2004; Caietal., 2014; 孙晓玲等, 2016)。进而,有研究利用茉莉酸和两个茉莉酸通路抑制子[二乙基二硫代氨基甲酸钠(三水)(sodium diethyldithiocarbamate trihydrate, DIECA)和水杨羟肟酸(salicylhydroxamic acid, SHAM)]进一步证实茉莉酸信号通路正调控茶树对茶尺蠖的直接抗性(Zhang Jetal., 2020)。鉴于此,利用茉莉酸信号通路调控茶树的防御反应,开发茶树的诱导抗虫剂等绿色防控技术就成为茶树植物保护工作者关注的热点问题。
基于诱导抗虫性开发作物的诱导抗虫剂,并合理使用,是实现害虫绿色、有效及可持续治理的重要途径之一。在植物体内,游离JA在茉莉酸-氨基酸合成酶的催化下与异亮氨酸形成(+)-7-异-茉莉酸-L-异亮氨酸(JA-Ile),JA-Ile与受体蛋白复合体COI1-JAZ1结合后方可启动植物体内的系列生理与生化反应(Sheardetal., 2010)。有研究工作者利用有机合成的方法获得了JA-Ile结构类似物JA-Ile-macrolactone 5b,并发现利用该物质处理不仅能够提高茶树对茶尺蠖的直接抗性,而且还能够降低茶炭疽病对茶树的侵染速率。机理研究结果表明该物质可以诱导CsOPR3表达量的显著上调,同时抑制了黄酮类化合物圣草酚-5-氧-葡萄糖苷的累积,从而提高了茶树对茶尺蠖的抗性(Linetal., 2020)。由于这些化学激发子本身对害虫和天敌没有杀伤作用,加上这些化合物容易降解,因此据此开发的诱导抗虫剂是今后开发茶园有害生物可持续治理技术的一个重要发展方向。
4 小结及展望
植物的诱导防御反应是一个复杂的生理生化过程,涉及到植物对植食性昆虫为害的识别、多种信号转导通路(如JA、SA、乙烯、过氧化氢、赤霉素、ABA等)的激活与互作、植物防御相关基因的表达和调控,以及防御化合物的合成和积累等多个方面(Chen SLetal., 2019; Chen XYetal., 2019; Erb and Reymond, 2019; Erb and Kliebenstein, 2020; Maetal., 2020; Erbetal., 2021)。这一反应不仅受多种环境因子(如温度、光照等)的影响,而且还受到植食性昆虫取食策略或习性(如口器类型、寄主范围等)的影响(Acevedoetal., 2015; Züst and Agrawal, 2016),并且这一防御反应具有植物种类、基因型、生育期、部位以及植食性昆虫种类、种群、龄期等的特异性(Lankau and Strauss, 2008; Poelman and Kessler, 2016; De Bobadillaetal., 2021)。本文基于茶树主要害虫为害诱导茶树防御反应及其相关调控机理的研究,首先综述了不同害虫为害诱导茶树释放的挥发物种类、生态功能及其在绿色防控中的应用等多个方面;其次,概述了参与调控茶树诱导抗虫性的重要防御信号网络,并重点阐述了茶树茉莉酸信号转导通路的研究进展。此外,文中还综述了基于茶树诱导抗虫性开发的害虫引诱剂和茶树诱导抗虫剂等茶树害虫绿色防控技术。茶树是多年生的木本经济作物,异花授粉,基因组杂合度高,生育周期长,目前尚无成熟的遗传转化体系。受上述因素制约,害虫诱导茶树防御反应的分子调控机制尚无法深入展开。但是,茶原产中国,复杂的生态环境和长期演化选择形成了丰富多样的茶树种质资源,尤其是野生资源丰富,这些资源具有重要育种价值的抗虫基因和主效数量抗性位点,这非常有利于重要抗虫基因的鉴定分离、防御信号通路网络的建立、抗虫性与生长发育平衡调控机理的解析以及茶树-害虫互作机理的深入研究。从现有的研究进展可以看出,茶树诱导防御反应与模式植物有许多相同之处,但是又有着茶树与不同害虫研究系统的特异性。利用茶树诱导抗虫性开发害虫的绿色防控技术,并加以合理使用,有望减少化学农药的使用量,逐步恢复茶园生态系统自身的抗性,从而彻底解决长期大量使用化学农药引发的环境污染、生态系统平衡失调以及食品安全等问题。今后,除了进一步深入挖掘茶树害虫诱导的防御反应分子机理之外,还可从茶树诱导抗虫性的利用方面展开以下研究:(1)建立茶树诱导抗虫剂的高通量筛选模型,从天然存在的小分子化合物中筛选活性物质,并研究其配套使用技术;(2)结合茶树害虫性信息素的研究,筛选虫害诱导挥发物中的活性组分,开发茶树害虫性信息素增效剂;(3)进一步研究茶树害虫诱导特异性挥发物的生态功能,明确其在茶树-害虫-天敌三级营养关系中的作用,从而开发出高效的天敌引诱剂。本文通过综述茶树诱导抗虫性相关研究的进展,旨在为深入研究其分子机理和进一步利用茶树诱导抗虫性开发害虫绿色防控技术提供参考
