西沙热带珊瑚岛典型乔木叶片性状和养分再吸收特征
2022-04-16龙靖黄耀刘占锋简曙光魏丽萍王俊
龙靖 ,黄耀,刘占锋,简曙光,魏丽萍,王俊*
1. 中国科学院海岛与海岸带生态恢复重点实验室/中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049
珊瑚岛是海洋中的一种特殊岛屿,由珊瑚虫死后的残体堆积而成,在热带海域分布广泛。中国西沙群岛是一群处于南(中国)海中地质史比较年轻的珊瑚岛,这些岛屿面积小且远离大陆,陆域植被以单种优势群落为主,生态系统结构简单而脆弱(广东省植物研究所西沙群岛植物调查队,1977)。基于热带珊瑚岛独特的生境特点及其在开发利用中面临的生境破碎化、植被退化、有害植物入侵等生态问题,近年来有关岛屿植被恢复与保护的研究逐渐受到生态学者的关注(黄宝荣等,2009)。
植物养分再吸收是指植物器官(叶片、细枝和根)在凋落前,将营养物质从衰老组织转移到生长组织的过程,是植物重要的养分保存策略(Aerts,1996;Aerts et al.,1999)。养分再吸收对植物的养分收支平衡至关重要,是植物和生态系统养分经济的重要决定因素(Zhang et al.,2015)。目前,陆地生态系统植物叶片养分再吸收模式及其调控因素在多个尺度上已得到广泛研究,包括群落(Pang et al.,2018)、生态系统(Chen et al.,2021)、区域(Tang et al.,2013)和全球尺度(Yuan et al.,2009a)等。全球尺度上,大约有62.1%的氮(N)和64.9%的磷(P)被再吸收到植物活组织中(Vergutz et al.,2012)。由于地理位置封闭隔绝、生态环境特殊,目前我们对于海岛植物,尤其是热带珊瑚岛植物的养分再吸收模式尚知之甚少。
植物叶片性状是易于观察和度量的植物特征,对气候的变化非常敏感,能够反映植物在复杂生境下的适应能力和自我调节能力(刘晓娟等,2015;王晶媛等,2017)。每个叶片性状在进化选择下都有其对应的适应策略,可用于预测植物适应环境变化后所形成的生态策略(Poorter et al.,2006)。其中,比叶面积、叶片养分浓度和养分再吸收在植物资源获取和保留的能力中起着关键作用(Niinemets et al.,2003;Wright et al.,2005),并因此影响许多关键生态系统过程。热带珊瑚岛生境条件恶劣,生长在该立地条件下的植物必然存在其有效的适应机制。因此,有必要针对热带珊瑚岛优势物种的叶片性状开展研究,以评估其在热带珊瑚岛特殊环境下的生态策略。
土壤磷是影响植物生长和功能的重要因素,在热带地区被认为是最主要的限制性养分之一(Elser et al.,2007;Ji et al.,2018)。Yuan et al.(2009a)的全球尺度荟萃分析表明,由于磷的限制,热带地区植物磷再吸收效率要高于氮再吸收效率。然而,由于鸟粪的输入和贝壳类动物残体的沉积,同属热带气候条件下的西沙珊瑚岛的土壤磷含量显著高于中国其他热带亚热带地区(龚子同等,2013)。在这种富磷生境下,岛屿上的植被很可能受到氮限制而非磷限制。因此,西沙热带珊瑚岛植物的养分再吸收特征和养分利用策略可能与同纬度地区的热带植被相比存在很大的差异,相关研究的开展将有助于丰富我们对植物养分再吸收模式和生物地球化学循环的认知。
相比近纬度地区的热带雨林或季雨林顶级群落,中国西沙热带珊瑚岛的自然环境条件极为恶劣,包括高温、高盐、强碱、长期强烈的紫外辐射以及频繁的自然灾害(台风和潮汐)等,植物很难定居和生长(Zhang et al.,2019)。这使得西沙珊瑚岛天然植物群落中的优势种少但优势度突出,形成了以抗风桐(Pisonia grandis)、海岸桐(Guettarda speciosa)、橙花破布木(Cordia subcordata)、榄仁树(Terminalia catappa)等为主的单优乔木群落(任海等,2017)。研究表明,植物可以通过增加养分再吸收来提高其自身的整体适应性,这种现象在生境恶劣的条件下尤为明显(Gloser,2005;Marty et al.,2009;Zhang et al.,2015)。例如,刘宏伟等(2015)发现生长在条件恶劣的石灰岩地区的植物具有较高的养分再吸收程度。周丽丽等(2019)发现木麻黄(Casuarina equisetifolia)和湿地松(Pinus elliottii)会通过较高的养分再吸收效率来适应高盐干旱的滨海沙地生境。因此,研究珊瑚岛生境下植物叶片养分再吸收特征及其影响因素,对于深入理解海岛植物适应性和海岛陆域生态系统养分循环至关重要。本研究以中国西沙海域东岛的两种单优乔木树种(抗风桐和海岸桐)为研究对象,对养分再吸收特征和叶片性状进行了测定分析,旨在揭示西沙珊瑚岛典型优势树种的养分保存机制,明晰热带珊瑚岛的优势树种是否会通过较高的养分再吸收效率来适应热珊瑚岛特殊生境。研究结果将有助于探讨热带珊瑚岛环境下植物的养分利用策略,为中国西沙海岛天然植被的保护与管理提供科学依据。
1 材料与方法
1.1 研究地概况
本研究地点位于南海西沙海域的东岛(112°43′—112°45′E,16°39′—16°41′N),该岛长约2.4 km,宽约0.5 km,面积约为1.7 km2,海拔约3—6 m(刘晓瞳等,2017)。东岛属于典型的热带海洋性气候,年平均气温 26—27 ℃,年平均降水量为1500 mm,年平均蒸发量为2500 mm(Huang et al.,2020)。该岛受人类干扰影响较小,且拥有中国鸟类密度最大的自然保护区(主要为红脚鲣鸟Sula sula)(王森浩等,2019),鸟粪资源丰富。岛上的自然森林群落结构简单,大多为单优种群落,主要由抗风桐林(P.grandis)和海岸桐林(G.speciosa)组成。抗风桐和海岸桐均具优良的耐盐碱和耐旱性(王馨慧等,2017;李晓盈等,2021),是当地气候和土壤条件下的顶级树种。其中抗风桐为常绿大乔木,生长速度快、断枝可再殖,平均树高约9.8 m,平均胸径约10.54 cm,主要分布于岛屿内部的平坦区域,约占岛屿植被面积的50%;海岸桐为常绿小乔木,抗风性强,平均树高约2.8 m,平均胸径约3.88 cm,主要分布于岛屿周边的沙丘区域,约占岛屿植被面积的 20%。林下植物以海滨木巴戟(Morinda citrifolia)和锥穗钝叶草(Stenotaphrum micranthum)为主。岛上成熟的土壤以石灰质腐殖土为主,未成熟土壤以冲积珊瑚砂为主(广东省植物研究所西沙群岛植物调查队,1977)。
1.2 样品采集与测定分析
1.2.1 植物样品采集与分析
在抗风桐林和海岸桐林群落内,设置 5个 15 m×15 m的样方进行采样。在每个样方中选取3—5株大小、长势相同且生长良好的个体作为取样植株并进行标记。2017年8月,从各样株的树冠中上部采集 10—20片左右的成熟叶片,同一样方内的样品混合作为1个混合样。将采集的叶片密封在聚乙烯袋中以保持水分,并尽快运往实验室用于叶片养分含量和相关叶片性状的测定。同年10月,通过轻轻摇晃标记植株的树干和树枝收集新近衰老叶片,衰老叶片可以通过叶片颜色进行辨认(通常为黄色)。衰老叶的处理方法同成熟叶。
采用叶面积仪(LI-3000C,LI-COR,USA)来测定成熟叶片的叶面积,并计算平均叶片面积(Mean leaf area,MLA)。测定完毕后,将其置于烘箱内105 ℃杀青30 min,然后在65 ℃下烘干72 h后测定干质量,比叶面积(Specific leaf area,SLA)即为叶片面积与叶片干质量之比。将烘干后的成熟叶片磨碎,用重铬酸钾法、改良的凯氏定氮法和钼锑抗比色法分别测定成熟叶片的碳含量(Green leaf carbon content,w(C)gr)、氮含量(Green leaf nitrogen content,w(N)gr)和磷含量(Green leaf phosphorus content,w(P)gr),用范式洗涤纤维分析法测定成熟叶片酸性洗涤纤维(Green leaf acid detergent fiber content,ADFgr)、纤维素(Green leaf cellulose content,Cellulosegr)和木质素(Green leaf lignin content,Ligningr)的含量(Rowland and Roberts,1994)。衰老叶片养分的测定方法同成熟叶片。
养分再吸收通常用养分再吸收效率来量化和检验,即植物在衰老过程中叶片再吸收的养分百分比,其计算公式如下(Vergutz,2012):

式中:
R——养分再吸收效率,%;
ws——成熟叶养分含量,mg·g−1;
wg——衰老叶养分含量,mg·g−1;
F——叶片衰老过程中的质量损失校正因子,本研究中两物种的F值为 0.784(Vergutz et al.,2012)。
另外,用养分再吸收程度来描述植物的养分再吸收能力,其定义为叶片彻底衰老后的养分最终含量(mg·g−1),能反映养分再吸收的生理极限(Killingbeck,1996)。
1.2.2 土壤样品采集与分析
在抗风桐林和海岸桐林群落的各样方中,随机选取5个点,用内径为4 cm的土钻采集深度为0—20 cm和20—40 cm的土壤样品,同层混合,作为一个混合样。每个土壤样品在去除砂石和植物残体后过2 mm的筛网,最后将土壤样品装入聚乙烯袋后带回实验室进行分析。
采集的土壤样品在室内进行风干以后,采用玻璃电极pH计测定土壤pH值,土水体积比为1꞉2.5。采用溶解-烘干法测定土壤含盐量(Soil salinity content,Salinity)。采用重铬酸钾氧化-外加热法、改良的凯氏定氮法和钼锑抗比色法分别测定土壤样品的有机质(Soil organic matter,SOM)含量、总氮(Soil total nitrogen,TN)含量和总磷(Soil total phosphorus,TP)含量。采用NaHCO3浸提-钼锑抗比色法测定土壤有效磷(Soil available phosphorus,AP)含量(刘光崧,1996)。
1.3 数据分析
统计分析前使用Shapiro-Wilk检验和Levene’s检验来检验数据的正态性和方差齐性,并在分析前对不符合条件的数据进行了log 10转换。利用t检验比较抗风桐林和海岸桐林土壤化学性质的差异,以及抗风桐和海岸桐叶片性状和养分再吸收之间的差异。利用Pearson相关性分析和线性回归分析法分析养分再吸收效率与土壤养分和叶片性状之间的关系。数据分析和作图均使用R 3.6.1完成。
2 结果与分析
2.1 抗风桐林和海岸桐林的土壤化学性质
抗风桐林和海岸桐林的土壤养分含量总体上随着土壤深度的增加而显著减少(表1)。抗风桐林中不同深度土层的pH值均显著低于海岸桐林,但二者的土壤含盐量(Salinity)无显著差异。抗风桐林的土壤有机质(SOM)和全氮(TN)含量在不同深度土层均显著高于海岸桐林。就 0—20 cm土层而言,抗风桐林土壤全磷(TP)和有效磷(AP)含量显著高于海岸桐林,但土壤氮磷比(N꞉Psoil)显著低于海岸桐林。抗风桐林和海岸桐林间20—40 cm土层的全磷、有效磷及氮磷比差异不显著。总体来看,抗风桐林的土壤肥力高于海岸桐林。
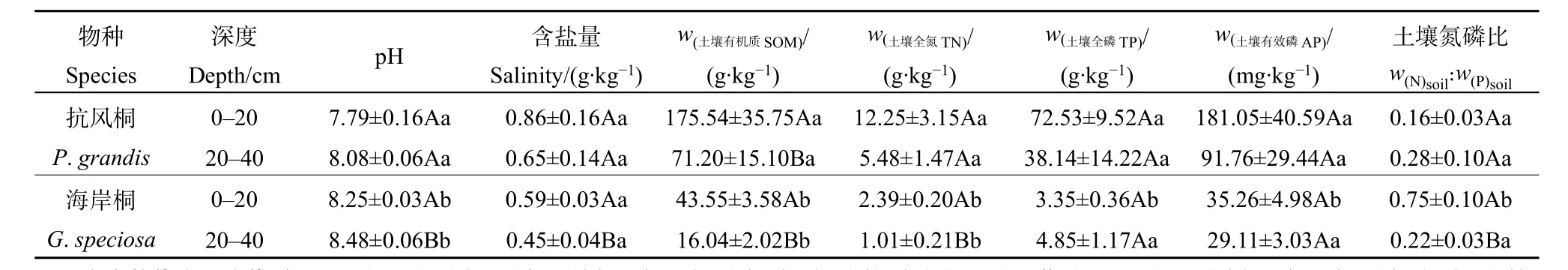
表1 抗风桐林和海岸桐林的土壤化学性质Table 1 Soil chemical properties of P. grandis and G. speciosa forests
2.2 抗风桐和海岸桐的叶片性状
抗风桐的平均叶面积(MLA)显著低于海岸桐,而抗风桐的比叶面积(SLA)则显著高于海岸桐(表2)。抗风桐的成熟叶氮含量w(N)gr和氮磷比w(N)gr꞉w(P)gr显著高于海岸桐,而成熟叶碳含量w(C)gr、磷含量w(P)gr、碳氮比w(C)gr꞉w(N)gr和木质素与氮含量之比w(lignin)gr꞉w(N)gr均显著低于海岸桐。另外,抗风桐在成熟叶片和衰老叶片中的木质素含量和纤维素含量均显著低于海岸桐,并且抗风桐的衰老叶氮含量显著高于海岸桐(图1d),说明抗风桐的凋落物质量优于海岸桐。
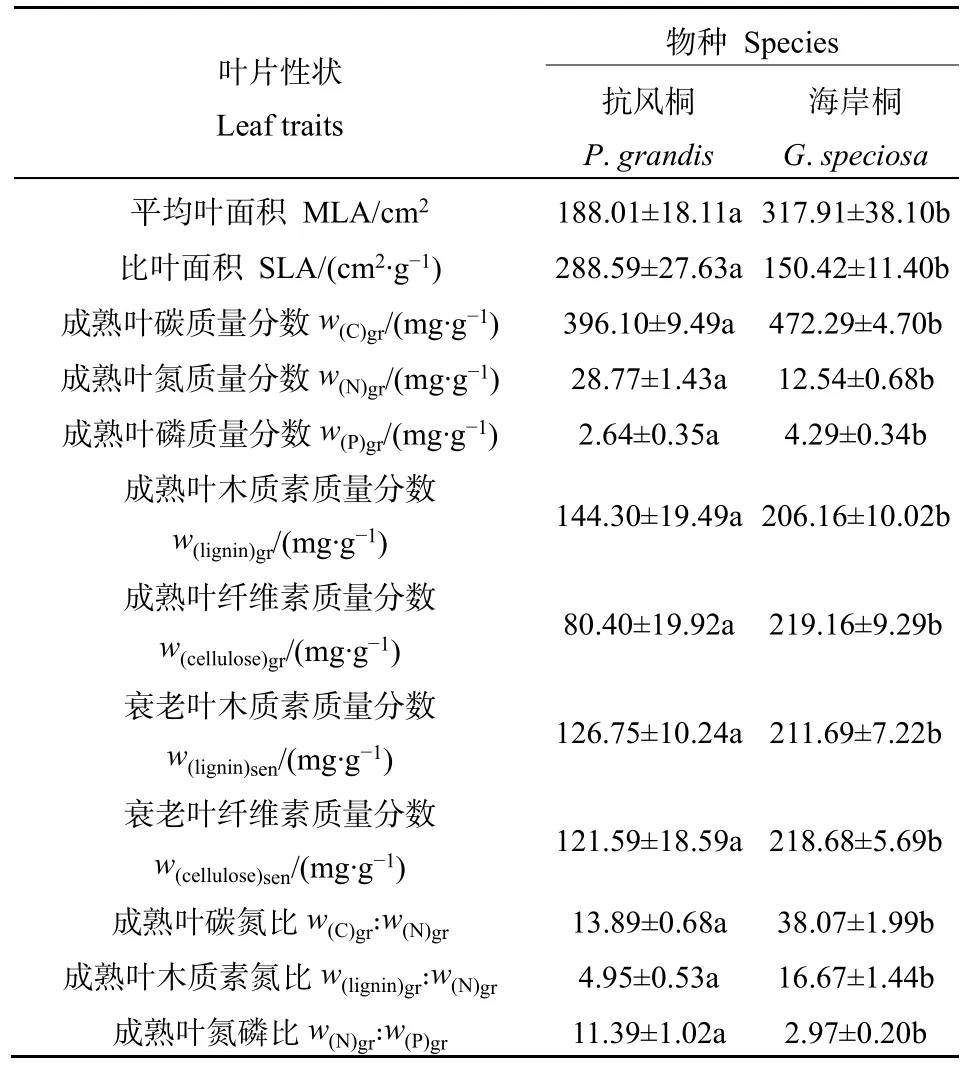
表2 抗风桐和海岸桐的叶片性状Table 2 Leaf traits of P. grandis and G. speciosa

图1 海岸桐和抗风桐叶片养分再吸收效率和养分再吸收程度Figure 1 Leaf nutrient resorption efficiency and proficiency of P. grandis and G. speciosa
2.3 抗风桐和海岸桐的叶片养分再吸收效率和养分再吸收程度
抗风桐和海岸桐的氮再吸收效率(NRE)之间没有显著差异(图1a),而抗风桐的磷再吸收效率(PRE)显著高于海岸桐(图1b)。海岸桐的氮再吸收效率显著高于磷再吸收效率,抗风桐的氮再吸收效率高于磷再吸收效率但不显著。抗风桐的氮磷再吸收效率比(NRE꞉PRE)显著低于海岸桐(图1c),两者的氮磷再吸收效率比均大于 1,说明两者倾向于再吸收更高比例的氮。另外,抗风桐的氮再吸收程度(NRP)显著高于海岸桐(图1d),而磷再吸收程度(PRP)则显著低于海岸桐(图1e),抗风桐的氮磷再吸收程度比(NRP꞉PRP)也显著高于海岸桐(图1f)。
2.4 养分再吸收与叶片性状和土壤因子之间的关系
Pearson相关性分析结果表明,绝大多数叶片性状与养分再吸收效率之间不存在显著相关关系。土壤化学性质对养分再吸收存在不同程度的影响,尤其是土壤养分含量的影响更大(表3)。为此,我们采用了线性回归模型来进一步分析养分再吸收效率与成熟叶养分含量和土壤养分含量之间的关系。结果表明,抗风桐和海岸桐的氮再吸收效率与叶片氮含量没有显著相关性,磷再吸收效率与叶片磷含量呈显著正相关(图2a,b)。抗风桐的氮再吸收效率与土壤全氮呈显著正相关,海岸桐的氮再吸收效率与有机质呈显著负相关(图2c,d)。抗风桐的磷再吸收效率与土壤全磷和有效磷之间均不存在显著相关关系,而海岸桐的磷再吸收效率与土壤全磷呈显著正相关,与土壤有效磷不存在显著相关关系(图2e,f)。

表3 抗风桐和海岸桐叶片养分再吸收效率与成熟叶片性状和土壤性质之间的相关性分析Table 3 Pearson correlations between leaf nutrient resorption efficiencies and mature leaf traits, soil properties in the P. grandis and G. speciosa forests
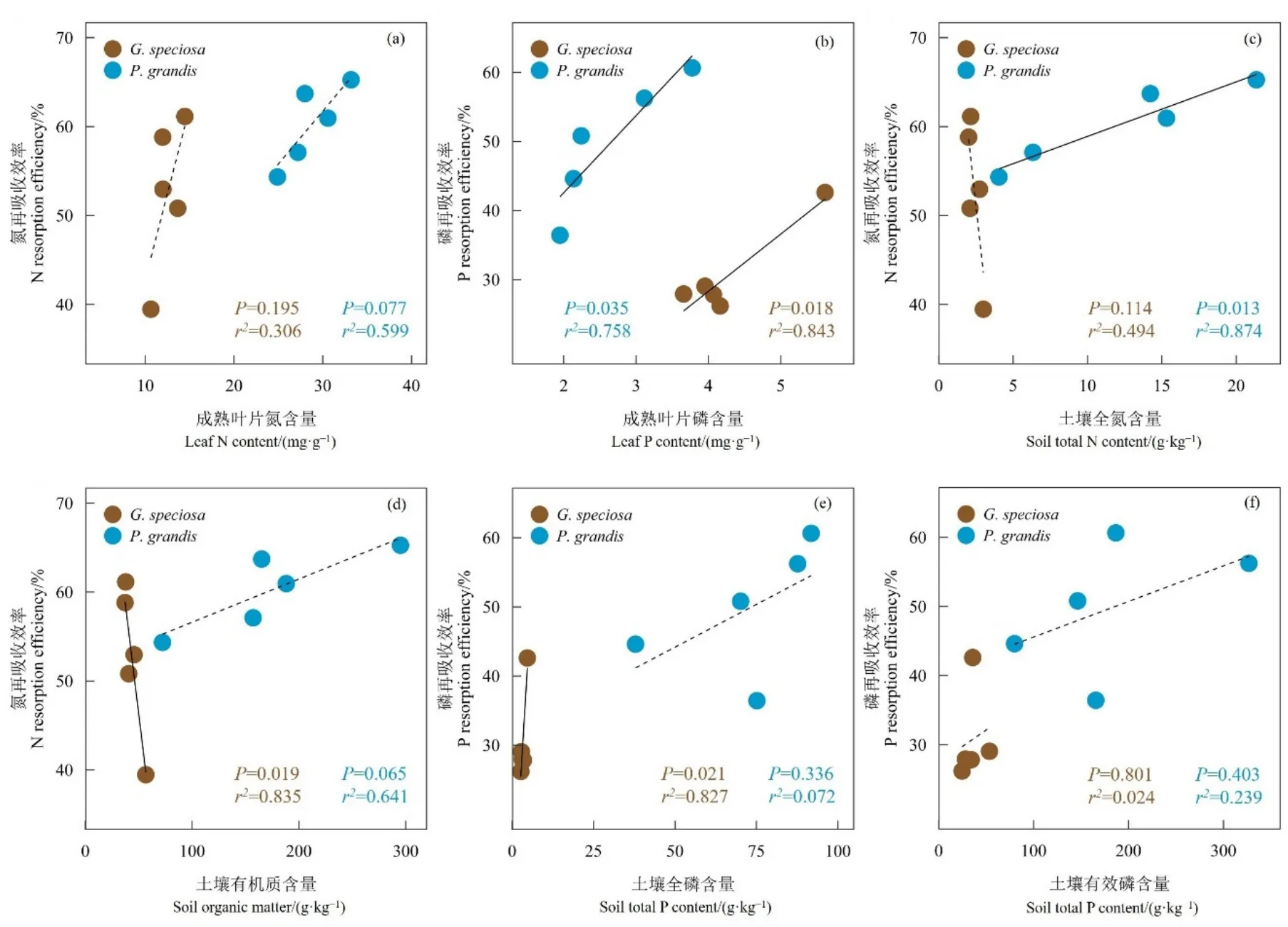
图2 抗风桐和海岸桐养分再吸收效率与叶片养分含量和土壤养分含量之间的线性回归分析Figure 2 Linear regression between leaf nutrient resorption and leaf nutrient content and soil nutrient content in the P. grandis and G. speciosa
3 讨论
3.1 热带珊瑚岛典型单优乔木树种的生态策略
西沙热带珊瑚岛抗风桐和海岸桐的叶片性状和养分再吸收特征存在较大差异,主要原因可能是这两个单优种林分间的土壤养分状况存在较大差异(表1)。抗风桐林的表层土壤有机质含量是海岸桐林的4倍,全氮、有效磷含量是海岸桐林的6倍,全磷含量更是达到了10倍以上。张宏达(1974)通过野外调查发现,海鸟更倾向于在高大郁闭的抗风桐林中栖息。海鸟是世界上许多沿海地区和海岛陆域生态系统的主要驱动力(Wardle et al.,2009),这些海鸟通过鸟粪的形式将大量的海洋资源输入至抗风桐林的土壤中,极大地提高了抗风桐林的土壤肥力。以往基于岛屿的研究表明,生长在海鸟群体附近的植物拥有更高的δ15N值、养分浓度和生产力,从而可能导致植物功能性状和植被功能组成发生重要转变(Maron et al.,2006)。植物为了最大化实现其对环境的适应性,通常会在叶片的各项功能性状之间进行权衡(Lohbeck et al.,2013)。本研究中,抗风桐的比叶面积和成熟叶氮含量显著高于海岸桐,而成熟叶碳含量、木质素含量、碳氮比以及木质素氮比显著低于海岸桐(表2),表明在适应海岛极端环境的过程中,海岸桐会将更多的生长资源分配给基于碳的防御化合物,以更好地适应珊瑚岛边缘相对贫瘠的土壤条件。而生长在珊瑚岛内部高土壤肥力条件下的抗风桐则会将更多的生长资源分配给光合蛋白等含氮化合物来增加光获取以支持其快速生长,这与Cornelissen et al.(2006)在瑞典北部的研究结果一致。植物的资源快速获取策略通常与较高的比叶面积、叶片氮含量相关,而高碳含量、木质素含量和碳氮比则反映了植物的资源保护策略。抗风桐在西沙热带珊瑚岛上的适应策略偏向于资源获取,而海岸桐则偏向于资源保护。
3.2 热带珊瑚岛典型单优乔木树种的养分再吸收特征
本研究中,抗风桐和海岸桐的成熟叶氮磷比均明显低于目前普遍认为的氮限制阈值(N꞉P<14;Koerselman et al.,1996),表明西沙热带珊瑚岛上这两种单优乔木树种的生长主要受到氮素限制。与近纬度但普遍受到磷限制的马来西亚(Tsujii et al.,2017)、斯里兰卡(Suriyagoda et al.,2018)、墨西哥(Renteria et al.,2011)、哥斯达黎加(Tully et al.,2013)和夏威夷(Cordell et al.,2001)等热带地区的植被相比,西沙抗风桐和海岸桐都表现出了相对较高的氮再吸收效率和较低的磷再吸收效率。其中,氮再吸收效率(60.28%和52.64%)高于Vergutz et al.(2012)报告的热带地区常绿乔木的平均值(49.5%),磷再吸收效率(分别为49.75%和30.73%)低于热带地区常绿乔木的平均值(59.3%)。这一现象可能与热带珊瑚岛的土壤年龄有关,因为长期的土壤成土作用会导致土壤养分有效性发生重要变化。例如,Laliberte et al.(2012)的研究证实了植物生产力会随着土壤年龄的增长从氮限制转变为磷限制,这说明热带珊瑚岛的土壤比绝大部分热带地区的风化土更为年轻,其生态系统的发展程度可能还停留在早期的氮限制阶段。抗风桐和海岸桐的叶片氮磷再吸收比均大于 1,表明热带珊瑚岛上的乔木种在受到氮限制时,会再吸收更多比例的氮,符合Han et al.(2013)提出的“相对再吸收假说”。因此,我们认为西沙热带珊瑚岛上的植物总体上表现出较为保守的氮利用策略。
海岸桐的叶片氮磷比显著低于抗风桐,说明海岸桐比抗风桐经历更为严重的氮限制。这可能是因为海鸟粪大量输入增加了土壤的氮含量,进而改善了抗风桐林的养分限制状况(王雪莹,2020)。然而,对新西兰的岛屿研究发现,海岛上的森林群落在鸟粪的影响下,其氮磷再吸收效率分别只有16.3%和11.0%(Wardle et al.,2009)。尽管该研究中的植被生长也受到了氮限制,但其养分再吸收效率明显低于本研究中的两个乔木种。一方面可能是因为单优种群落的养分需求高于共优种群落,另一方面也说明养分再吸收除了受土壤养分的调控外,可能还会受到当地环境条件的强烈影响(Agren et al.,2012)。植物可能会通过更高的养分再吸收来适应西沙珊瑚岛的恶劣环境,以维持其在岛上的生长优势。值得注意的是,在土壤养分含量明显更高的生境条件下,抗风桐的氮磷再吸收效率均高于海岸桐(尽管二者间氮再吸收效率差异不显著)。通常大多数单一优势树种都会发展出菌根以增加对土壤养分的获取(Connell et al.,1989),且菌根策略是植物在相对年轻的土壤中更为重要的策略之一(Lambers et al.,2008)。本研究中海岸桐可能将更多的生物量用于菌根的投资,使得其从土壤中获取养分的效率高于抗风桐(Newbery et al.,1997)。
养分再吸收程度是评估多年生植物养分再吸收中的重要指标(KillingBeck,1996;McGroddy et al.,2004;Yuan et al.,2009b)。本研究中,抗风桐和海岸桐的氮再吸收程度分别为 14.53 mg·g−1和7.53 mg·g−1,磷再吸收程度分别为1.63 mg·g−1和3.76 mg·g−1。参照 KillingBeck(1996)划分养分再吸收是否充分的标准,抗风桐对氮的再吸收不充分,而海岸桐较为充分的再吸收了叶片中的氮。这支持了我们的观点,即鸟粪的输入显著改善了抗风桐林的土壤养分条件及氮限制状态,使得抗风桐对氮的再吸收程度显著低于海岸桐。另外,抗风桐和海岸桐对于磷的再吸收均不充分(>0.5 mg·g−1),表明在富磷的珊瑚岛生态系统中,植物更倾向于从土壤中直接获取磷。根据Wright et al.(2003)的理论模型,在低肥力条件下,植物从衰老叶片中再吸收养分的能量成本低于直接从土壤中吸收养分的能量成本。从成本效益的角度来看,富磷生境下抗风桐和海岸桐从土壤中获取养分的能量成本低于养分再吸收。
3.3 叶片性状和土壤养分对养分再吸收的影响
抗风桐和海岸桐的磷再吸收效率均随着叶片磷含量的增加而显著增加(图2b),这与Kobe et al.(2005)和Vergutz et al.(2012)发现的叶片中养分含量的升高导致养分再吸收效率下降的研究结果不一致,可能与西沙热带珊瑚岛上的极端季节性干旱气候有关。抗风桐和海岸桐均具较强的耐旱性,本研究中的采样时间为雨季末期,在间歇性降雨的诱导下,这些耐旱植物会通过在一个生长季中产生多个叶片生产周期来适应干旱环境,进而导致养分再吸收效率出现较大的波动(Killingbeck,1992;Killingbeck,1993)。这种波动可能会掩盖植物叶片养分含量对养分再吸收的影响,未来仍需通过长期观测以深入了解西沙热带珊瑚岛植物养分再吸收动态特征。本研究发现,养分再吸收效率随着土壤养分含量的增加而增加,这与前人观察到的养分再吸收效率随土壤养分变化模式不一致(Aerts,1996;Achat et al.,2018)。养分再吸收作为植物的养分保存策略,其随着土壤养分的增加而降低这一推论理应具有逻辑性与正确性,但也有其他研究观察到与本文类似的现象(de Campos et al.,2013;See et al.,2015;Xu et al.,2020)。有趣的是,本研究中与养分再吸收效率呈显著正相关的土壤养分含量均显著高于Cleveland et al.(2007)报告中的全球平均值,比如抗风桐林的土壤全氮含量(12.25 mg·g−1vs 4.56 mg·g−1)以及海岸桐林的土壤全磷含量(3.3 mg·g−1vs 0.72 mg·g−1)。作为植物应对环境养分胁迫的关键策略,养分再吸收应受到影响环境养分供应的因素的影响(Tang et al.,2013),因此我们推断养分再吸收效率与土壤养分含量之间的正相关关系可能与鸟粪的输入密切相关。这一证据也表明,在高土壤肥力的生境中,土壤养分状况对植物的养分再吸收效率可能存在积极影响。然而,鸟粪在林下的分布通常具有随机性和不均匀性,可能导致了本研究中土壤养分与相应叶片养分再吸收效率之间关系的复杂性,相关机理仍需后续研究进一步探究。
4 结论
中国西沙群岛乔木植被代表性树种抗风桐和海岸桐表现出不同的生态策略,前者倾向于资源获取而后者则倾向于资源保护。与近纬度普遍受到磷限制的热带地区相比,西沙热带珊瑚岛的植物生长主要受到氮素的限制。抗风桐和海岸桐均表现出相对较高的氮再吸收效率和较低的磷再吸收效率,趋于较保守的氮利用策略。抗风桐和海岸桐衰老叶的磷再吸收程度均较高,对磷的再吸收不充分,在富磷珊瑚岛生态系统中更倾向于从土壤中直接获取磷。总之,抗风桐和海岸桐通过协调叶片资源分配、提高叶片氮再吸收效率和降低叶片磷再吸收效率以适应热带珊瑚岛的特殊生境。
