祁连山灌丛群落结构特征随海拔梯度和经度的变化
2022-04-16王小娜徐当会王谢军方向文
王小娜,徐当会,王谢军,方向文
兰州大学生态学院/草地农业生态系统国家重点实验室,甘肃 兰州 730000
灌丛通常是指高度在3 m以内、茎干具有多个分枝且主干不明显的木本植物。灌丛作为陆地生态系统中最重要的自然植被类型,在群落演替、生态系统碳固和生物多样性保育等方面扮演着不可替代的角色(Piaoet al.,2009;方精云等,2015),同时,在区域生态环境保护和替代能源方面也起着非常重要的作用(刘国华等,2003)。中国陆地生态系统中,灌丛分布广泛,面积为2×108hm2,中国是全球灌丛分布面积最广泛的国家(胡会峰等,2006),且近年来其分布面积有扩大的趋势(雷蕾,2012),但其易受海拔、气候、经纬度等环境因子的影响。其中,海拔梯度主要通过温度的变化影响灌丛群落分布格局,而经度则主要通过降雨变化影响灌丛群落(丁松爽等,2010;王金兰等,2019)。
祁连山位于中国青海省与甘肃省,是中国主要山系之一,祁连山国家公园是中国第一批国家公园试点,其天然物种资源丰富,其中,灌丛物种数可达乔木物种数的3倍。祁连山作为中国第一阶梯和第三阶梯的分界线(付建新等,2020),也是青藏高原、内蒙古高原和黄土高原的过渡区(齐鹏等,2015),由于其高纬度、高海拔、低气温的气候特征,被认为气候变化的敏感区和脆弱区(刘玉祯等,2019)。同时,也是中国西部地区重要的安全屏障和水源涵养区(雷蕾,2012;刘玉祯等,2019)。祁连山区灌丛物种丰富,分布面积大,约6.5×105hm2,约占林用面积的71.5%(梁倍等,2013),是森林生态系统的重要组成部分,在祁连山区具有非常重要的地位和作用(赵维俊等,2017)。由于人类活动和生态环境的恶化,祁连山生物多样性锐减趋势不断加剧,群落结构发生改变,因此,祁连山逐渐成为研究的热点地区(蔡蔚等,2020)。
目前国内外对灌丛群落结构的研究有大量报道(Evans et al.,1990;Xu et al.,2009)。王飞等(2018)对白龙江干旱河谷不同坡向主要灌丛群落随海拔梯度变化的物种多样性研究,发现海拔和坡向是影响生物多样性的主要因子;丁松爽等(2010)在研究黑河上游祁连山区植物群落随海拔生境的变化特征时发现,灌木物种丰富度随海拔升高呈现多峰分布格局;梁倍等(2013)对祁连山天涝池流域典型灌丛地上生物量沿海拔梯度变化规律的研究,发现灌丛地上生物量随海拔升高呈先增加后减小的单峰型曲线变化。综上发现,虽然对灌丛的研究已有大量报道,但以上报道仅限于同一个典型流域,对祁连山灌丛群落结构随海拔梯度和经度的变化却鲜有报道。因此,本研究以祁连山中东段灌木群落为研究对象,研究其群落结构随海拔梯度和经度的变化,旨在揭示祁连山灌丛群落结构随经度和海拔梯度的变化规律,对于祁连山地区的矿区修复以及植被恢复具有重要的指导意义。
1 研究地区与研究方法
1.1 研究区概况
研究区位于祁连山中东地段甘肃省境内(36°48′—38°22′N,100°37′—103°1′E),年均温度4.1—5.8 ℃,年均降水量322.8—354 mm(数据来源于中国气象数据网 http://data.cma.cn/site/index.html)。乔木建群种为青海云杉(Picea crassifolia),在阳坡有少量祁连圆柏林(Sabina przewalskii);灌木层主要优势种有银露梅(Potentilla glabra)、金露梅(Potentilla fruticosa)、山生柳(Salix oritrepha)、高山绣线菊(Spiraea alpina)、鬼箭锦鸡儿(Caragana jubata);草本层主要优势种有禾本科(Gramineae)、珠芽蓼(Polygonum viviparum)和嵩草(Kobresia myosuroides)(唐志红等,2020)。
1.2 野外观测样地的选择及样方设置
为了研究祁连山中东段灌丛群落结构沿经度的变化,本研究于2019年7—8月,从祁连山东段到中东段分别选择永登县奖俊埠林场(东段)、古浪县夏玛林场(中东)、民乐县海潮坝森林公园(中段),经度跨度约为2.5°(图1)。另外,为了研究同一经度灌丛群落结构随海拔梯度的变化,在以上各地点分别设置2700、2900、3100 m 3个海拔,本次野外调查共计27个灌丛样方(3经度×3海拔×3重复)。用GPS测定每个样地的经纬度,坡度仪测定样地坡度,罗盘仪测定样地坡向,样地信息见表1。在每个样地以对角线设置3个5 m×5 m的灌丛样方,并估量灌丛总盖度及各物种分盖度、测量灌木各物种的多度、高度、基径和冠幅并计算其地上生物量(雷蕾,2012)。

表1 样地基本信息Table 1 Basic information of sites

图1 观测样地空间位置Figure 1 The spatial position of the sample sites
灌丛群落结构变化用Shannon-wiener多样性指数、物种重要值、均匀度表示,其中物种重要值用来衡量各物种的优势度,各公式参照邓清月等(2019)的计算方法:

式中:
VI——物种重要值;
Rc——相对盖度;
Rh——相对高度;
Rf——相对频度;
Pc、Ph和Pf——样方内某物种的盖度、平均高度和频度;
Nc、Nh和Nf——样方内所有物种的盖度、平均高度和频度之和;
E——Pielou均匀度指数;
H——Shannon-wiener多样性指数;
S——所在样方中的物种数目;
Pi——物种i的相对重要值;
N——样方内出现的物种数。
1.3 数据处理
采用Excel 2010、SPSS 25.0将所调查的数据进行整理分析;采用Origin 2018和Sigmaplot 12.5软件作图。灌丛地上生物量、盖度、海拔高度的关系采用Pearson相关性分析;各个海拔高度和经度之间生物量以及盖度的差异采用One-way ANOVA(单因素方差分析)和LSD(多重比较)分析。
2 结果与分析
2.1 物种优势度、多样性和均匀度指数的变化
所有优势物种分布格局均为均匀型分布。金露梅、银露梅均匀分布在样地内,而高山绣线菊生长于金露梅灌丛中间,故也呈均匀型分布。同一经度,随着海拔高度的升高,灌丛林中银露梅的优势度逐渐下降甚至完全消失,金露梅的优势度增加;高山绣线菊在3个海拔梯度均有分布,且其优势度并没有显著变化;山生柳主要分布在海拔高度为2900 m以上,海拔高度为2700 m时,并未见山生柳,鲜黄小檗(Berberis diaphana)则相反,在海拔高度为3100 m时,未见其分布(表2)。Shannon-Wiener指数、Pielou均匀度在各段并未表现出相同的变化趋势。祁连山东段和中东段,随着海拔高度的升高,Shannon-Wiener多样性指数、Pielou均匀度增加;而在祁连山中段,前者呈递减趋势且差异显著(F=44.521,P<0.001),后者在3个海拔高度差异不显著(F=0.007,P=0.993;图2a、b)。
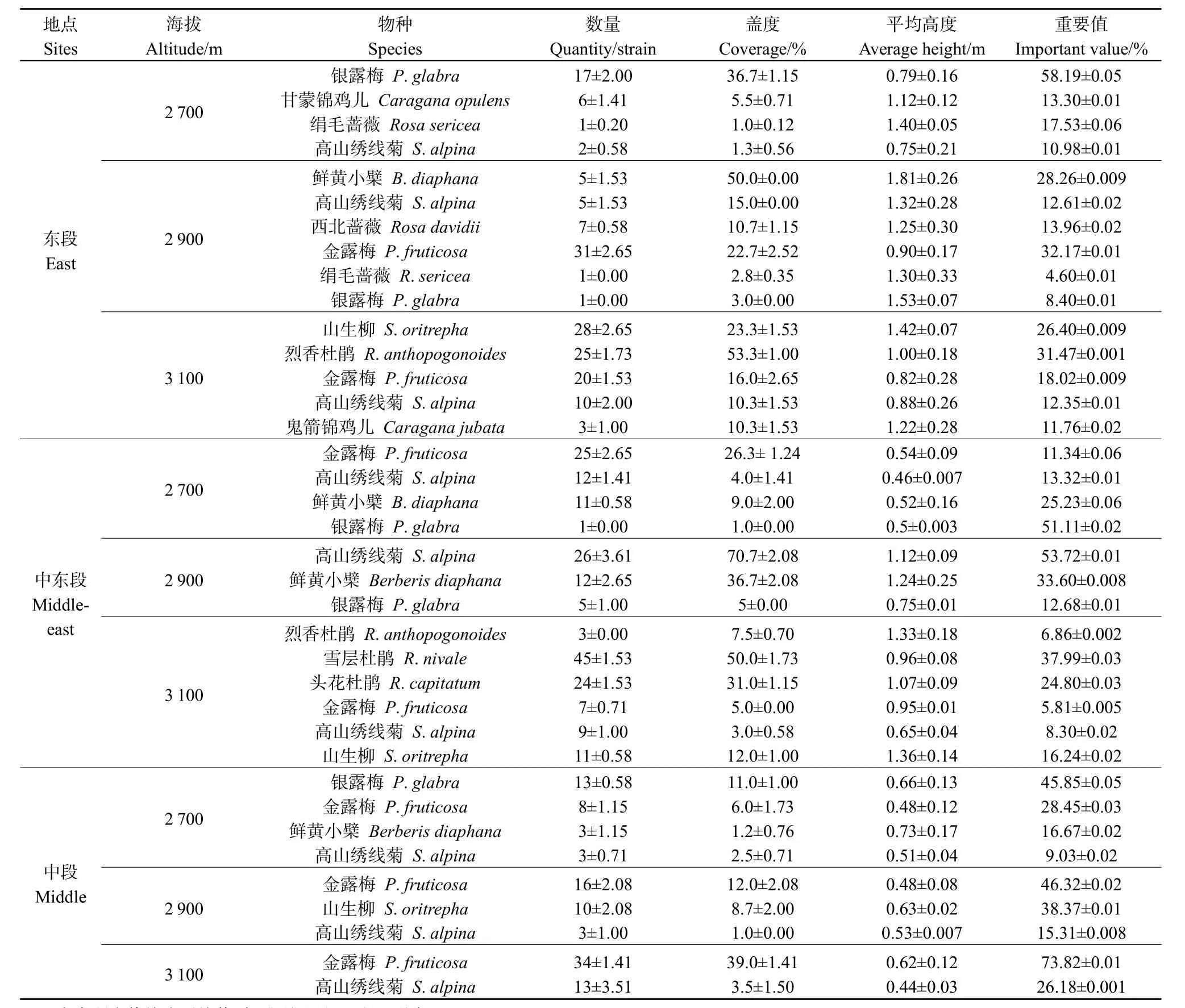
表2 不同海拔灌丛群落结构变化Table 2 The community structure changes of shrub at different elevations

图2 香农—威纳指数(a)、Pielou均匀度(b)、地上生物量(c)、盖度(d)随海拔梯度的变化Figure 2 Changes of Shannon-Wiener index (a), Pielou homogeneous degree (b), Biomass (c) and coverage (d)with altitude gradient
群落结构随经度(从西到东)的变化规律为:海拔为2900 m、3100 m时,Shannon-Wiener多样性指数随经度的增加而增加(F=275.049,P<0.001);海拔高度为2700 m时,随经度的增加而呈递减趋势(F=6.203,P=0.035;图2a)。在祁连山东段,山生柳、烈香杜鹃(Rhododendron anthopogonoides)和鬼箭锦鸡儿出现,优势种为山生柳、金露梅、烈香杜鹃、高山绣线菊;到中东段后,物种丰富度增加,雪层杜鹃(Rhododendron nivale)和头花杜鹃(Rhododendron capitatum)出现,并成为优势种;至中段只有金露梅和高山绣线菊两个物种(表2)。
2.2 地上生物量的变化
在同一经度,随着海拔高度的升高,地上生物量升高。海拔高度为2700 m和2900 m时,东段地上生物量差异不显著(F=31.719,P=0.076);中东段各个海拔之间地上生物量差异显著(F=261.862,P< 0.001);中段各个海拔之间地上生物量差异显著(F=1515.721,P<0.001)。在海拔高度为3100 m时,东段地上生物量最高,可达21454.1 kg·hm−2,约为海拔高度为2700 m时的4倍。在海拔高度为2700 m时,中段地上生物量最低,为175.73 kg·hm−2,仅占东段海拔高度为 3100 m时地上生物量的 1/122(图2c)。
在相同海拔高度,从西到东,随着经度的增加,地上生物量呈现递增的规律,即中段<中东段<东段(图2c)。海拔为2700m时,东段生物量为5701.33 kg·hm−2,约为中东段和中段生物量的2倍和32倍;海拔为2900 m时,东段生物量为10082.8 kg·hm−2,约为中东段和中段的1.5倍和17倍;海拔为3100 m时,东段生物量为21454.1 kg·hm−2,约为中东段和中段的2倍和8.5倍。
2.3 盖度的变化
盖度随海拔的变化为:随着海拔高度的升高,灌丛盖度升高。东段(F=438.311,P<0.001)和中东段(F=1056.692,P<0.001)在各个海拔之间盖度差异显著;中段海拔为2900 m时,与海拔为2700 m和3100 m相比,灌丛盖度差异不显著(F=5.094,P=0.051)。在海拔高度为2700—3100 m,东段盖度分别为42%、92%和99%;中东段盖度分别为39%、95%和99%;中段盖度为20%、26%和30%。在海拔为2700 m时,盖度下降明显,约为海拔高度为2900 m和3100 m时盖度的1/2(图2d)。
但在相同海拔高度,从西到东,随着经度的增加,盖度呈增加趋势。祁连山东段和中东段盖度差异不显著(F=33.555,P=0.37),但祁连山中段盖度与二者差异显著(F=656.032,P<0.001)。海拔为2700 m时,东段和中东段盖度分别为42%和39%,约为祁连山中段盖度的2倍;海拔为2900 m时,东段和中东段盖度分别为92%和95%,中段盖度为26%;海拔为3100 m时,东段和中东段盖度都为99%,中段盖度为30%(图2d)。
2.4 群落优势种高度随经度的变化
群落优势种高度随经度的变化规律如图3所示。金露梅、银露梅是主要优势种,其分布极广,从西到东都有分布。银露梅高度从西到东呈现下降趋势,且具有极显著性(r2=0.4344,P<0.001,图3a)。金露梅高度自西向东呈现上升趋势,且具有极显著性(r2=0.4641,P<0.001,图3b)。

图3 银露梅(a)、金露梅(b)高度随经度的变化Figure 3 The changes of height of P. glabra (a) and P. fruticosa (b) with longitude
2.5 海拔和经度对灌丛群落结构的影响
海拔和经度对地上生物量、Shannon-Wiener指数、Pielou均匀度、盖度双因素分析见表3。除经度对Shannon-Wiener指数影响不显著外,其余各项指标都受到海拔及经度和其交互作用的的影响,且影响显著。
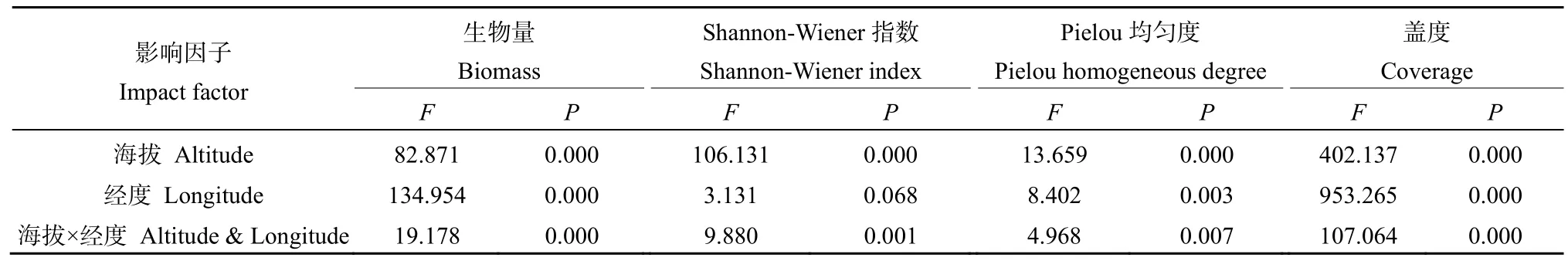
表3 海拔和经度对地上生物量、Shannon-Wiener指数、Pielou均匀度、盖度双因素分析Table 3 Two-factor analysis of elevation and longitude on aboveground biomass, Shannon-Wiener index, Pielou homogeneous degree and Coverage
2.6 灌丛地上生物量、Shannon–Wiener指数、Pielou均匀度、盖度、海拔高度相关性分析
对灌丛地上生物量、Shannon-Wiener指数、Pielou均匀度、盖度、海拔高度相关性分析见表4。除盖度与Pielou均匀度之间相关性不显著外,其余各项都存在显著的相关关系。海拔高度与灌丛地上生物量、Shannon-Wiener指数以及灌丛盖度存在极显著相关关系,相关系数分别为0.926、0.965、0.913,与Pielou均匀度相关性显著,相关系数为 0.762;Shannon-Wiener指数与地上生物量和灌丛盖度极显著相关,相关系数分别为0.943、0.872,与Pielou均匀度显著相关,相关系数为0.776;灌丛地上生物量与Pielou均匀度相关性极显著,相关系数为0.855,与灌丛盖度相关性显著,相关系数为0.740。

表4 灌丛生物量,Shannon–Wiener指数,Pielou均匀度,盖度,海拔相关性分析Table 4 Correlation analysis of shrub biomass, Shannon-Wiener index, Pielou homogeneous degree, Coverage and Altitude
3 讨论
3.1 海拔对物种组成的影响
由于海拔高度的不同,生物生境也存在很大差异,进而发生了随着海拔高度的变化,群落结构也发生改变的现象(丁松爽等,2010)。丁松爽等(2010)在研究黑河上游祁连山区植物群落随海拔生境的变化特征时发现,银露梅主要分布在海拔为2700—3000 m的阴坡,金露梅主要分布在海拔为3000—3300 m的阴坡,柳属灌木主要分布在中高海拔地区。占玉芳等(2015)在研究祁连圆柏群落物种多样性的海拔梯度效应时发现,海拔为2800 m时,银露梅为主要优势种,随着海拔高度的升高,金露梅逐渐出现,这与本研究的结果一致。本研究发现,随着海拔升高,银露梅优势度逐渐下降,金露梅优势度逐渐升高,高山绣线菊优势度随海拔梯度的变化不显著,这可能是由于银露梅、金露梅和高山绣线菊自身特性决定的。有研究表明,金露梅较银露梅有较强的抗胁迫能力,随着海拔高度的升高,温度降低,而金露梅耐寒性强,因此其便成为灌木群落的优势种,而银露梅由于竞争优势下降,逐渐被金露梅取代;而高山绣线菊在海拔 2000—4000 m的海拔处均有分布,主要原因是其耐寒、耐旱、适应性强(武龙庆等,2021;王晶等,2016)。
3.2 海拔对物种多样性的影响
对于物种多样性随海拔梯度的变化,虽然有大量报道,但未得出一致结论。目前,有5种变化模式,正相关、负相关、不相关,中度膨胀理论和中等海拔较低(温静等,2019),其中,中度膨胀理论被大量报道。本研究结果显示,随着海拔高度的升高,灌木物种多样性升高。邓清月等(2019)对晋西北饮马池山植物群落物种多样性沿海拔梯度的变化的研究中发现,灌木层α多样性指数呈单峰型变化趋势,且在2060 m处最大,这与本研究的结果存在一定差异;Rahbek(2005)的研究表明,随着海拔梯度的升高,物种多样性降低,这与本研究在祁连山中段的研究结果一致。形成这种结果的可能原因有两点:(1)研究区域的不同,造成了植物生境的不同,因此,物种多样性随海拔的变化模式也有所差异,本文通过Shannon-Wiener指数对海拔和经度的响应进行双因素分析(表3)也证实了这一假设;(2)海拔梯度的设置并没有达到物种多样性的峰值,因此物种多样性也不会呈单峰曲线模式。
3.3 海拔对灌丛生物量和盖度的影响
生物量是植物群落的重要指标,大部分学者通过以地上生物量来反映一个植物群落的生产力及生长状况(王金兰等,2019);植被盖度可以反映植物群落生长状况和生态环境质量,可以被用来评估生态系统的服务功能,同时也被用来估计土地退化和沙漠化程度(陈祖刚等,2014;王金兰等,2019)。生物量和盖度的大小受多方面因素的影响。一方面,受到群落自身状况的影响,包括物种组成、结构、植物多度、盖度、丰富度等;另一方面,还受到环境因子的作用,例如植物所生长的地形、地貌、海拔、坡向、土壤类型、温度和湿度等(唐志红等,2020),其中海拔被认为是最重要的影响因子(Gaston,2000)。目前,灌丛地上生物量随海拔的变化主要有3种形式,单峰曲线、正相关、负相关。而本研究结果显示,在海拔高度为 2700—3100 m范围内,灌丛地上生物量和盖度与海拔存在着显著的正相关关系,即随海拔高度的升高,灌丛地上生物量和盖度增加,这可能是由于在海拔高度为2700—3100 m范围内,降水量成为灌木生长的主要影响因子。随着海拔高度的升高,日照增强、降水量增加,为灌丛群落提供了较好的水热条件(Dorji et al.,2014),因此,灌丛长势良好,具有较高的盖度,同时盖度的增加也会导致灌丛地上生物量的增加,最终灌丛地上生物量和盖度表现为随着海拔的升高而升高。这与陈泓等(2007)、雷蕾(2011)、罗黎鸣等(2014)的研究结果一致。
3.4 经度对灌丛结构的影响
本研究发现,随着经度的增加(自西向东),灌丛地上生物量、盖度呈递增趋势。这可能是由于随着经度的增加,降雨量增加(邓少福,2013),从而使得各物种的生存环境优越,土壤水肥条件较好,由此使得各物种长势较好,最终导致生物量以及盖度的增加。另一方面,随着经度的增加(自西向东),灌木优势种银露梅和金露梅高度变化表现出相反的趋势,即银露梅高度降低,而金露梅高度上升,这可能是由这二者自身特点决定的。金露梅较银露梅更喜湿,因此其在祁连山东部长势更好,高度更高;而银露梅适应性强,耐旱性更强,对土壤肥力要求不高,因此较金露梅而言,其在祁连山中部其高度更高(王晶,2016)。
4 结论
通过对祁连山中东地段灌丛群落结构随海拔变化的研究发现,随着海拔高度的升高,群落结构发生变化,银露梅优势度降低,金露梅优势度升高;物种多样性随海拔梯度的变化表现为两种关系,即正相关和负相关;随着海拔高度的升高,灌丛地上生物量、盖度增加。随着经度的增加(自西向东),灌丛群落主要优势种银露梅和金露梅高度变化呈现出相反趋势,银露梅随着经度的增加,其高度逐渐降低,而金露梅高度则随着经度的增加而呈现递增趋势;灌丛群落生物量和盖度都呈现递增趋势。对祁连山灌丛群落结构、物种多样性、生物量、盖度随经度和海拔梯度变化的研究,可以为祁连山自然保护区的科学管理提供一定的理论指导,同时也可以为从植物、土壤理化性质、养分循环等方面研究祁连山自然保护区灌丛对海拔和经度的响应奠定基础。
