青海湖流域泥炭湿地地气系统不同时间尺度上CO2交换特征
2022-04-15左弟召陈克龙李英年张法伟
左弟召, 陈克龙, 李英年, 张法伟
青海湖流域泥炭湿地地气系统不同时间尺度上CO2交换特征
左弟召1,2,3, 陈克龙2,3,*, 李英年4, 张法伟4
1. 青海师范大学地理科学学院, 青海西宁 810008 2. 青海师范大学青藏高原地表过程与生态保育教育部重点实验室, 青海西宁 810008 3. 青海师范大学青海省自然地理与环境过程重点实验室, 青海西宁 810008 4. 中国科学院西北高原生物研究所, 青海西宁 810008
为了定量分析2017年青海湖流域泥炭湿地地气系统不同时间尺度上的碳交换的变化特征及影响机制, 利用涡动相关技术对其不同时间尺度上的碳通量进行了测定, 结果表明: 1)青海湖流域泥炭湿地地气系统在2017年表现为“碳源”, 全年合计排放209.312 gC·m–2。2)生态系统总初级生产量(GPP)和生态系统总呼吸(Re)年变化均呈倒V型, 而净生态系统碳交换(NEE)年内变化则呈双峰型。3)NEE和GPP 与各环境要素(气温、土壤温度、月平均降水量、土壤含水量)呈现负相关关系, 而Re与之呈显著的正相关关系(<0.01)。4)NEE受温度因子影响较大, 主要受控于气温。5)GPP和Re与各水热因子都有较大的相关性, 但GPP受温度因子影响较显著, 而水、热季节变化及其协调程度对Re有更大的影响。
泥炭湿地; 涡度相关; 碳通量; 青藏高原
0 前言
全球湿地面积约为5.7×106km2, 约占陆地面积的6%, 湿地生态系统土壤中有机碳含量约占全球碳含量的20%—25%[1]。观测湿地生态系统的碳通量是明确湿地碳源汇功能的重要手段, 也是了解陆地生态系统碳循环的重要环节。涡度相关技术(Eddy covariance, EC)是一种非破坏性的、能长期连续观测的微气象观测技术, 其优势在于研究不同时间尺度上生态系统碳收支的变化规律及其对环境因子的响应机理[2]。陆地生态系统碳循环已广泛使用涡度相关技术进行测定, 该技术渐渐成为生态系统水平测量碳通量的标准方法[3–4]。
自20世纪70年代以来, 有关陆地生态系统地表CO2通量的研究大多集中于森林、农田、草地、荒漠和盐沼湿地等生态系统[5–15]。高海拔地区的泥炭湿地不同时间尺度上量化碳收支及其环境要素响应机理却少有涉及。青藏高原被誉为世界“第三极”[16]。青藏高原东北部的青海湖高寒湿地位于西北干旱区及东部季风区的交汇处, 是高寒湿地生态系统的典型区域[17]。泥炭地储量了全球湿地绝大多数的碳, 且广布于北半球温带及寒冷地区[18]。因研究区域不同, 青藏高原不同的下垫面和泥炭层厚度呈现不同的碳源、碳汇潜能。如玉树隆宝湿地[19]、高寒草甸那曲站[20]和青藏高原东北部[21]表现为碳汇, 而高寒湿地生态系统(海北站)[22]和青海湖高寒藏嵩草湿草甸[23]则表现为碳源。为此, 有必要开展高寒泥炭湿地地气系统碳交换特征及其碳源汇机制的观测研究, 探究高海拔不同下垫面和泥炭厚度有何种碳源汇功能, 以期对高海拔地区泥炭湿地的碳通量的时空变异性和碳收支进行精确估算。
本文基于伊克乌兰瓦颜站2017年全年涡动相关观测数据, 通过定量分析青海湖流域高寒泥炭湿地碳通量的变化特征及其环境因子, 并明晰其碳收支在日变化和年变化尺度的变化规律, 进而探究高寒泥炭湿地有何种碳源汇功能?并对其环境因子影响机制如何?不仅对量化净生态系统碳交换(Net ecosystem exchange, NEE)、生态系统总呼吸(ecosystem respiration, Re)和生态系统总初级生产量(gross primary productivity, GPP)及其环境因子影响机制具有重要的意义, 而且为评估未来气候变化背景下青藏高原相关的高寒湿地生态系统的碳收支研究提供理论参考和数据支持。
1 研究区概况
研究区位于青海省海北藏族自治州刚察县伊克乌兰乡瓦颜山(37°44′35"N, 100°05′40"E), 北靠祁连山, 南临青海湖, 是沙柳河上游支流瓦颜曲的河源湿地。依托该区建设青海青海湖湿地生态系统国家定位观测研究站——伊克乌兰瓦颜站(图1)。该站点年均气温为(-3.06±0.40) ℃, 最高气温为33.3 ℃, 最低气温为-36.3 ℃。年均降水量为462.8 mm, 5—9月降水量约占年降水量的90%, 属于典型的高原大陆性气候[24], 主要特征为太阳辐射强, 气温低而日温差大, 降水少而集中, 雨热同期。该区海拔3750 m, 主要植被类型是高寒沼泽化草甸, 藏嵩草 ()是该区的优势种, 苔草鹅绒委陵菜为主要伴生种。主要土壤类型为沼泽土和草甸土, 土壤厚度约为1.2 m, 表层广布有厚度约1.7 m的季节性冻土层, 深层为永久性冻土[25]。

图1 研究区地理位置示意图
Figure 1 Geographical location of the study area
2 研究材料与方法
2.1 观测系统与观测要素
观测站的各观测要素和观测系统如下(表1), 是由安装于长期观测样地内每30 min记录一次的数据采集器(CR5000, Campbell Scientific, USA) 进行数据采集, 主要包括气温、降水量、总辐射和反射辐射等气象数据和不同梯度土层含水量数据。

表1 仪器及架设高度
2.2 数据解析与插补
选取了2017年1月1日至2017年12月31日涡度相关系统所观测的原始数据, 采用Eddypro5.0软件对其进行预处理, 包括坐标旋转、WPL 校正和光谱校正等[26–28]。由于LI-7500A型号分析仪存在表面自加热效应, 所以对1—5和10—12月气温低于-10 ℃时的数据处理时进行仪器自加热效应的校正[29–30]。为保证数据质量, 由仪器故障、微气象异常等因素造成观测数据出现的缺失或者“野点”进行剔除和插补; 剔除降水同期的观测数据, 降水对湍流平稳性有一定影响, 并有可能导致仪器失灵[27]; 针对夜间。(光合光量子通量密度PPFD<10 µmol·m–2·s–1)湍流发展不充分, 剔除夜间摩擦风速低于0.15 m·s–1对应的通量数据[31]
其中对非生长季全天以及生长季夜间采用Van't Hoff方程[32]进行插补:

式(1)中,为非生长季全天以及生长季夜间生态系

表2 缺失或丢失的通量数据插补法

对生长季白天缺失CO2通量数据, 利用Michaelis- Menten[33]方程进行插补:

涡度相关系统观测的CO2通量为NEE,将夜间呼吸函数外延至白天(Re, d), 即可得生态系统的总呼吸量(Re), 进而可得生态系统总交换量(GEE)[31]。在生态系统的尺度上, 生态系统总初级生产量(GPP)与GEE数值等同, 但方向相反。
2.3 数据分析
使用EDDY涡度软件来转换和处理碳通量数据。采用Excel 2013对碳通量、气象、土壤等数据作进一步计算处理。利用SPSS 18.0对数据进行相关分析与显著性检验。用Origin Pro 2018来拟合回归方程与制图。
3 结果与分析
3.1 微气象要素分布状况
研究时段内, 微气象要素在总体上呈现明显的季节变化特征(图2)。研究区气温(Ta)年均值为-3.06 ℃,月最高、最低气温分别为7月的9.3 ℃和1月的-14.9 ℃。5 cm深处的土壤温度(Ts)的月最高、最低分别出现在7月(10.36 ℃)和1月(-7.29 ℃)。Ts变化规律和Ta相似, 年平均Ts为1.49 ℃。降水总量(PPT)为540.258 mm, 主要集中在生长季。在5月和9月有两次脉冲性降水, 7月达到降雨量旺盛期。5—9月的降水总和为488.95 mm, 约占全年的90%。土壤含水量(SWC)季节变化与降水有相似的变化特征但有所滞后, 是由于植被的根系吸收水分, 导致土壤含水量不会迅速增加。光合光量子通量密度(PPFD)在7月达到峰值(744.01µmol·m–2·s–1), 夏季通量密度较大, 冬季通量密度较小。总辐射(DR)与PPFD的变化特征相似, 最高值出现在7月(374.553 W·m–2), 而反射辐射(UR)变化规律不同于其他, 全年总量较低且变化较平稳, 高值主要集中在4月和5月, 其中全年的最高值是5月3日(231.657 W·m–2), 之后便波动减小。

图2 2017年瓦颜山泥炭湿地气温和5 cm深处的土壤温度(a)、降水量(b)、土壤含水量(c)、光合光量子通量密度(d)、总辐射(e)、反射辐射(f)的季节变化
Figure 2 Seasonal variation of air temperature and soil temperature at 5 cm depths(a), precipitation (b), soil water content (c), photoperiod quantum flux density (d), total radiation (e), reflection radiation (f) in Wayanshan peat wetland in 2017
3.2 生态系统总初级生产量(GPP)在不同时间尺度上的变化特征
3.2.1 GPP日变化
根据CO2通量变化的基本形态将全年划分为生长季(5—9月)和非生长季(10—4月)[26]。生长季夜间GPP月平均日变化非常平稳(图3), 从日出后开始, GPP逐渐增加, 且减少速率随月份的不同其加快程度也不尽相同, 到北京时间11: 30左右半小时开始呈现波动减少的趋势, 持续到19: 00左右便归于平稳。主要是由于随着植物叶面积加大, 其光合效率、水分利用率逐渐增加, 生态系统的固碳能力随植物生长旺盛而增强。非生长季的植物处于无呼吸状态, 土壤表层受季节性冻土影响, 所以非生长季的GPP接近于0。
3.2.2 GPP年变化
2017年瓦颜山GPP总量约为246.470 gC·m–2。在5—9月生长季期间的日GPP年变化呈现先上升后下降的V字型(图4), 于第208日达到最高值4.318 gC·m–2·d–1, 非生长季的总量接近于0。整个生长季的总初级生产力具有明显的季节性, 在一年中水热条件最好的7、8月份最高, 其中7月>8月>6月> 9月>5月。
5月伊始, GPP较小且平稳; 6—7月是植物生长的旺盛时期, 总初级生产量逐渐增多; 8月开始逐渐减少, 直到9月(即生长季末期)变化趋势渐缓, 且总初级生产力降低, 非生长季则接近于0。
3.3 生态系统总呼吸(Re)在不同时间尺度上的变化特征
3.3.1 Re日变化
生态系统呼吸日变化十分平缓(图5), 夜晚呼吸值在全年都很小, 在北京时间8: 00左右达到日内最低值, 日间呼吸作用逐渐增强, 在14:30左右上升到日内最大值, 之后逐渐下降, 可见生态系统呼吸作用受温度影响最大。
1月的Re日变化最小(0.003—0.027 mgC·m–2·s–1), 5月开始其日变化趋势愈加显著(0.007—0.136 mgC·m–2·s–1), 4月和10月的变动水平相近(0.040—0.093 mgC·m–2·s–1), 主要归因于1月土壤温度极低, 水热条件差, 且土壤水分处于冻结状态, 微生物活动极弱, Re对土壤温度变化响应最不敏感。从5月持续到9月的植物生长季, 水热充足, 生态系统生物因素活跃, Re对土壤的温度变化响应最敏感, 4月和10月则处于中间状态, 但4月的温度低于10月。
3.3.2 Re年变化
2017年瓦颜山泥炭湿地生态系统总呼吸的变化总趋势为倒V型(图6), 在非生长季分布较为集中(0—1.0 gC·m–2·d–1), 冬季呼吸作用微弱且波动小。整个生长季分布相对分散, 7、8月份为呼吸作用最强的时期, 7、8月份呼吸最旺盛且基本在3 gC·m–2·d–1左右, 最大值为3.410 g(8月15日), 年内最低值为0.145 g(1月23日)。其中5月份和10月份有两次生态系统总呼吸次高峰, 可能是降水使草甸生态系统土壤微生物和植被根系的呼吸作用增强, 导致整个生态系统呼吸的增加, 植物返青期和枯草期的脉冲性降水会通过土壤含水量来间接促进草甸生态系统的碳排放, 成为影响碳收支的因素之一[23]。

图3 2017年瓦颜山沼泽湿地GPP生长季各月平均日变化
Figure 3 Monthly average diurnal variation of GPP during growth season in Wayanshan peat wetland 2017
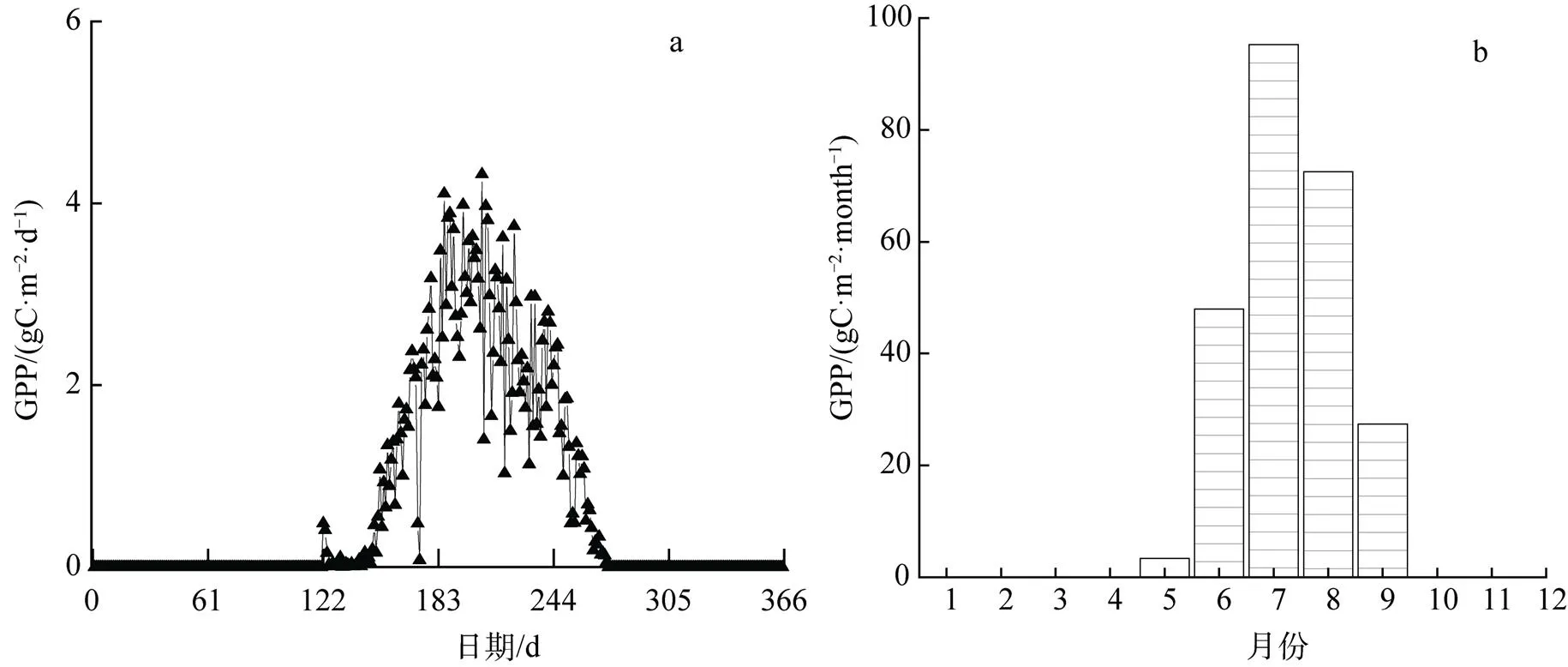
图4 2017年瓦颜山泥炭湿地日GPP年变化(a)和月总量年变化(b)
Figure 4 Daily GPP annual variation (a) and monthly total annual variation (b) for the Wayanshan peat wetland in the 2017
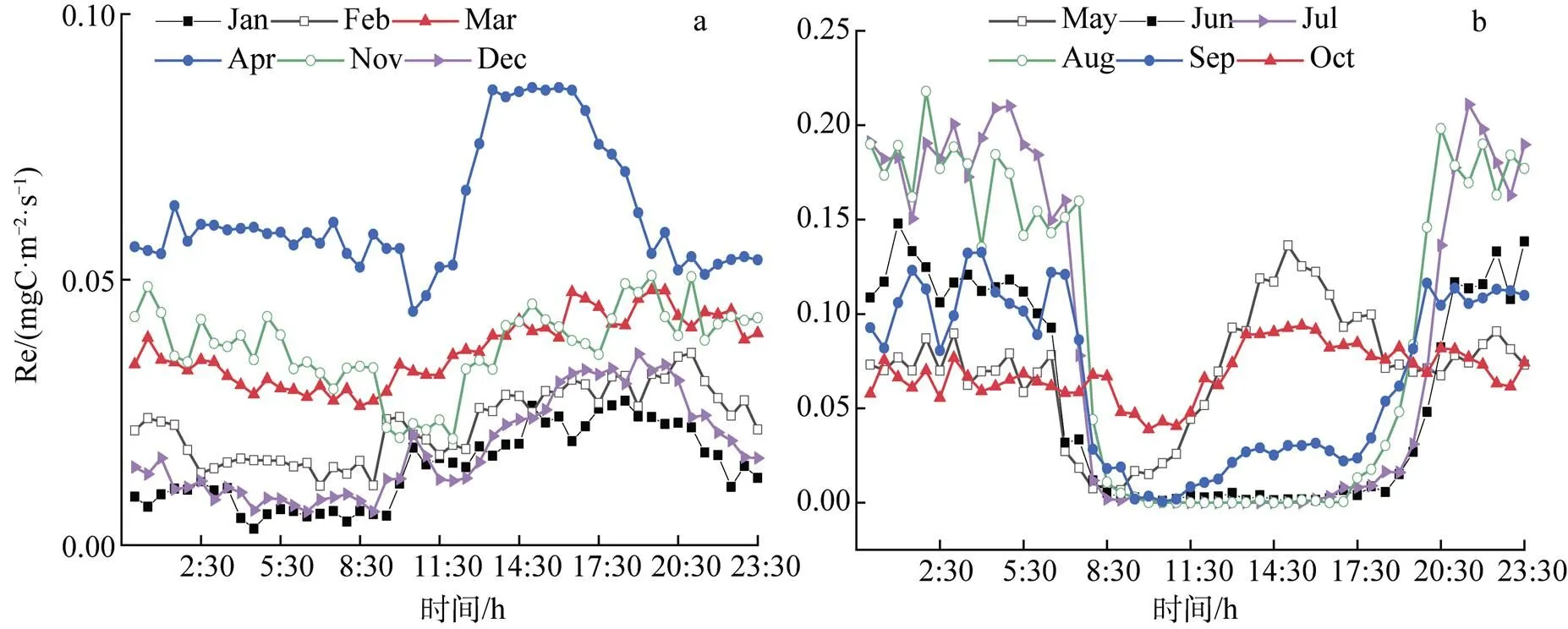
图5 2017年瓦颜山泥炭湿地Re非生长季(a)和生长季(b)各月平均日变化
Figure 5 Monthly average diurnal variation of Re during vegetation no-growing seasonal (a) and growing season (b) over the Wayanshan peat wetland in 2017
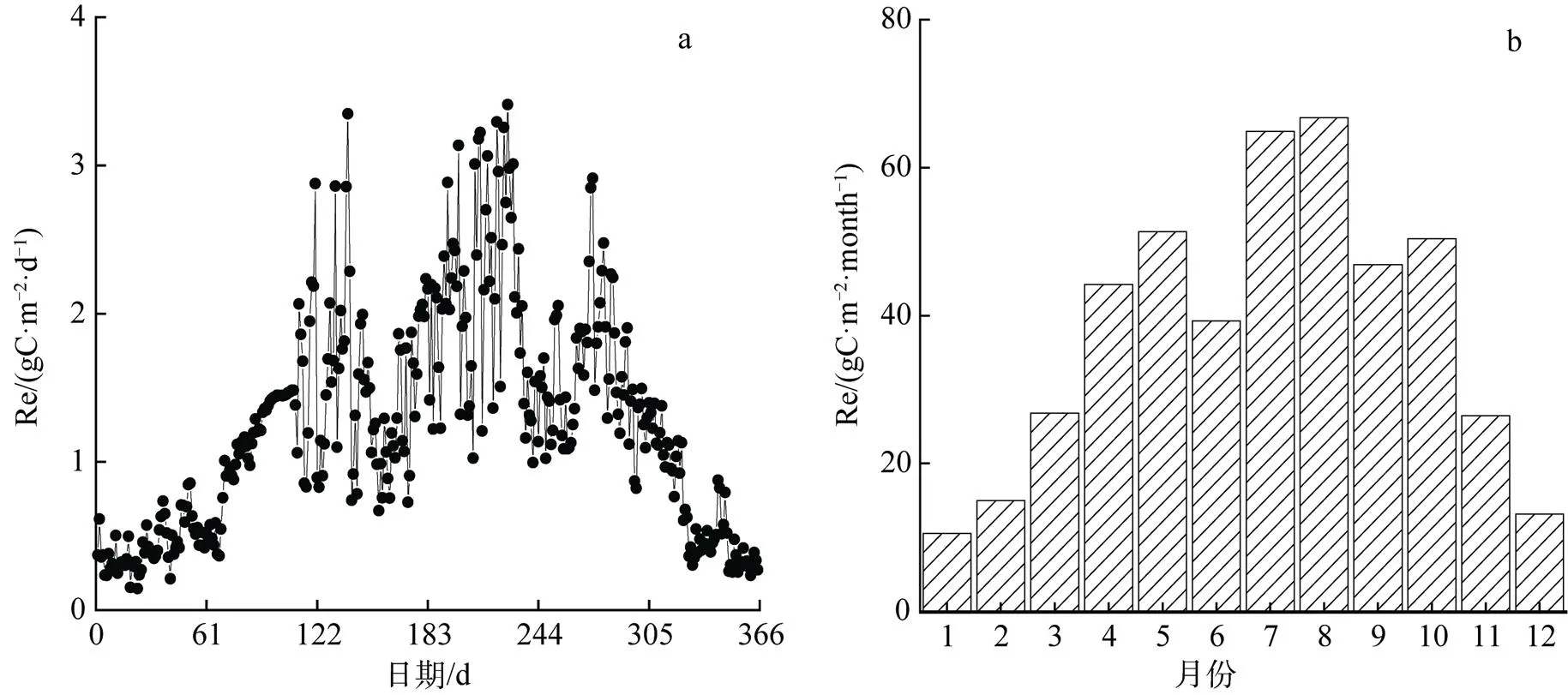
图6 2017年瓦颜山泥炭湿地日Re年内变化(a)和月总量年变化(b)
Figure 6 Daily Re annual variation (a) and monthly total annual variation (b) for the Wayanshan peat wetland in the 2017
2017年Re呼吸总量为455.781 gC·m–2。Re生长季总量为186.63 gC·m–2, 非生长季为269.145 gC·m–2。1月的月呼吸总量最少(约10.587 gC·m–2·month–1)。在生长季初期的5月, 太阳辐射较小, 气温较低, 地表大部分冻土尚未融解, 植物进入返青期, 但是呼吸作用仍较弱, Re维持在相对较低的水平。随着气温的回升, 地表的冻土逐渐融化, 植被的萌发和微生物活动渐强, 呼吸作用也愈强, 7、8月份的呼吸总量最大, 分别为64.893 gC·m–2·month–1和66.758 gC·m–2·month–1。此后由于太阳总辐射量的减少, 气温的降低及降水量的减少, 植被进入枯草期且微生物活动渐弱, Re于11月底下降到以土壤呼吸为主的冬季水平, 即使在极低气温和土壤温度下仍然存在一定的呼吸强度(0—1 gC·m–2·d–1)。
3.4 净生态系统碳交换(NEE)在不同时间尺度上的变化特征
3.4.1 NEE日变化
半小时NEE月平均日变化在非生长季变化较平稳且都有碳释放能力, 在0—0.5 mgC·m–2·s–1范围内变化, 只是不同月份释放的量大小不同(图7)。白天NEE碳吸收最高可达到1.095 mgC·m–2·s–1, 夜间生态系统则只有呼吸作用, 表现为CO2释放, 且维持在相对稳定的较低水平。由于青藏高原在非生长季气候寒冷, 低温抑制了土壤微生物的代谢活动, 再加上受到季节性冻土的影响, 导致该时段内生态系统碳通量整体较小[34]。
NEE在生长季(5月—9月)呈倒钟型, 7、8月份为生长季旺期, 生长季各月份平均日变形态基本一致, 夜间碳释放速率较低, 生长季的白天北京时间8: 00—18: 00期间NEE为负值, 表现为吸收状态, 其中7月>8月>6月>9月>5月。7:00之后由正转负, 在11: 30左右一个小时存在明显的低谷, 18: 00之后再由负转正, 但随着四季更替, 日照时间长短不同, NEE正负值转换的具体时间会不尽相同。
3.4.2 NEE年变化
日NEE年内变化呈双峰型(图8), 从第1日开始高寒湿地生态系统碳释放能力不断增强, 到第139日逐渐增大至第一个峰值(3.327 gC·m-2·d–1), 进入生长季后, 经过短暂的释放期, 日NEE由正转负并迅速下降, 在第208日降至最小值(-2.945 gC·m-2·d–1), 随后到第274日左右, 迅速上升并完成由吸收转外释放的转换过程, 随即到达第二个峰值(2.913 gC·m-2·d–1), 之后日NEE在小幅波动中略微下降并趋近于0值, 且波动幅度也越来越小。
NEE生长季总量为186.637 gC·m–2·month–1, 非生长季的总量为22.674 gC·m–2·month–1。5月开始由正转负, 6、7和8月为负值, 9开始由负转正。在5月和10月达到了NEE的高值, 分别为47.986 gC·m–2·month–1和50.400 gC·m–2·month–1。7月处于低谷期, 为-30.380 gC·m–2·month–1。

图7 2017年瓦颜山泥炭湿地NEE非生长季(a)与生长季(b)各月平均日变化
Figure 7 Monthly average diurnal variation of NEE during vegetation no-growing seasonal (a) and growing season (b) over the Wayanshan peat wetland in 2017

图8 2017年瓦颜山泥炭湿地日NEE年内变化(a)和月总量年变化(b)
Figure 8 Daily Re annual variation (a) and monthly total annual variation (b) for the Wayanshan peat wetland in the 2017
3.5 水热因子对生态系统碳通量影响机制分析
3.5.1 气温对NEE、Re、GPP的影响
通过对气温(Ta)和泥炭湿地生态系统碳通量各组分的相关性分析和线性拟合(图9), 结果表明: Ta与NEE呈负相关关系, 两者表现为二次多项式关系(P<0.01, 双尾检验)。Re与Ta的响应曲线呈显著的线性正相关(P<0.01, 双尾检验), GPP与Ta的关系可用指数方程表示, 且相关性极显著(P<0.01, 双尾检验)。GPP随着气温的增加而表现出生产力不断减小的趋势。
3.5.2 土壤温度对NEE、Re、GPP的影响
生态系统碳通量各组分均受控于Ts, 且与之有极显著的相关关系(P<0.01, 双尾检验)。可以明显看出Ts与NEE和GPP呈负相关, 均表现为二次多项式关系, 而Ts同Re呈显著线性正相关(图10)。

图9 2017年瓦颜山泥炭湿地气温(Ta)对NEE、Re、GPP的影响
Figure 9 Effect of air temperature (Ta) on the NEE, Re, and GPP in the Wayanshan peat wetland in 2017

图10 2017年瓦颜山泥炭湿地土壤温度(Ts)对NEE、Re、GPP的影响
Figure 10 Effect of soil temperature (Ts) on NEE, Re, GPP in Wayanshan peat wetland in 2017
3.5.3 月平均降水量对NEE、Re、GPP的影响
月平均降水量(PPT)与高寒泥炭湿地碳通量也存在显著的相关关系(P<0.01, 双尾检验), 其中, NEE和GPP与PPT呈现负相关, 与PPT的响应曲线分别为三次多项式和线性方程, 且GPP有更高的线性相关性(图11)。土壤呼吸随季节降水量的变化而一般呈正相关关系[35]。而本文Re与PPT的拟合结果也很好地印证了这结论。
3.5.4 SWC对NEE、Re、GPP的影响
地下5cm土壤含水量(SWC)和泥炭湿地生态系统碳通量各组分均有显著的相关关系(P<0.01, 双尾检验)。NEE和GPP与SWC呈负相关, 均表现为二次多项式关系(图11), Re与SWC则为显著的线性正相关。
4 讨论
4.1 泥炭湿地与其他生态系统碳通量特征的比较
青海湖流域泥炭湿地地气系统在2017年表现为“碳源”, NEE年总量为209.312 gC·m–2, 较藏北高寒灌丛[36]的66.75 gC·m–2高, 但较北美混合高草草原[37]的361.90 gC·m–2低。可能由于不同下垫面和泥炭层厚度, 土壤有机碳含量不同, 使得植物的碳源潜力不同[38]。由于研究地位于纬度靠北、海拔较高的河源, 山区气温日较差较大, 泥炭湿地NEE最大吸收速率均出现在11: 30左右, 最高可达1.095 mgC·m–2·s–1晚于半干旱高寒草原生态系统(拉萨班戈)的9:00和高寒草甸生态系统(云南丽江)的11:00[39]。
4.2 水热因子对NEE、Re、GPP的影响机制分析
泥炭湿地的形成和发展是一个复杂而漫长的过程, 是各种自然要素综合作用的结果, 而水、热季节变化及其协调程度是影响生态系统碳通量大小和季节变异的主要环境因子[40]。结果表明NEE和GPP 与部分环境要素(气温、土壤温度、月平均降水量、土壤含水量)呈现相似的相关性, Re则与其相关性则呈现相反的趋势, 这与海北高寒灌丛[41]和高寒湿地生态系统[22]的结论相似。NEE受温度因子影响较大, 主要受控于Ta。这与青海湖北岸高寒草甸草原[31]研究结果一致。随着气温的升高, 泥炭湿地生态系统NEE值降低, 可能由于气温的不断升高, 导致光合作用和呼吸作用的效率同时提高, 但呼吸作用的效率提速要高于呼吸作用, 使得NEE减少, 泥炭湿地生态系统的固碳作用不断增强[42]。Ta、Ts、PPT和SWC都对Re有显著影响。由于土壤温度的增加, 促进了植物的生长速率和土壤微生物活性, 使得生态系统总呼吸不断增多。降水可以通过影响土壤中生物活动和根系生长所需要的水量、土壤含水量以及土壤温度来影响土壤呼吸, 降水的发生使得土壤含水量的增加, 从而增加植物根系呼吸和增强土壤微生物的活性, 导致土壤呼吸释放的碳通量也会迅速升高[34]。GPP主要受Ta、Ts和PPT的影响, 表明在温度是主要驱动因素的生态系统中, 降水的季节分配对GPP的影响也较为明显。

图11 2017年瓦颜山泥炭湿地月平均降水量(PPT)对NEE、Re、GPP的影响
Figure 11 Effect of monthly average precipitation (PPT) on NEE, Re, GPP in Wayanshan peat wetland in 2017

图12 2017年瓦颜山泥炭湿地5cm深处土壤含水量(SWC)对NEE、Re、GPP的影响
Figure 12 Effect of soil water content (SWC) on NEE, Re, GPP in Wayanshan peat wetland in 2017
5 结论
(1)青海湖流域泥炭湿地地气系统在2017年表现为“碳源”, 全年合计排放209.312 gC·m–2, 其碳通量各组分在不同时间尺度上具有明显的日变化和年变化特征。GPP年变化呈倒V型, Re年变化总趋势为倒V型, 而日NEE年内变化则呈双峰型。生态系统碳通量在生长季都处于交换活跃时段, 在非生长季处于平稳状态。
(2)对泥炭湿地生态系统的环境控制因子进行相关分析, NEE和GPP与各水热因子(气温、土壤温度、月平均降水量、土壤含水量)呈负相关关系。其中, NEE受温度因子影响较大, 主要受控于气温; GPP与各水热因子都有较大的相关性, 温度因子影响较显著。Re则与各水热因子呈显著的正相关, 水、热季节变化及其协调程度对Re有更大的影响。因此, 应该进一步探究水热因子对的各自贡献多少?以及研究水热协调度对湿地生态系统碳通量有何种的影响机制。
[1] GORHAM E. The biogeochemistry of northern peatlands and its possible responses to global warming[C]// Woodwell G M, Mackenzie F T. Biotic Feedbacks in the Global Climate System: Will the Warming Feed the Warming? New York, Oxford University Press, 1995: 169–187.
[2] CIAIS P, REICHSTEIN M, NIOVY N, et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003[J]. Nature, 2005, 437(7058): 529–533.
[3] FALGE E, TENHUNEN J, BALDOCCHI D, et al. Phase and amplitude of ecosystem carbon release and uptake potentials as derived from FLUXNET measurements[J]. Agricultural and Forest Meteorology, 2002, 113(1): 75–95.
[4] Gilmanov T G, Soussana J F, Aires L, et al. Partitioning European grassland net ecosystem CO2exchange into gross primary productivity and ecosystem respiration using light response function analysis[J]. Agriculture, Ecosystems and Environment, 2006, 121(1): 93–120.
[5] 同小娟, 张劲松, 孟平. 基于涡度相关法的森林生态系统碳交换及其控制机制[J]. 温带林业研究, 2018, 1(2): 1– 9.
[6] 田容才, 文双雅, 阳会兵. 基于涡度相关法的农田生态系统碳通量研究进展[J]. 激光生物学报, 2019, 28(5): 415–420.
[7] 王杰, 叶柏生, 张世强, 等. 祁连山疏勒河上游高寒草甸CO2通量变化特征[J]. 冰川冻土, 2011, 33(3): 646–653.
[8] 刘冉, 李彦, 王勤学, 等. 盐生荒漠生态系统二氧化碳通量的年内、年际变异特征[J]. 中国沙漠, 2011, 31(1): 108– 114.
[9] 张丽华, 陈亚宁, 李卫红, 等. 准噶尔盆地梭梭群落下土壤 CO2释放规律及其影响因子的研究[J]. 中国沙漠, 2007, 27(2): 266–272.
[10] 李凌浩, 王其兵, 白永飞, 等. 锡林河流域羊草草原群落土壤呼吸及其影响因子的研究[J]. 植物生态学报, 2000, 24(6): 680–686.
[11] 王广帅, 杨晓霞, 任飞, 等. 青藏高原高寒草甸非生长季温室气体排放特征及其年度贡献[J]. 生态学杂志, 2013, 32(8): 1994–2001.
[12] 于贵瑞. 全球变化与陆地生态系统碳循环和碳蓄积[M]. 北京: 气象出版社, 2003.
[13] 王雷, 刘辉志, Schaffrath D, 等. 内蒙古羊草和大针茅草原下垫面水汽、CO2通量输送特征[J]. 高原气象, 2010, 29(3): 605–613.
[14] 乔春连, 李婧梅, 王基恒, 等. 青藏高原高寒草甸生态系统CO2通量研究进展[J]. 草业科学, 2012, 29(2): 204– 210.
[15] 贺文君. 潮汐作用对黄河三角洲盐沼湿地生态系统CO2和CH4交换的影响[D]. 烟台: 中国科学院烟台海岸带研究所, 2018.
[16] 闫巍, 张宪洲, 石培礼, 等. 青藏高原高寒草甸生态系统CO2通量及其水分利用效率特征[J]. 自然资源学报, 2006, 21(5): 756–767.
[17] 王记明, 陈克龙, 曹生奎, 等. 青海湖高寒湿地生态系统夏季CO2通量日变化及其影响因子研究[J]. 生态与农村环境学报, 2014, 30(3): 317–323.
[18] EINOLAPPALAINE N. Global Peat Resources[M]. Finland: International Peat Society of Finland, 1996.
[19] 张海宏, 李林, 周秉荣, 肖宏斌. 青藏高原高寒湿地CO2通量特征及影响因子分析[J]. 冰川冻土, 2017, 39(1): 54–60.
[20] 朱志鹍, 马耀明, 胡泽勇, 等. 青藏高原那曲高寒草甸生态系统CO2净交换及其影响因子[J]. 高原气象, 2015, 34(5): 1217–1223.
[21] SUN Shaobo, CHE Tao, LI Hongyi, et al. Water and carbon dioxide exchange of an alpine meadow ecosystem in the northeastern Tibetan Plateau is energy-limited[J]. Agricultural and Forest Meteorology, 2019, 283–295.
[22] 张法伟, 刘安花, 李英年, 等. 青藏高原高寒湿地生态系统CO2通量[J]. 生态学报, 2008, 28, (2): 453–462.
[23] 吴方涛, 曹生奎, 曹广超, 等. 青海湖高寒藏嵩草湿草甸湿地生态系统CO2通量变化特征[J]. 生态与农村环境学报, 2018, 34(2): 124–131.
[24] 干珠扎布. 模拟气候变化对高寒草地物候期、生产力和碳收支的影响[D]. 北京: 中国农业科学院, 2017.
[25] 张乐乐, 高黎明, 陈克龙. 青海湖流域瓦颜山湿地辐射平衡和地表反照率变化特征[J]. 冰川冻土, 2018, 40(6): 1216–1222.
[26] 王江涛, 周剑虹, 欧强, 等. 崇明东滩滨海围垦湿地CO2通量贡献区分析[J]. 生态与农村环境学报, 2014, 30(5): 588–594.
[27] 李春, 何洪林, 刘敏, 等. China FLUX CO2通量数据处理系统与应用[J]. 地球信息科学, 2008(5): 557–565.
[28] WEBB E K, PEARMAN G I, LEUNING R. Correction of Flux Measurements for Density Effects Due to Heat and Water Vapor Transfer[J]. Quarterly Journal of the Royal Meteorological Society, 1980, 106(447): 85–100.
[29] 朱先进, 于贵瑞, 王秋凤, 等. 仪器的加热效应校正对生态系统碳水通量估算的影响[J]. 生态学杂志, 2012, 31(2): 487–493.
[30] 吉喜斌, 赵文智, 康尔泗, 等. 仪器表面加热效应对临泽站开路涡动相关系统CO2通量的影响[J]. 高原气象, 2013, 32(1): 65–77.
[31] 张法伟, 李英年, 曹广民, 李凤霞, 等. 青海湖北岸高寒草甸草原生态系统CO2通量特征及其驱动因子[J]. 植物生态学报, 2012, 36(3): 187–198.
[32] LLOYD J, TAYLOR J A. On the Temperature Dependence of Soil Respiration[J]. Functional Ecology, 1994, 8(3): 315–323.
[33] FALGE E, BALDOCCHI D, OLSON R, et al. Gap Filling Strategies for Defensible Annual Sums of Net Ecosystem Exchang[J]. Agricultural and Forest Meteorology, 2001, 107(1): 43–69.
[34] 岳广阳, 赵林, 赵拥华, 等. 青藏高原草地生态系统碳通量研究进展[J]. 冰川冻土, 2010, 32(1): 166–174.
[35] 徐世晓, 赵亮, 李英年, 等. 降水对青藏高原高寒灌丛冷季CO2通量的影响[J]. 水土保持学报, 2007(3): 193– 195.
[36] ZHAO Liang, LI Yingnian, XU Shixiao, et al. Diurnal, seasonal and annual variation in net ecosystem CO2exchange of an alpine shrubland on Qinghai-Tibetan plateau[J]. Global Change Biology, 2006, 12(10), 1940– 1953.
[37] LAW B E, FALGE E, GU L, et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation[J]. Agricultural and Forest Meteorology, 2002, 113(1): 97–120.
[38] 蔡倩倩. 若尔盖高寒嵩草草甸湿地土壤碳储量研究[D]. 北京: 中国林业科学研究院, 2012.
[39] WANG Lei, LIU Huizhi, SHAO Yaping, et al. Water and CO2 fluxes over semiarid alpine steppe and humid alpine meadow ecosystems on the Tibetan Plateau[J]. Theoretical Applied Climatology, 2018, 131: 547–556.
[40] 柴曦, 李英年, 段呈, 等. 青藏高原高寒灌丛草甸和草原化草甸CO2通量动态及其限制因子[J]. 植物生态学报, 2018, 42(1): 6–19.
[41] 徐世晓, 赵亮, 赵新全, 等. 青藏高原高寒灌丛非生长季节CO2通量特征[J]. 西北植物学报, 2006, 26, (12): 2528– 2532.
[42] 吴灏, 叶柏生, 吴锦奎, 等. 疏勒河上游高寒草甸生态系统CO2通量观测研究[J]. 草业学报, 2013, 22(4): 18–26.
Characteristics of CO2exchange on different time scales of peat wetland land-atmosphere system in Qinghai Lake basin
ZUO Dizhao1,2,3, CHEN Kelong2,3,*, LI Yingnian4, ZHANG Fawei4
1. College of Geographical Sciences, Qinghai Normal University, Xining 810008, China 2. MOE Key Laboratory of Tibet Plateau Land Surface Process and Ecological Conservation, Xining 810008, China 3. Qinghai Province Key Laboratory of Physical Geography and Environmental Process, Xining 810008, China 4. Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, China
In order to quantitatively analyze the carbon exchange change characteristics and impact mechanism of the land-atmosphere system of peat wetland on the Qinghai Lake basin at different time scales in 2017, this paper used eddy covariance technique to measure its carbon flux on different time scales. The results showed that: 1) The land-atmosphere system of peat wetland in Qinghai Lake Basin behaved as a "carbon source" in 2017, with a total annual emission of 209.312 gC·m-2.2) The annual variation of gross primary productivity (GPP) and ecosystem respiration (Re)of the system were both inverted V type, while the annual variation of net ecosystem exchange(NEE) was bimodal pattern. 3) NEE and GPP had a negative correlation with various environmental factors (air temperature, soil temperature, monthly average precipitation, and soil water content), but Re had a significant positive correlation with them (<0.01). 4) NEE was greatly affected by temperature factors and was mainly controlled by air temperature. 5) Both GPP and Re had greater correlations with various hydrothermal factors, but GPP was significantly affected by temperature factors, while the seasonal changes in water and heat and their coordination degree had a greater impact on Re.
peat wetland;eddy covariance;CO2fluxes;Qinghai-Tibetan Plateau
10.14108/j.cnki.1008-8873.2022.02.021
S157.2
A
1008-8873(2022)02-174-10
2020-03-14;
2020-04-22
国家重点研发计划项目(2017YFC0403601); 国家自然科学基金项目(41661023); 青海省科技厅项目(2017-ZJ-782; 2018-ZJ-T09)
左弟召(1996—), 男, 四川资阳人, 硕士研究生, 从事生物地理与自然保护研究, E-mail: 17808324634@163.com
通信作者:陈克龙(1965—),男, 安徽芜湖人, 教授, 博士生导师, 主要从事生物地理与湿地生态研究, E-mail: ckl7813@163.com
左弟召, 陈克龙, 李英年, 等. 青海湖流域泥炭湿地地气系统不同时间尺度上CO2交换特征[J]. 生态科学, 2022, 41(2): 174–183.
ZUO Dizhao, CHEN Kelong, LI Yingnian, et al. Characteristics of CO2exchange on different time scales of peat wetland land- atmosphere system in Qinghai Lake basin[J]. Ecological Science, 2022, 41(2): 174–183.
