樟树、阴香种子萌发及幼苗生长研究
2022-04-15董斌黄永芳张晖张祥会李荣喜李孟南洪文泓刘文
董斌 黄永芳 张晖 张祥会 李荣喜 李孟南 洪文泓 刘文
摘要:樟树(Cinnamomum camphora)和阴香(Cinnamomum burmannii)是樟科优质的木本植物,生产中以播种繁殖为主。为探索出樟树和阴香高效苗圃育苗技术,对其苗圃催芽、萌发营养物质变化以及一年生幼苗生长规律开展系统研究。结果表明,樟树种子和阴香种子苗圃播种的最优组合条件为土壤基质、冬播、50 ℃浸种处理。樟树种子和阴香种子的可溶性糖、粗蛋白质和脂肪含量均较高,在萌发过程中,3种营养成分整体呈下降趋势。相反地,2种种子初始淀粉含量均较低,但是在萌发过程中持续升高。樟树一年生冬播苗各项生长指标均优于春播苗,其最佳播种育苗期是冬季。阴香在冬季或者翌年春季播种育苗均可行,一年生冬播苗和春播苗生长指标差异不显著。
关键词:樟树;阴香;种子;发芽;苗木
中图分类号:S718.43 文献标志码: A
文章编号:1002-1302(2022)06-0131-06
收稿日期:2021-05-21
基金项目:广东省高校重点科研平台和项目(编号:2020GCZX009,2019KZDZX2035);广东省林业科技创新项目(编号:2017KJCX005);广东农工商职业技术学院热带作物应用技术协同创新中心建设项目(编号:XJZX1902);2020年度广东省教育厅高职教育教学改革研究与实践项目(编号:JGGZKZ2020063)。
作者简介:董 斌(1982—),男,广东广州人,博士,副教授,研究方向为城市绿化及作物学。E-mail:bbeenn@163.com。
通信作者:黄永芳,教授,硕士生导师,研究方向为经济林及森林培育。E-mail:hyfang@scau.edu.cn。
樟树(Cinnamomum camphora)和阴香(Cinnamomum burmannii)是樟科应用广泛的优质木本植物,因其生态、用材、药用、油用等诸多应用价值。目前香樟和阴香已成为我国南方重点发展的乡土阔叶树种,作为优良的园林绿化树种或工业用材等材料进行模范化开发的前景非常广阔,市场对其苗木需求量也非常巨大。针对这2种优良树种,科技工作者开展了多方面的研究,近期有环境适应与生态效益[1-2],造林密度与用材生产[3-4],精油提取与精深加工[5-6]等研究成果见报,进一步加深了对这2种树种的认识。
在诸多的城市绿化苗木来源中,苗圃育苗占据着绿化苗木供应量的90%以上,是主要的苗木繁育途径。樟树和阴香可采取播種、扦插、组培等多种方式育苗,其中,种子育苗最经济高效。针对樟树种苗培育的研究较多,其中,肖祖飞等跟踪了不同贮藏条件樟树种子发芽特性及2个月幼苗的生长指标[7],陈一群等和罗敦等跟踪了不同移植基质对香樟组培苗幼苗生长的影响[8-9],李彦强等跟踪研究了樟树扦插苗的生长规律及生物量[10-12] 。但是与樟树相比,针对阴香苗木培育开展的研究却较少见报,仅周纪刚等概括性介绍了阴香栽培技术[13],以及庄姗等对阴香扦插进行了试验研究[14]。总体而言,基于生产条件对2种树种开展种苗培育研究较少。因此,本研究基于苗圃条件对樟树和阴香开展种苗培育试验,了解种子萌发过程中主要营养物质的动态变化,并对幼苗生长开展一年期的长期跟踪研究,对于探索一套适合生产一线的苗圃育苗方案、促进产业发展具有积极意义。
1 材料与方法
1.1 试验材料
从广西壮族自治区调入成熟樟树种子和阴香种子各1批,调入前经过种子调制和混样,可以直接开展后续试验。种子调回后统一放置于华南农业大学经济林研究中心实验室备用,4 ℃低温密封保存。
1.2 试验方法
1.2.1 苗圃发芽试验
分别将樟树种子和阴香种子放置在常温、50、80 ℃的水中浸泡48 h,保持种子与水的体积比约为1 ∶3,让其自然冷却。浸种结束后将种子分别放入沙和黄心土2种基质中进行自然发芽。所有试验均在同一温室大棚中进行,保持环境条件和水肥条件一致性,所有组合重复3次。
发芽率=(实际发芽数/供试种子总数)×100% 。
1.2.2 种子萌发过程中营养物质变化研究
将萌发的种子根据胚根长划分为5个阶段:阶段A(浸种前未萌发种子);阶段B(胚根长0~<1 cm的种子);阶段C(胚根长1~<2 cm的种子);阶段D(胚根长2~<3 cm的种子);阶段E(胚根长3~4 cm的种子)。分别测定樟树种子和阴香种子不同萌发阶段可溶性糖、可溶性淀粉、蛋白质和脂肪的百分含量。
可溶性糖含量测定采用蒽酮比色法;淀粉含量测定采用蒽酮法;粗脂肪含量测定采用索式抽提法;蛋白质含量通过提取蛋白质上清液后,用Sepectrophotometer ND-1000分光光度计测定。每处理每指标3次重复,取平均值。
1.2.3 基质配备及营养袋育苗
1.2.3.1 栽培基质的配备
栽培基质(泥炭土 ∶椰糠 ∶珍珠岩=20 ∶4 ∶1)与黄心土按1 ∶3~4的比例混合,加入少量石灰、过磷酸钙和有机肥。
1.2.3.2 营养袋育苗
分别于冬季和翌年春季进行种子播种,待种子萌发后即移入7 cm×11 cm的营养袋中进行育苗。
1.2.4 生长量的测定
1.2.4.1 月度生长指标记录
随机挑选100株健壮的樟树幼苗和阴香幼苗并做好标记,于每月26—30日记录各株苗木的地径、苗高和叶片数,为期1年。冬播苗测量时间为3月至翌年2月,春播苗测量时间为5月至翌年4月。
1.2.4.2 年生长量测定
随机挑选一年生樟树幼苗和阴香幼苗,测定其植株总干质量、根干质量、茎干质量、叶干质量和叶面积5个生长指标,每指标5个重复。
质量测定采用烘箱高温烘干法。分别称取全株、根、茎、叶片的鲜质量,然后将其放于烘箱中,调至135 ℃恒温烘干4 h至恒质量,随后快速放入干燥皿中,冷却后称取质量。
叶片面积用Li-3100型台式叶面积仪测定。
1.3 数据分析
采用Microsoft Excel 17.0、SAS 9.0、SPSS 23.0等统计软件进行数据分析。
2 结果与分析
2.1 苗圃发芽试验
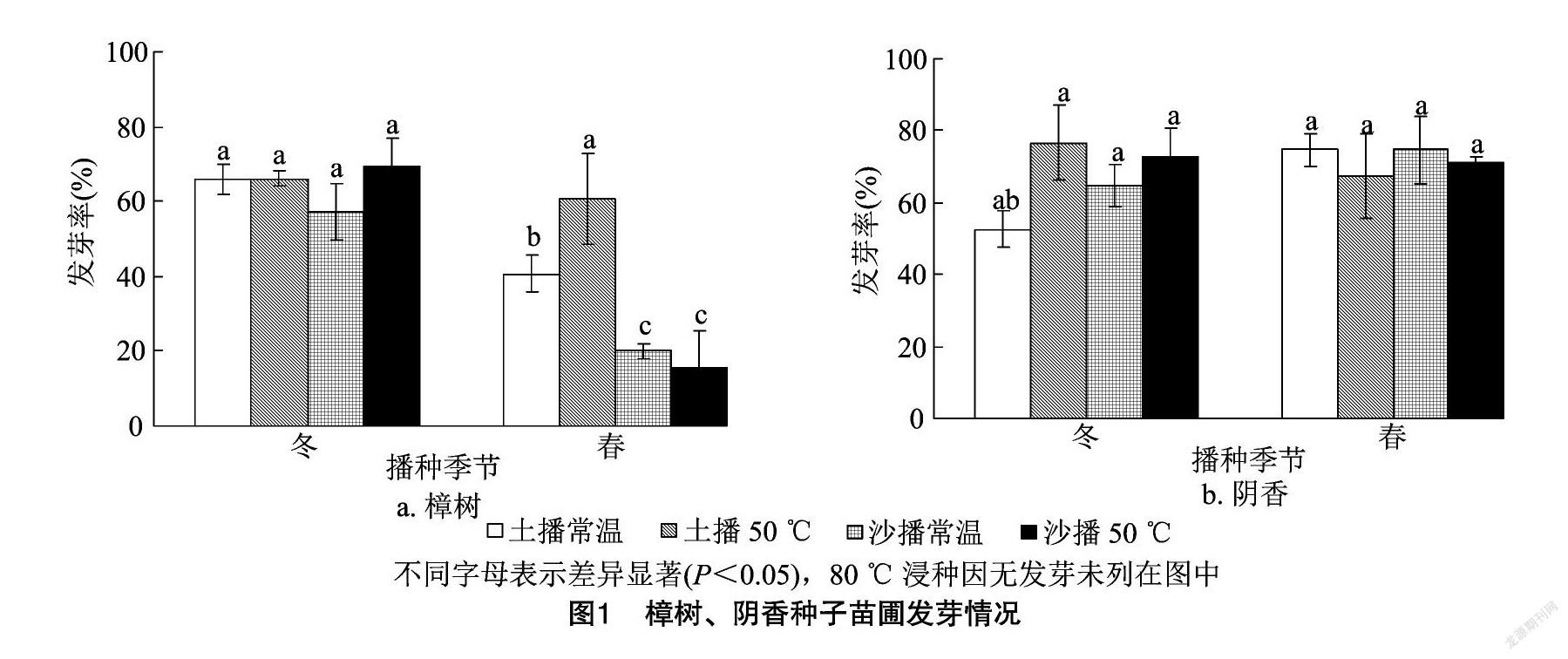
如图1所示,樟树种子沙冬播50 ℃条件发芽率最高,达到69.33%;土冬播50 ℃、土冬播常温和土春播50 ℃这3个处理发芽率较高,达60.00%以上;80 ℃浸种的几个处理均未见发芽。不同处理的樟树种子发芽率由高到低排序依次为沙冬播50 ℃>土冬播常温、土冬播50 ℃>土春播50 ℃>沙冬播常温>土春播常温>沙春播常温>沙春播50 ℃>沙冬播80 ℃、沙春播80 ℃、土冬播80 ℃、土春播80 ℃。
阴香种子土冬播50 ℃处理发芽率最高,达到76.67%;沙冬播50 ℃、沙春播常温、沙春播50 ℃和土春播常温这几个处理发芽率较高,可达70.00%以上;80 ℃浸种的几个处理均未发芽。不同处理的阴香种子发芽率由高到低的排序依次为土冬播 50 ℃>土春播常温、沙春播常温>沙冬播50 ℃>沙春播50 ℃>土春播50 ℃>沙冬播常温>土冬播常温>沙冬播80 ℃、沙春播80 ℃、土冬播80 ℃、土春播80 ℃。
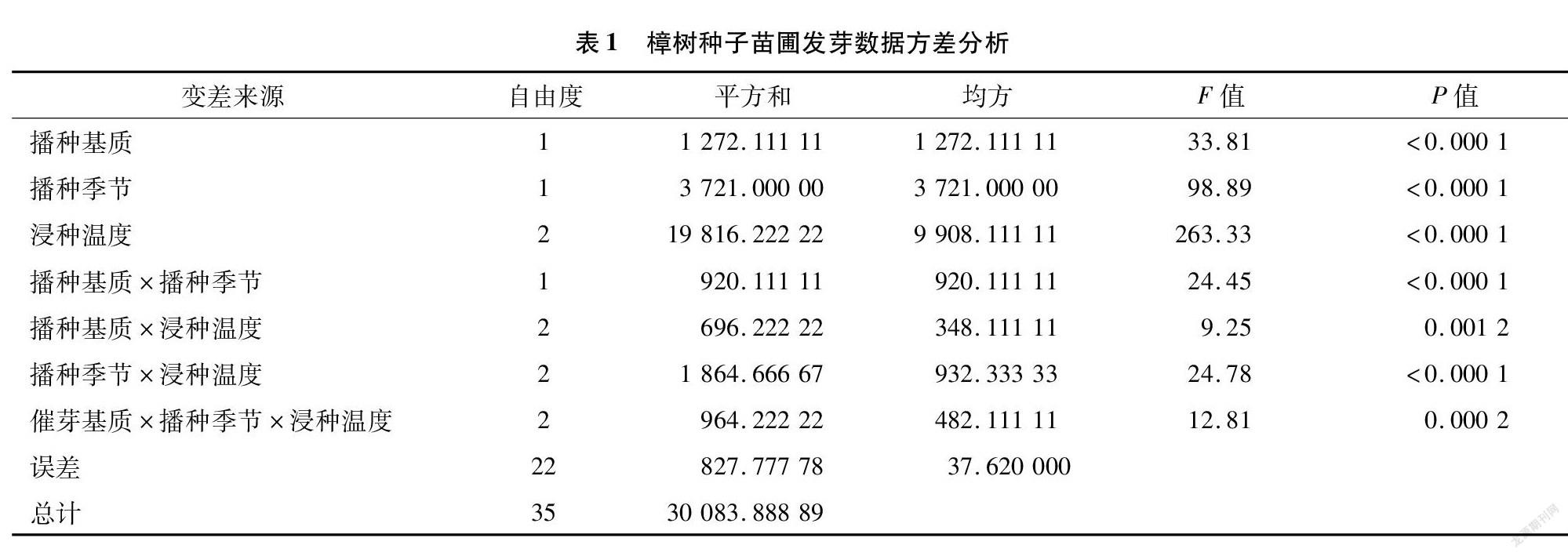
对樟树种子苗圃发芽结果进行方差分析(表1), 其中催芽基质、 播种季节、浸种温度、播种基质与播种季节互作、播种季节与浸种温度互作、催芽基质与播种季节和浸种温度互作影响极显著(P<0.01)。
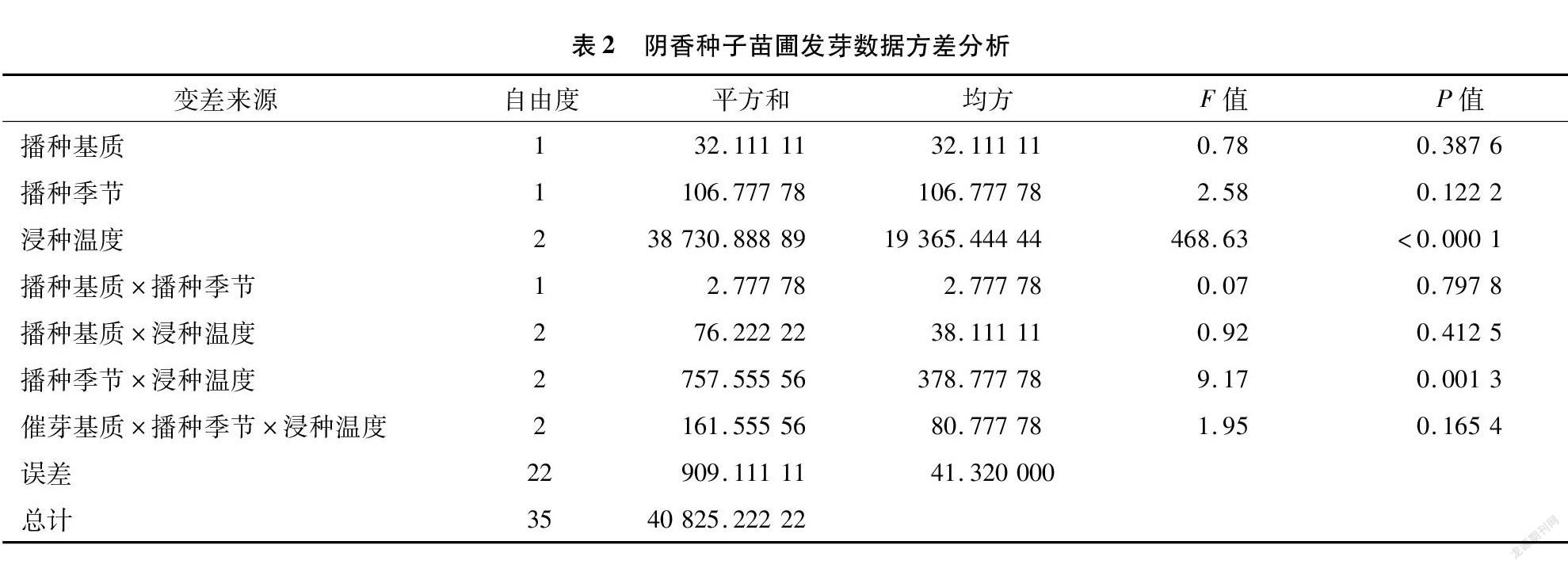
而阴香种子苗圃发芽方差分析结果显示(表2),浸种温度、播种季节与浸种温度互作的影响极显著(P<0.01),其余结果均不显著。
2.2 种子萌发过程中营养物质的动态变化
可溶性糖、蛋白质、脂肪和淀粉是种子重要的营养成分。在萌发过程中,樟树种子和阴香种子可溶性糖、蛋白质和脂肪这3种营养物质含量均整体下降。其中,樟树种子的可溶性糖、蛋白质含量在萌发初期即大幅下降,之后变化幅度小且波动。至阶段E,樟树种子的可溶性糖、蛋白质和脂肪含量分别降低了50.91%、41.14%和22.19%,其中脂肪的降幅较可溶性糖和蛋白质都小。阴香种子蛋白质和脂肪降幅较大,至种子阶段E,2种营养成分分别降低了43.15%和39.58%,而可溶性糖含量相对稳定(图2)。
2种种子淀粉含量与其余3种营养成分变化相反,呈波动升高。其中,樟树种子初始淀粉含量为0.17%,至阶段E增长至0.69%,为初始值的4.06倍;阴香种子初始淀粉含量為0.47%,至种子阶段E含量增长至1.09%,为初始值的2.32倍(图2)。
2.3 苗期生长特性的研究
2.3.1 一年生苗木地径变化规律
如图3所示,冬播樟树苗3月上袋后地径即快速增粗,直至10月后逐步增粗放缓;5月上袋的春播苗与冬播苗上袋时地径值相当,随后几个月增粗较快,到10月后进入增粗缓慢增期,变化趋势与冬播苗一致。综合可见,3月至10月为樟树幼苗增粗生长快速期,11月至翌年2月则为缓慢期。
阴香冬播苗6月至7月和10月至翌年3月地径增粗较慢,其余时间增粗较快;春播苗6月至7月和12月至翌年3月增粗较慢,其余时间段增粗较快。综合可见,6月至7月和11月至翌年3为阴香幼苗增粗生长缓慢期,其余时间段增粗较快。
2.3.2 一年生苗木苗高变化规律
试验结果(图4)显示,冬播樟树幼苗3月上袋后出现了2个苗高增长高峰,第1个是6月至10月,第2个是12月至翌年2月。而春播樟树苗同样出现6月至9月和12月至翌年3月2个生长高峰,与冬播苗基本一致。综合可见,樟树苗9月至12月的苗高增速较慢,其余月份较快。
阴香冬播苗在4月至6月和9月至翌年1月增高缓慢,春播苗在翌年2月至4月增高缓慢,其余时间段增高较快。
2.3.3 一年生苗木叶片数变化规律
从图5可见,冬播樟树苗3月至8月叶片数增长较快,8月至12月停滞,随后又进入新一轮快速增长期。春播樟树苗5月至9月叶片数快速增长,10月至翌年1月基本停止增加,随后又进入新一轮快速增长期。
阴香冬播苗9月至翌年1月,春播苗10月至翌年1月,皆为叶片数增长缓慢期,其余时间叶片数增长较快。冬播苗和春播苗叶片数生长趋势基本一致。
2.3.4 一年生苗木生长量综合比较
为进一步综合评价樟树和阴香冬播苗和春播苗的生长差异,对2种幼苗地径、苗高、叶数、叶面积指数、植株干质量、叶干质量、茎干质量、根干质量共8个生长指标进行比较,以评价不同播种时期的生长差异。
如表3所示,樟树一年生冬播苗各项生长量指标均优于春播苗,其中茎干质量、苗高、地径、叶数这4个指标差异较大。进一步进行方差分析和多重比较,樟树冬播苗和春播苗的茎干质量、苗高差异极显著(P<0.01),地径和叶片数差异显著(P<0.05)。而阴香一年生冬播苗各生长指标与春播苗相当,均差异不显著。其中,地径、苗高、叶面积指数、株总干质量、叶干质量、茎干质量冬播苗稍高于春播苗,而叶片数和根干质量则春播苗稍高于冬播苗。
3 结论与讨论
樟树种子无论采取常温浸种还是50 ℃温水浸种,冬播发芽率较高于春播,表明樟树适宜冬播环境,生产中应优先选择冬播育苗。而在不同基质的春播催芽试验中,樟树土壤基质播种发芽率明显优于沙基质,原因可能是土壤的保水保温性能更好,给种子提供更好的水温环境。如果生产中不可避免需在春季播种育苗,可采用50 ℃土播的催芽方式,以提高樟树种子发芽率。阴香不同处理条件种子苗圃播种发芽率均较高,冬季采取50 ℃温水浸种可以明显提高发芽率。综合分析浸种温度对樟树种子和阴香种子萌发的影响,大部分处理50 ℃温水优于室温,主要原因是温水能促进种子吸水,而种子充分吸水是种子萌发的必要条件,能有效提高种子发芽率[15]。但是,由于80 ℃的浸种温度过高导致种子死亡,严重影响发芽率,因此不能根据樟树硬质种壳而采用高温浸种的催芽方式。结合发芽数据和方差分析,樟树种子苗圃播种最优组合为土壤、冬播、50 ℃浸种处理;阴香种子苗圃播种最优组合为土壤、冬播、50 ℃浸种处理。
对樟树种子和阴香种子主要营养物质进行测定发现,樟树种子和阴香种子内脂肪和蛋白质含量均较高,可溶性糖次之,淀粉含量最低。由此判断樟树、阴香种子属于脂肪含量较高的油料种子,而高脂肪含量对于后续种子萌发和幼苗生长具有积极意义[16]。相关研究显示,脂肪类种子对于贮藏和萌发期环境要求较高[17],必须给予较适宜的贮藏环境和萌发条件。在本试验中,可溶性糖、蛋白质和脂肪含量随种子萌發均降低,表明种子通过消耗这些营养物质为萌发提供必需能量[18]。其中,樟树种子和阴香种子蛋白质和脂肪消耗量较可溶性糖多,表明前两者是种子萌动的主要营养源。蛋白质会逐渐转化成可溶性蛋白,进而水解成游离氨基酸[19],用于合成新蛋白质或被去氨基后为呼吸氧化提供碳架,为种子萌发和新组织、新器官的形成提供氮源,对种子各类生理活动的正常进行和萌发过程中的营养供给有重要作用[20]。而脂肪首先水解为甘油和脂肪酸,然后大部分脂肪酸在乙醛酸体内氧化分解为乙酰CoA,并通过乙醛酸循环转化为糖,进而被萌发时的生长发育等生理活动消耗[21]。樟树、阴香种子贮藏的脂肪就是通过乙醛酸循环迅速消耗,并且伴有可溶性糖和淀粉增长的现象[22]。可溶性糖能为种子萌发提供碳源、能量等,是萌发到光合自养前的呼吸底物[23-24],而2种种子通过消耗自身的可溶性糖,以及通过脂肪等分解后形成的可溶性糖,均能支持种子的正常萌发。相关研究表明,可溶性糖和淀粉在种子中作为贮藏的碳水化合物,两者的合成方向和合成量处于动态平衡[25-26],这2个因素可能是导致樟树种子和阴香种子在萌发过程中淀粉含量不断升高的主要原因。
对一年生幼苗生长指标的跟踪发现,樟树幼苗3月至10月为增粗快速期,6月至9月和12月至翌年3月是增高快速期,4月至9月为叶片数增长快速期;阴香幼苗4月至11月为增粗快速期,6月至10月是增高快速期,1月至10月为叶片数增长快速期。因此,针对性地在茎增粗生长期多施用磷肥和钾肥,在长叶和拔高期多施用氮肥和钾肥具有重要的指导意义[27-29]。樟树一年生冬播苗的各项生长指标均比一年生春播苗高,尤其茎干质量、苗高、叶数和地径4个指标差异明显。因此,樟树最佳播种育苗期在冬季,此时可在同等条件下培育出更健壮的苗木。阴香除根干质量、叶片数这2个指标外,一年生冬播苗的各项生长指标均比一年生春播苗高,但是差异不显著,因此阴香在冬季或者次年春季进行播种育苗皆宜。
参考文献:
[1]燕一波. 4种樟科幼树叶片对汽车尾气胁迫的生理响应[J]. 四川农业大学学报,2017,35(2):234-240.
[2]李小婷,陈 斌,王海军,等. 城市夜间灯光对香樟生长的影响[J]. 应用生态学报,2019,30(7):2284-2290.
[3]刘新亮,章 挺,邱凤英,等. 造林密度对材用樟树幼林生长和蓄积量的影响[J]. 中南林业科技大学学报,2019,39(3):23-27,60.
[4]陈柏旭,黄腾华,韦鹏练,等. 阴香木材物理力学性质研究[J]. 西北林学院学报,2016,31(5):271-275.
[5]Yang Z R,An W L,Liu S S,et al. Mining of candidate genes involved in the biosynthesis of dextrorotatory borneol in Cinnamomum burmannii by transcriptomic analysis on three chemotypes[J]. Peer J,2020,8:e9311.
[6]Fu J,Zeng C,Zeng Z L,et al. Cinnamomum camphora seed kernel oil ameliorates oxidative stress and inflammation in diet-induced obese rats[J]. Journal of Food Science,2016,81(5):H1295-H1300.
[7]肖祖飞,张北红,李凤,等. 樟树种子发芽及幼苗生长研究[J]. 安徽农学通报,2020,26(7):40-45,63.
[8]陈一群,丘佐旺,汪迎利,等. 樟树组培苗轻基质育苗技术研究[J]. 亚热带植物科学,2015,44(2):140-145.
[9]罗 敦,黄汉林,吴坤林,等. 移栽基质对香樟组培苗生长的影响[J]. 安徽农业科学,2020,48(2):130-132.
[10]李彦强,孙小艳,钟永达,等. 几种常见绿化树种扦插苗生长及固碳能力[J]. 江苏农业科学,2016,44(7):256-258.
[11]李彦强,胡晓健,高 柱,等. 大棚内不同基质扦插对香樟苗生长的影响[J]. 北方园艺,2015(22):58-60.
[12]叶润燕,童再康,张俊红,等. 樟树茎段组培快繁[J]. 浙江农林大学学报,2016,33(1):177-182.
[13]周纪刚,徐 平,舒夏竺,等. 阴香高效栽培技术[J]. 林业实用技术,2014(4):58-59.
[14]庄 姗,胡松竹,王光云,等. 阴香扦插繁殖试验[J]. 安徽农业科学,2009,37(18):8639-8640.
[15]程 瑶,方向文,蒋志荣,等. 温水浸种对蒙古黄芪种子萌发特性的影响[J]. 植物科学学报,2017,35(3):413-420.
[16]Martínez-Arias R,Ronquillo-López M,Schechert A.Quantification of oil content in intact sugar beet seed by near-infrared spectroscopy[J]. Agronomy,2018,8(11):254.
[17]朱 鹤,李憬霖,徐敏,等. 作物种子的营养成分与其萌发特性关系的探讨[J]. 种子,2019,38(8):47-52.
[18]Fatihi A,Boulard C,Bouyer D,et al. Deciphering and modifying LAFL transcriptional regulatory network in seed for improving yield and quality of storage compounds[J]. Plant Science,2016,250:198-204.
[19]Liu R R,Wang L,Tanveer M,et al. Seed heteromorphism:an important adaptation of halophytes for habitat heterogeneity[J]. Frontiers in Plant Science,2018,9:1515.
[20]Müntz K,Belozersky M A,Dunaevsky Y E,et al. Stored proteinases
and the initiation of storage protein mobilization in seeds during germination and seedling growth[J]. Journal of Experimental Botany,2001,52(362):1741-1752.
[21]Wei S,Liu Y N,Wu M L,et al. Disruption of the transcription factors Thi2p and Nrm1p alleviates the post-glucose effect on xylose utilization in Saccharomyces cerevisiae[J]. Biotechnology for Biofuels,2018,11:112.
[22]孙 建,周红英,乐美旺,等. 芝麻种子萌发动态及其代谢生理变化研究[J]. 中国农业科技导报,2020,22(8):41-48.
[23]马 琳,王志清,张秀莲,等. 北细辛种子储藏过程中储藏物质变化的研究[J]. 种子,2015,34(12):1-3,8.
[24]王海杰,杨 宇,黄家权. 瓜儿豆种子萌发糖代谢动态分析[J]. 山东农业大学学报(自然科学版),2019,50(1):99-102.
[25]Pairochteerakul P,Jothityangkoon D,Ketthaisong D,et al. Seed germination in relation to total sugar and starch in endosperm mutant of sweet corn genotypes[J]. Agronomy,2018,8(12):299.
[26]Tan-Wilson A L,Wilson K A.Mobilization of seed protein reserves[J]. Physiologia Plantarum,2012,145(1):140-153.
[27]高小麗. 施磷对豌豆产量及其构成因素的影响[J]. 作物杂志,2016(1):125-128.
[28]李 琦,裴怀弟,马忠明,等. 钾肥与有机肥配施对食用百合根际土壤酶活性、养分含量及鳞茎产量的影响[J]. 中国土壤与肥料,2020(1):91-99.
[29]权宝全,吕瑞洲,王贵江. 不同施氮量对甘薯生长发育及产量的影响[J]. 东北农业科学,2019,44(6):14-17.
