木芙蓉对淹水胁迫的响应及其耐涝性综合评价
2022-04-14徐倩邬梦晞曾心美姜贝贝贾茵李梦霞张曼瑜张璐蔡露黄文沛潘远智
徐倩, 邬梦晞, 曾心美, 姜贝贝, 贾茵, 李梦霞, 张曼瑜, 张璐,蔡露, 黄文沛, 潘远智*
木芙蓉对淹水胁迫的响应及其耐涝性综合评价
徐倩1, 邬梦晞1, 曾心美2, 姜贝贝1, 贾茵1, 李梦霞2, 张曼瑜1, 张璐1,蔡露2, 黄文沛1, 潘远智1*
(1. 四川农业大学风景园林学院,成都 611130;2. 成都市植物园,成都 611083)
为挖掘木芙蓉()耐涝种质资源,对13个木芙蓉品种水淹15 d后的形态和生理生化指标进行研究,并构建耐涝评价体系。结果表明,水淹胁迫后,木芙蓉品种各性状的变化趋势和幅度均不同,部分性状间的相关性显著(<0.05)。14个指标按贡献率大小归因为形态生长指标、不定根形态指标、膜脂过氧化指标、根系生物量指标、抗氧化指标及渗透调节指标。根据综合耐涝评价值()进行聚类分析,表明13个木芙蓉品种可分为强耐涝(3个品种)、较强耐涝(5个品种)、中等耐涝(3个品种)和不耐涝(2个品种)等4个类型。因此,株高、不定根数、不定根长可作为木芙蓉苗期耐涝性快速、简便的鉴定指标。
木芙蓉;水淹;耐涝性;隶属函数法;综合评价
随着全球气候变暖和城市化进程加快,干旱、暴雨等极端天气频繁出现,城市生态问题也愈发严重,大范围的地表径流是城市洪涝灾害的主要诱因之一[1]。此外,在园林应用中,管理措施不当造成的土壤板结、过量灌溉等,也会导致涝害的发生,轻者植物生长受抑制,重者引起植株死亡[2]。水淹胁迫是由于水分过多给植物带来的一系列次生胁迫,其实质是氧气的缺乏。缺氧与线粒体呼吸抑制引起的能量供应紊乱、氧化还原状态变化及活性氧(ROS)的积累有关[3]。植物也进化出了一系列的形态适应机制应对水淹胁迫,如通气组织[4]、不定根[5]和根系泌氧结构[6]等。
芙蓉花(),又名木芙蓉,为锦葵科(Malvaceae)木槿属落叶小乔木。木芙蓉在我国栽培历史悠久,目前已培育出100多个品种,但大规模种植和应用的品种不多,因为大部分木芙蓉品种在栽培中对水分的需求差异较大,导致木芙蓉在实际应用中往往出现生长不良等问题,严重影响其观赏性和应用效果。近年来,随着自主选育力度的加大,其品种数量也不断增加。因此,建立木芙蓉耐涝性的综合评价体系,客观全面的评价各品种的耐涝性,为其大规模推广应用提供理论依据显得尤为迫切。
不同植物对水淹胁迫的响应不同,同种植物不同品种间对水淹胁迫的响应也存在差异。大量研究建立了菊花()、牡丹()和杨树(sp.)[7–11]等观赏植物的耐涝性评价体系。此外,有研究表明,木芙蓉相对其他园林植物具有较强的耐淹性[12]。目前,水淹胁迫对木芙蓉的影响研究主要集中在光合生理和抗氧化等方面[13–14],而关于建立木芙蓉耐涝评价体系的研究鲜见报道。本研究采用多元统计分析方法,研究水淹胁迫对13个木芙蓉品种14个苗期性状的影响,建立系统科学的综合评价体系,以期为木芙蓉进一步推广应用和加速园林植物耐水湿性育种进程提供理论依据。
1 材料和方法
1.1 材料
供试木芙蓉()均取自成都市植物园。2020年8—10月,选取生长一致的8叶苗龄植株为当年生扦插苗,共计13个品种: ‘百日华彩’(‘Bairihuacai’)、‘醉芙蓉’(‘Zuifurong’)、‘锦蕊’ (‘Jinrui’)、‘醉红’(‘Zuihong’)、‘重瓣白’ (‘Chongban- bai’)、‘牡丹红’(‘Mudanhong’)、‘单瓣红’(‘Danban- hong’)、‘牡丹粉’(‘Mudanfen’)、‘金秋颂’ (‘Jinqiu- song’)、‘彩霞’(‘Caixia’)、‘醉云’(‘Zuiyun’)、‘锦碧玉’ (‘Jinbiyu’)和‘玉蕊’(‘Yurui’),其中‘醉芙蓉’、‘重瓣白’和‘单瓣红’是传统品种,‘醉云’为辐射突变选育,其余均为人工杂交品种。试验期间施用Hoagland营养液,栽植土壤为黄壤土。
1.2 方法
选择生长一致的无病虫害扦插苗,定植于装有等量基质(黄壤土)的塑料花盆(15 cm×15 cm)中。2020年8月,采用盆栽半水淹法,将盆栽苗置于矩形塑料容器(38 cm×26 cm×21 cm)中进行淹水处理。试验采用双因素完全随机设计,因素1为品种(13个),因素2为水分处理(2个): 对照(CK)和淹水(W),对照根据土壤干湿状况,每天补水1次, 维持含水量为田间持水量的60%~70%;淹水则保持水位高于盆栽土表面2~3 cm,每天上午9:00补水以保持水位,有的品种15 d左右叶片严重萎焉,因而确定15 d为采样时间。试验共26个处理,设4次重复,每个处理16盆。
形态指标和生物量测定 用游标卡尺测定植株基部(根茎交界以上5 mm处)的茎粗;观察并统计不定根数量,用游标卡尺测定不定根长和主根长;株高为植株基部到植株顶端的高度;将植株的地上部和地下部分别装进信封放入烘箱中,105 ℃下烘干,根冠比=植株地下部生物量/植株地上部生物量。
生理指标测定 剪取植株自顶部往下第3片完全展开叶测定相对电导率。相对电导率=初始电导率/煮沸电导率×100%。可溶性蛋白含量测定采用考马斯亮蓝G-250法;超氧化物歧化酶(superoxide dis- mutase, SOD)活性测定采用氮蓝四唑(nitro-bluetetra- zolium, NBT)法;过氧化物酶(peroxidase, POD)活性测定采用愈创木酚法;MDA含量测定采用硫代巴比妥酸法;过氧化氢酶(catalase, CAT)采用紫外吸收法[15][15]。
1.3 数据统计和分析
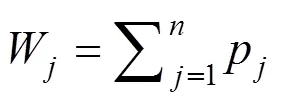
采用SPSS 19.0软件对木芙蓉耐涝指标数据进行统计分析,主要包括主成分分析、相关性分析和聚类分析等多元分析,方差分析采用双因素方差分析和Ducan’s法,显著性水平设定为<0.05,并对13个木芙蓉品种的耐涝性进行排序。
2 结果和分析
2.1 对形态和生理生化指标的影响
由表1~4可见,水淹胁迫后,木芙蓉各品种均产生了不定根,‘牡丹粉’的不定根数量最多,‘百日华彩’最少;‘牡丹红’的不定根最长,‘玉蕊’的最短,说明木芙蓉对水淹胁迫均产生了一定的形态适应机制,且品种间具有一定差异。与对照相比,地和‘锦蕊’; 大部分品种的株高和地径呈下降趋势,‘百日华彩’、‘锦蕊’和‘玉蕊’的株高显著低于其余品种(<0.05),‘百日华彩’、‘醉云’、‘锦碧玉’和‘玉蕊’的地径显著低于其余品种,但这些品种间无显著差异;MDA含量和相对电导率均呈上升趋势,分别是升了6.09%~244.18%和1.67%~62.96%,增幅最大的均是‘玉蕊’;可溶性蛋白含量有的品种上升, 有的品种下降,变化不一; 大部分品种的SOD、POD和CAT呈上升趋势,变化幅度分别为–31.46%~180.11%、–28.26%~91.79%和2.54%~177.81%,增幅最大的品种分别为‘锦碧玉’、‘玉蕊’和‘锦蕊’。这与外部形态的观察结果基本一致(图1)。

表1 水淹胁迫对木芙蓉形态生长指标的影响
ADW: 地上部分生物量; H: 株高; R/C: 根冠比; D: 地径; RL: 主根长; UDW: 地下部分生物量。CK: 对照; W: 水淹; 同列数据后不同字母表示差异显著(<0.05)。
ADW: Above-ground dry weight; H: Height; R/C: UDW/ADW; D: Diameter; RL: Main root length; UDW: Under-ground dry weight; CK: Control; W: Waterlogging. Data followed different letters indicate significant differences at 0.05 level.
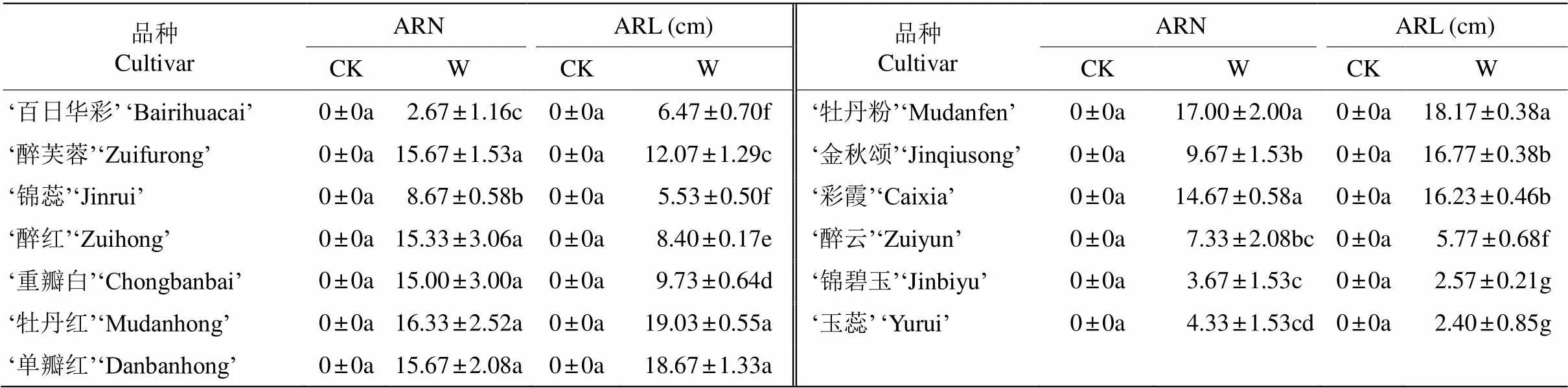
表2 水淹胁迫对木芙蓉不定根形态指标的影响
ARN: 不定根数; ARL: 不定根长; CK: 对照; W: 水淹。同列数据后不同字母表示差异显著(<0.05)。
ARN: Number of adventitious root; ARL: Length of adventitious root; CK: Control; W: Waterlogging. Data followed different letters indicate significant differences at 0.05 level.
SP: 可溶性蛋白含量; RC: 相对电导率; CK: 对照; W: 水淹。同列数据后不同字母表示差异显著(<0.05)。
SP: Soluble protein content; RC: Relative conductivity; CK: Control; W: Waterlogging. Data followed different letters indicate significant differences at 0.05 level.
2.2 指标的耐涝系数及其相关性
品种指标处理与对照的比值更能准确反映耐涝能力的大小。由表5可见,不同品种木芙蓉不同指标的耐涝系数具有差异性,MDA含量、SOD活性和相对电导率的耐涝系数>1,地上和地下生物量的耐涝系数<1,其余性状,如株高、根冠比等的变化不一,导致筛选结果缺乏一致性。
另一方面,由表6可见,各指标间存在不同程度的相关性,部分性状间的相关性达到显著水平(<0.05),如不定根数与不定根长、地下生物量、根冠比、MDA、CAT、POD、SOD和相对电导率间呈显著相关;不定根长与地下干重、根冠比、主根长、MDA、CAT、POD和SOD间呈显著相关, 因而提供的信息发生重叠。此外,各指标在不同品种木芙蓉耐涝能力中发挥的作用也不同(表6),表明木芙蓉的耐涝性是一个复杂的综合性状,利用单项指标直接评价不能客观反映其真实情况。因此,将单一性状的耐涝系数对木芙蓉进行耐涝性评价存在一定片面性。

表4 水淹胁迫对木芙蓉抗氧化酶活性的影响
CK: 对照; W: 水淹; 同列数据后不同字母表示差异显著(<0.05)。
CK: Control; W: Waterlogging. Data followed different letters indicate significant differences at 0.05 level.
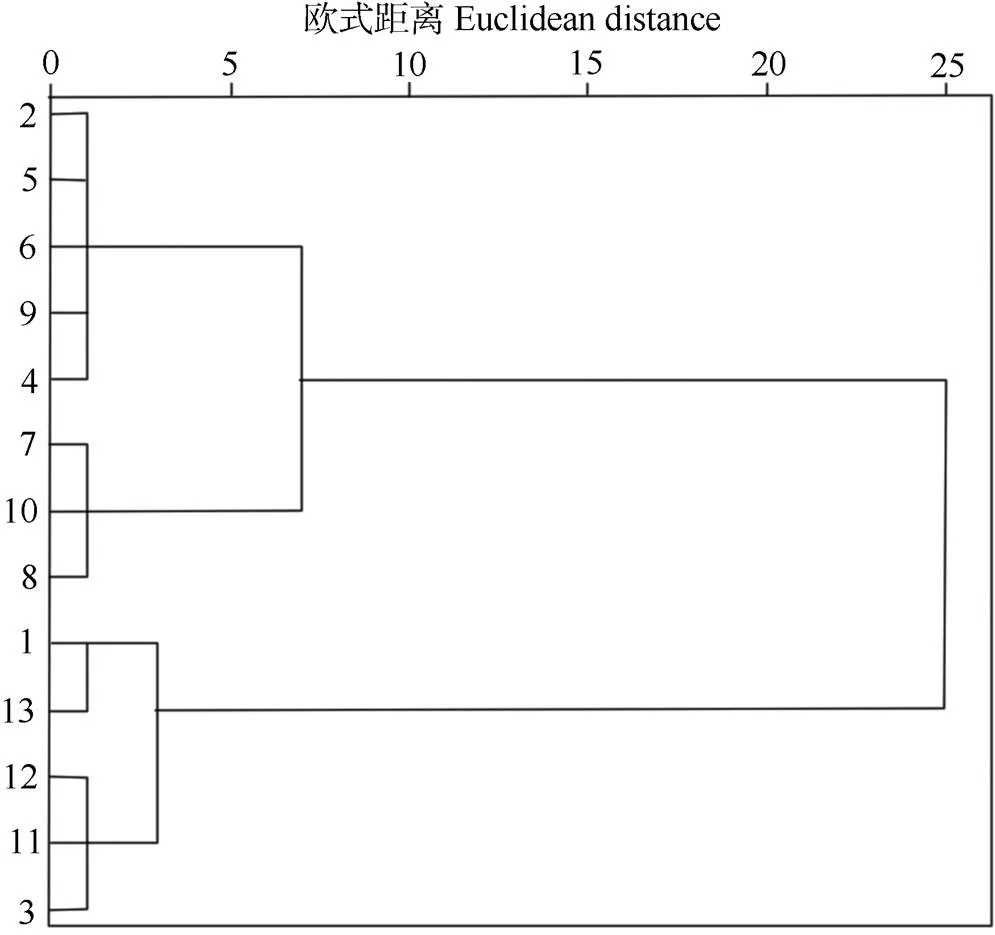
图1 木芙蓉品种的聚类图。1: ‘百日华彩’; 2: ‘醉芙蓉’; 3: ‘锦蕊’; 4: ‘醉红’; 5: ‘重瓣白’; 6: ‘牡丹红’; 7: ‘单瓣红’; 8: ‘牡丹粉’; 9: ‘金秋颂’; 10: ‘彩霞’; 11: ‘醉云’; 12: ‘锦碧玉’ ; 13: ‘玉蕊’。
2.3 主成分分析
本试验运用主成分分析来评价木芙蓉的耐涝性。对14个指标的耐涝系数进行主成分分析,前6个主成分(CI1~CI6)的贡献率分别为24.935%、17.453%、14.840%、10.414%、9.728%和8.357%, 累计贡献率达85.728%,其余可忽略不计(表7)。分别用这6个新的综合指标进行概括性分析,对应的特征向量表达式分别为:1=–0.0431–0.0082+ 0.4983+0.3074+0.4185-0.3596+0.4237+ 0.33578–0.3419–0.03710+0.12411–0.18212–0.06313–0.34014;2=0.5571+0.5432+0.1043– 0.2524+0.1245–0.2966+0.1287+0.2028+0.2829+0.39410+0.22111+0.41012+0.07713+ 0.199814;3=–0.0841–0.0572–0.0533+ 0.1264–0.2005+0.0406+0.1097+0.2848– 0.04419+0.36710–0.51811+0.15212+0.36713– 0.114414;4=–0.1521–0.11562+0.0803- 0.3094+0.1295–0.2096+0.1857–0.0108+ 0.1469-0.25210–0.00111–0.15512+0.56113– 0.31814;5=0.2911+0.3352–0.0793+0.0594– 0.2435+0.2126+0.1997–0.1718–0.3269– 0.10410+0.10311–0.27812+0.29613–0.09214;6=–0.0421–0.0532–0.0343+0.4784+0.0675+ 0.3546+0.0637+0.2148+0.1739–0.12210+0.31811+0.23412+0.19613+0.19514,1~14分别代表不定根数、不定根长、地上干质量、地下干质量、株高、根冠比、地径、主根长、MDA、CAT、POD、SOD、可溶性蛋白、相对电导率等性状。
第1主成分主要包括地上生物量、株高、根冠比、地径和主根长,相关系数分别为0.498、0.418、–0.359、0.423和0.357,表明第1主成分主要反映了生物量等生长形态指标,定义为形态生长指标;第2主成分主要包括不定根数和不定根长,相关系数分别为0.557和0.543,定义为不定根形态指标;第3主成分中,MDA、SOD和相对电导率的相关系数分别为-0.041、0.152和-0.144,定义为膜脂过氧化指标,第4主成分主要包括地下生物量,可反映全部数据的10.414%信息,定义为根系生物量指标;第5主成分主要包括POD和CAT,相关系数分别为0.103和–0.104,定义为抗氧化指标;第6主成分中,可溶性蛋白的相关系数为0.196,定义为渗透调节指标。因此,本研究利用主成分分析把多个木芙蓉苗耐涝指标归为了具有代表性的因子, 进一步量化变量数目,获取了木芙蓉苗期耐涝性主要影响指标。

表5 木芙蓉苗期指标的耐涝系数
ARN: 不定根数; ARL: 不定根长; ADW: 地上部分生物量; UDW: 地下部分生物量; H: 株高; R/C: 根冠比; D: 地径; RL: 主根长; SP: 可溶性蛋白含量; RC: 相对电导率。
ARN: Number of adventitious root; ARL: Length of adventitious root; ADW: Above-ground dry weight; UDW: Under-ground dry weight; H: Height; R/C: UDW/ADW; D: Diameter; RL: Main root length; SP: Soluble protein content; RC: Relative conductivity.

表6 木芙蓉苗期指标间的相关性
*:<0.05.ARN: 不定根数; ARL: 不定根长; ADW: 地上部分生物量; UDW: 地下部分生物量; H: 株高; R/C: 根冠比; D: 地径; RL: 主根长; SP: 可溶性蛋白含量; RC: 相对电导率。
*:<0.05. ARN: Number of adventitious root; ARL: Length of adventitious root; ADW: Above-ground dry weight; UDW: Under-ground dry weight; H: Height; R/C: UDW/ADW; D: Diameter; RL: Main root length; SP: Soluble protein content; RC: Relative conductivity.

表7 综合指标的相关系数及贡献率
2.4 耐涝性综合评价
由表8可见,对于相同综合指标, 如CI1,水淹胁迫下‘单瓣红’的隶属函数(1)最大,为1.000,表明其在CI1表现为耐涝性最强,而‘玉蕊’的(1)最小,为0.000,说明其在CI1表现为耐涝性最弱。
根据各综合指标贡献率大小,计算获得6个综合指标的权重分别为0.291、0.204、0.173、0.122、0.113和0.097。
根据综合耐涝评价值()对各品种的耐涝能力进行排序。由表8可见,‘牡丹粉’的值最大(0.807),表明其耐涝性最强;‘玉蕊’的值最小(0.240),说明其耐涝性最弱。13个木芙蓉品种耐涝性从强到弱分别为:‘牡丹粉’、‘单瓣红’、‘彩霞’、‘金秋颂’、‘醉红’、‘醉芙蓉’、‘重瓣白’、‘牡丹红’、‘锦蕊’、‘锦碧玉’、‘醉云’、‘百日华彩’、‘玉蕊’。采用最远距离法对值进行聚类分析(图1),可以将13个品种分为4类:‘单瓣红’和‘牡丹粉’为第1类,属于强耐涝型;‘醉芙蓉’、‘重瓣白’、‘牡丹红’、‘金秋颂’和‘醉红’为第2类,属于较强耐涝型;‘锦碧玉’、‘醉云’和‘锦蕊’为第3类,属于中等耐涝型;‘玉蕊’和‘百日华彩’为第4类,属于不耐涝型。评价结果与外部形态观察结果基本一致(图2)。

表8 木芙蓉的综合指标值、权重、u(Xj)、D值和耐涝性
3 结论和讨论
水分条件是植物生长的重要环境因子之一,影响着形态、生理生化代谢和地理分布范围。西南地区位于东北-西南走向气候生态过渡带的南端,对气候变化尤为敏感,是我国降水局部差异最大、变化最复杂的地区之一[18],因此西南地区园林植物通常受到水淹胁迫的影响。植物的耐涝程度直接影响其在水淹条件下的生长发育[19–20]。目前,菊花、牡丹、杨树等观赏植物耐涝性已有研究报道,利用单项指标或多项指标提出了多种鉴定耐涝性的方法,如根据外形指标,制定等级评分标准和评价方案,建立评价体系[8],该方法简单易行,但是没有考虑到每个性状对耐涝性起的作用不同。翟丽丽等[7]提出利用层次分析法建立综合评价体系,量化了不同因子对综合性状的影响,增加了评价体系的可操作性,但是该方法的评价因子需要主观赋权,因此具有一定局限性。朱向涛等[10]提出以各指标的耐涝系数为衡量耐涝性的指标,利用主成分和聚类分析法对牡丹耐涝性进行综合评价,弥补了前一方法的不足。但是目前关于木芙蓉耐涝评价体系的研究还鲜见报道,不同植物耐涝性具有差异性,亟需构建木芙蓉耐涝体系的评价方法。

图2 水淹胁迫对木芙蓉形态的影响。A: 对照; B: 水淹; C: 水淹后的根部; 1: ‘百日华彩’; 2: ‘醉芙蓉’; 3: ‘锦蕊’; 4: ‘醉红’; 5: ‘重瓣白’; 6: ‘牡丹红’; 7: ‘单瓣红’; 8: ‘牡丹粉’; 9: ‘金秋颂’; 10: ‘彩霞’; 11: ‘醉云’; 12: ‘锦碧玉’ ; 13: ‘玉蕊’。标尺=10 cm
不定根的形成是植物能否适应水淹胁迫的关键[21]。本研究中,水淹胁迫后,木芙蓉不同品种产生不定根的数量具有差异性,耐涝性强的品种不定根数量更多(如‘单瓣红’、‘牡丹粉’),地上部和地下部生物量减小,根冠比、地径、株高和主根长的变化不一,且耐涝性强的品种耐涝系数更大,这与刘聪聪等[22]对樱桃番茄()的研究结果一致,原因在于叶片黄化脱落、新叶发育受阻导致地上部生物量减小。水淹胁迫的实质是氧气的缺乏(氧气在空气中的传播速度是水里的104倍)[2],线粒体呼吸抑制引起的能量供应紊乱,初生根系大量死亡,主根长及地下部分生物量减小[23–24]。植物为了获取足够的氧气,基径变粗,皮孔增生, 原有初生根系大量死亡,不定根大量形成[25–27]。本研究中,采用主成分分析对14个耐涝指标进行评价分析,贡献率最高的两个因子分别是形态生长指标和不定根形态指标,表明地上生物量、株高、不定根数、不定根长是评价木芙蓉耐涝性的关键指标。
在水淹胁迫条件下,植物会通过渗透调节物质的积累来降低渗透势而维持细胞膨压,减轻受害程度,研究表明渗透调节能力的强弱与植物的抗涝性密切相关[28]。本研究中,水淹胁迫后可溶性蛋白含量有的升高有的降低,这与吴江等[29]、朱向涛等[10][10]的研究结果一致。水淹低氧胁迫下,ROS增加并大量积累,破坏活性氧产生和清除系统的平衡,本研究中,水淹胁迫后,MDA含量和相对电导率升高,说明膜脂发生过氧化和脱脂化,膜的流动性、膜选择性吸收等功能的降低或丧失,且耐涝性弱的品种上升幅度更大(如‘玉蕊’、‘醉云’等)。植物为了清除过多的ROS,会激发体内的活性氧清除系统,本研究中,淹水胁迫后,SOD活性均上升,POD和CAT活性有的上升,有的下降。耐涝性强的品种变化幅度更小,这与Arbona等[30]的研究结果一致,原因在于耐涝性强的品种具有更强的延迟活性氧受害的能力,抗氧化酶活性变化不大,甚至低于对照处理,而耐涝性弱的品种抗氧化酶活性大幅上升,随着胁迫时间的延长,当ROS含量超过自身能承受的范围时,抗氧化酶活性降低[31]。
植物的耐涝性是由多种因素相互作用而构成的一个较为复杂的综合性状,用单一指标进行评价具有一定局限性,必须将多个指标运用多种方法进行综合分析才能真实反映植物的耐涝性[32]。本研究中,13个木芙蓉品种苗期的14个耐涝指标按贡献率可归因为形态生长指标、不定根形态指标、膜脂过氧化指标、根系生物量指标、抗氧化指标和渗透调节指标等6类。各因子得分不同,其中地上生物量、株高、不定根数、不定根长的得分较高,说明在木芙蓉耐涝性鉴定中有较大贡献。但是考虑到操作的便捷性,本研究认为株高、不定根数、不定根长可作为木芙蓉苗期耐涝性鉴定的最优指标,筛选出3个强耐涝品种、5个较强耐涝品种、3个中等耐涝品种和2个不耐涝品种。本研究筛选出的耐涝种质,可作为木芙蓉耐涝育种和机理研究材料,为不同立地条件(水分)的绿地配置适宜的木芙蓉品种提供了实践依据,同时为加速园林植物耐水湿性育种进程、助力海绵城市建设提供了理论依据。这可为今后木芙蓉耐涝性的鉴定提供理论支持。
[1] BAILEY-SERRES J, VOESENEK L A C J. Flooding stress: Accli- mations and genetic diversity [J]. Annu Rev Plant Biol, 2008, 59: 313– 339. doi: 10.1146/annurev.arplant.59.032607.092752.
[2] LORETI E, VAN VEEN H, PERATA P. Plant responses to flooding stress [J]. Curr Opin Plant Biol, 2016, 33: 64–71. doi: 10.1016/j.pbi. 2016.06.005.
[3] ZHOU W G, CHEN F, MENG Y J, et al. Plant waterlogging/flooding stress responses: From seed germination to maturation [J]. Plant Physiol Biochem, 2020, 148: 228–236. doi: 10.1016/j.plaphy.2020.01.020.
[4] YAMAUCHI T, WATANABE K, FUKAZAWA A, et al. Ethylene and reactive oxygen species are involved in root aerenchyma formation and adaptation of wheat seedlings to oxygen-deficient conditions [J]. J Exp Bot, 2014, 65(1): 261–273. doi: 10.1093/jxb/ert371.
[5] CHEN L, OGOREK L L P, PEDERSEN O, et al. Oxygen in the air and oxygen dissolved in the floodwater both sustain growth of aquatic adventitious roots in rice [J]. J Exp Bot, 2020, 72(5): 1879–1890. doi: 10.1093/jxb/eraa542.
[6] YAMAUCHI T, COLMER T D, PEDERSEN O, et al. Regulation of root traits for internal aeration and tolerance to soil waterlogging- flooding stress [J]. Plant Physiol, 2018, 176(2): 1118–1130. doi: 10. 1104/pp.17.01157.
[7] QU L L, FANG W M, CHEN F D, et al. Comprehensive appraisal of the ornamental value and drought and flooding resistance of Guoqingwith small inflorescences [J]. Sci Agric Sin, 2012, 45 (4): 734–742. doi: 10.3864/j.issn.0578-1752.2012.04.014.
翟丽丽, 房伟民, 陈发棣, 等. 国庆小菊观赏性和耐旱、涝性的综合评价[J]. 中国农业科学, 2012, 45(4): 734–742. doi: 10.3864/j.issn. 0578-1752.2012.04.014.
[8] YIN D M, GUAN Z Y, CHEN S M, et al. Establishment of evaluation system for waterlogging tolerance and identification of waterlogging tolerance in chrysanthemum morifolium and its related genera plants [J]. J Plant Genet Resour, 2009, 10(3): 399–404. doi: 10.13430/j.cnki. jpgr.2009.03.021.
尹冬梅, 管志勇, 陈素梅, 等. 菊花及其近缘种属植物耐涝评价体系建立及耐涝性鉴定[J]. 植物遗传资源学报, 2009, 10(3): 399–404. doi: 10.13430/j.cnki.jpgr.2009.03.021.
[9] SHI H J, XI J W, ZHU X T. Screening of waterlogging tolerant peony cultivars by phenotypic characteristics [J]. J Jinling Inst Technol, 2018, 34(1): 88–92. doi: 10.16515/j.cnki.32-1722/n.2018.01.0021.
时浩杰, 奚建伟, 朱向涛. 应用表型特征筛选耐涝牡丹品种研究[J]. 金陵科技学院学报, 2018, 34(1): 88–92. doi: 10.16515/j.cnki.32- 1722/n.2018.01.0021.
[10] ZHU X T, JIN S H, AI J G, et al. Evaluation of waterlogging tolerance of peony variety [J]. J Nucl Agric Sci, 2017, 31(3): 607–613. doi: 10. 11869/j.issn.100-8551.2017.03.0607.
朱向涛, 金松恒, 哀建国, 等. 牡丹不同品种耐涝性综合评价 [J]. 核农学报, 2017, 31(3): 607–613. doi: 10.11869/j.issn.100-8551.2017. 03.0607.
[11] GUO X Y, HUANG Z Y, XU A C, et al. A comparison of physiological, morphological and growth responses of 13 hybrid poplar clones to flooding [J]. Forestry, 2011, 84(1): 1–12. doi: 10.1093/forestry/cpq037.
[12] WANG Z Y. Research on physiological characters of nine woody plants seedlings under waterlogging stress [D]. Nanjing: Nanjing Forestry University, 2014: 40–44.
王哲宇. 九种木本植物幼苗的淹水胁迫生理特征研究 [D]. 南京: 南京林业大学, 2014: 40–44.
[13] ZHAO B, ZHANG F, XUE C R. Antioxidant properties ofL. seedling leaves under NaCl stress [J]. Shandong Agric Sci, 2016, 48(11): 57–60. doi: 10.14083/j.issn.1001-4942.2016.11.013.
赵兵, 张凡, 薛春荣. NaCl胁迫下木芙蓉幼苗叶片的抗氧化特性 [J]. 山东农业科学, 2016, 48(11): 57–60. doi: 10.14083/j.issn.1001- 4942.2016.11.013.
[14] TONG L L, WANG Z Y, XU X G. Effects of flooding stress on morphological & physiological property of[J]. J Beihua Univ (Nat Sci), 2020, 21(2): 253–256. doi: 10.11713/j.issn. 1009-4822.2020.02.025.
童丽丽, 王哲宇, 许晓岗. 淹水胁迫对木芙蓉幼苗形态及部分生理指标的影响 [J]. 北华大学学报(自然科学版), 2020, 21(2): 253–256. doi: 10.11713/j.issn.1009-4822.2020.02.025.
[15] LI H S. Experimental Principle and Technology of Plant Physiology and Biochemistry [M]. Beijing: Higher Education Press, 2003: 10–78.
李合生. 植物生理生化实验原理和技术 [M]. 北京: 高等教育出版社, 2003: 10–78.
[16] WANG Y H, GAO H W, WANG Z, et al. Study and evaluation of drought resistance capacity ofgermplasm resources at seedling stage [J]. J Plant Genet Resour, 2009, 10(3): 443– 447. doi: 10.13430/j.cnki.jpgr.2009.03.012.
王艳慧, 高洪文, 王赞, 等. 胶质苜蓿种质资源苗期抗旱性综合评价 [J]. 植物遗传资源学报, 2009, 10(3): 443–447. doi: 10.13430/j. cnki.jpgr.2009.03.012.
[17] XIE Z J. Fuzzy Mathematical Methods in Agricultural Science [M]. Wuhan: Huazhong University of Science and Technology Press, 1983: 99–193.
谢志坚. 农业科学中的模糊数学方法[M]. 武汉: 华中理工大学出版社, 1983: 99–193.
[18] LIU X Q, SUN X L, LIU Y S, et al. Spatial division of climate change and its evolution characteristics in southwest China based on REOF- EEMD [J]. Geogr Res, 2020, 39(5): 213–230. doi: 10.11821/dlyj02019 0387.
刘晓琼, 孙曦亮, 刘彦随, 等. 基于REOF-EEMD的西南地区气候变化区域分异特征 [J]. 地理研究, 2020, 39(5): 213–230. doi: 10. 11821/dlyj020190387.
[19] LI Q H, LIU S P, ZHI C Y, et al. Adaptation mechanism of three herbs in the water-level-fluctuation-zone of reservoir to complete submer- gence [J]. J Trop Subtrop Bot, 2013, 21(5): 459–465. doi: 10.3969/j. issn.1005-3395.2013.05.013.
李秋华, 刘送平, 支崇远, 等. 三种水库消落带草本植物对完全水淹的适应机制研究 [J]. 热带亚热带植物学报, 2013, 21(5): 459–465.doi: 10.3969/j.issn.1005-3395.2013.05.013.
[20] CHEN F Q, HUANG Y Z, ZENG X, et al. Biological response of cynodon dactylon vegetative propagule to simulated flooding [J]. J Trop Subtrop Bot, 2010, 18(1): 15–20. doi: 10.3969/j.issn.1005-3395. 2010.01.003.
陈芳清, 黄友珍, 曾旭, 等. 狗牙根营养繁殖体对模拟水淹的生物学响应 [J]. 热带亚热带植物学报, 2010, 18(1): 15–20. doi: 10. 3969/j.issn.1005-3395.2010.01.003.
[21] VOESENEK L A C J, BAILEY-SERRES J. Flood adaptive traits and processes: An overview [J]. New Phytol, 2015, 206(1): 57–73. doi: 10. 1111/nph.13209.
[22] LIU C C, LAN C J, LI H, et al. Responses of waterlogging stress and evaluation of waterlogging tolerance in cherry tomato at seedling stage [J]. J Nucl Agric Sci, 2020, 34(3): 650–660. doi: 10.11869/j.issn.100- 8551.2020.03.0650.
刘聪聪, 兰超杰, 李欢, 等. 樱桃番茄苗期对淹水胁迫的响应及其耐涝性评价 [J]. 核农学报, 2020, 34(3): 650–660. doi: 10.11869/j. issn.100-8551.2020.03.0650.
[23] SAFAVI-RIZI V, HERDE M, STÖHR C. RNA-Seq reveals novel genes and pathways associated with hypoxia duration and tolerance in tomato root [J]. Sci Rep, 2020, 10(1): 1692. doi: 10.1038/s41598-020-57884-0.
[24] CHO H Y, LORETI E, SHIH M C, et al. Energy and sugar signaling during hypoxia [J]. New Phytol, 2021, 229(1): 57–63. doi: 10.1111/ nph.16326.
[25] AYDOGAN C, TURHAN E. Changes in morphological and physiolo- gical traits and stress-related enzyme activities of green bean (L.) genotypes in response to waterlogging stress and recovery treatment [J]. Hort Environ Biotechnol, 2015, 56(3): 391–401. doi: 10. 1007/s13580-015-0127-9.
[26] PENG Y J, ZHOU Z X, ZHE Z, et al. Molecular and physiological responses in roots of two full-sib poplars uncover mechanisms that contribute to differences in partial submergence tolerance [J]. Sci Rep, 2018, 8(1): 12829. doi: 10.1038/s41598-018-30821-y.
[27] ELSE M A, JANOWIAK F, ATKINSON C J, et al. Root signals and stomatal closure in relation to photosynthesis, chlorophyll a fluore- scence and adventitious rooting of flooded tomato plants [J]. Ann Bot, 2009, 103(2): 313–323. doi: 10.1093/aob/mcn208.
[28] WANG X Y, SHI J H, LIU M X, et al. Effects of flood overtopping on leaf osmotic adjustment substances and antioxidant enzyme activities of naturalforest in the middle reaches of the Tarim River [J]. Arid Zone Res, 2020, 37(6): 1544–1551. doi: 10.13866/j.azr. 2020.06.20.
王新英, 史军辉, 刘茂秀, 等. 洪水漫溢对塔里木河中游天然胡杨林叶渗透调节物质及抗氧化酶活性的影响[J]. 干旱区研究, 2020, 37(6): 1544–1551. doi: 10.13866/j.azr.2020.06.20.
[29] WU J, WU J S. Effect of flooding stress on physiological and bio- chemical ofseedlings [J]. J NE For Univ, 2015, 43(4): 34–36. doi: 10.3969/j.issn.1000-5382.2015.04.008.
吴江, 吴家胜. 淹水胁迫对杨桐幼苗生理生化性质的影响[J]. 东北林业大学学报, 2015, 43(4): 34–36. doi: 10.3969/j.issn.1000-5382. 2015.04.008.
[30] ARBONA V, HOSSAIN Z, LÓPEZ-CLIMENT M F, et al. Antioxidant enzymatic activity is linked to waterlogging stress tolerance in[J]. Physiol Plant, 2008, 132(4): 452–466. doi: 10.1111/j.1399-3054. 2007.01029.x.
[31] WANY A, GUPTA K J. Reactive oxygen species, nitric oxide pro- duction and antioxidant gene expression during development of aerenchyma formation in wheat [J]. Plant Signal Behav, 2018, 13(2): e1428515. doi: 10.1080/15592324.2018.1428515.
[32] MA R J, ZHANG B B, CAI Z X, et al. Evaluation of peach rootstock waterlogging tolerance based on the responses of the photosynthetic indexes to continuous submergence stress [J]. Acta Hort Sin, 2013, 40 (3): 409–416. doi: 10.16420/j.issn.0513-353x.2013.03.002.
马瑞娟, 张斌斌, 蔡志翔, 等. 不同桃砧木品种对淹水的光合响应及其耐涝性评价 [J]. 园艺学报, 2013, 40(3): 409–416. doi: 10. 16420/j.issn.0513-353x.2013.03.002.
Response ofto Waterlogging Stress and Comprehensive Evaluation of Waterlogging Tolerance
XU Qian1, WU Mengxi1, ZENG Xinmei2, JIANG Beibei1, JIA Yin1, LI Mengxia2, ZHANG Manyu1, ZHANG Lu1, CAI Lu2, HUANG Wenpei1, PAN Yuanzhi1*
(1. College of Landscape Architecture, Sichuan Agricultural University,Chengdu 611130, China; 2. Chengdu Botanical Garden, Chengdu 610083, China)
In order to excavate the waterlogging resistant germplasm resources of, the morphological, physiological and biochemical indexes of 13 cultivars were studied after 15 days of flooding, and build the evaluation system of waterlogging resistanceThe results showed that the variation trend and range of morphological and physiological indexes were different among cultivars, and the correlation of some indexes was significant (<0.05). The 14 indexes were attributed to morphological growth index, adventitious root morphological index, membrane lipid peroxidation index, root biomass index, antioxidant index and osmotic regulation index according to contribution rate. Cluster analysis conducted according to comprehensive waterlogging resistance evaluation value (D) showed that 13varieties could be divided into four waterlogging tolerance types, including very strong (3 varieties), stronger (5 varieties), medium (3 varieties) and none waterlogging tolerance (2 varieties). Therefore, the plant height, adventitious root number, adventitious root length could be used as rapid and simple indexes to identify the comprehensive waterlogging tolerance at seedling stage in
; Waterlogging; Waterlogging tolerance; Membership function; Comprehensive evaluation
10.11926/jtsb.4428
2021-04-16
2021-06-07
四川省重点研发项目(2020YFN0004)资助
This work was supported by the Key Research and Development Projects of Sichuan Province (Grant No. 2020YFN0004).
徐倩(1992~ ),女,在读博士研究生,主要从事园林植物的培育和应用研究。E-mail: xq713720@163.com
. E-mail: scpyzls@163.com
