杉木人工林土壤微团聚体中铁铝氧化物与微生物的分布及其关系
2022-04-02黄荣珍王金平朱丽琴黄国敏申思佳李燕燕万鸿宇吴昊泽
黄荣珍, 王金平, 朱丽琴, 黄国敏, 申思佳, 李燕燕, 万鸿宇, 吴昊泽
(南昌工程学院 江西省退化生态系统修复与流域生态水文重点实验室, 江西 南昌 330029)
目前,中国亚热带南方红壤丘陵区经过30 a多的生态恢复,取得一定成就,但土壤退化问题依然严重,其核心是土壤结构的退化。土壤团聚体作为土壤结构的基本单位,它在保持土壤肥力,调节土壤通气透水性、持水性,维持土壤微生物多样性方面具有十分重要的作用,其稳定性是反映土壤结构的重要指标[1-2]。其中,微团聚体作为团聚体的更小单元,它更加稳定,是大团聚体形成的基础,它在营养元素的保持,供应及转化等方面也发挥着重要作用,对土壤有机碳的长期储存和稳定起决定性作用[3-4]。
土壤中的胶结物质是土壤最活跃的组分之一,它分为有机胶体和无机胶体,它们相互作用复合形成微团聚体。铁铝无机氧化物属于有机胶结物质,其表面常具有不同类型和数量的电荷,其电荷可吸附在黏粒表面形成氧化物胶膜,促进土粒更加紧密,形成稳定的团聚体,因而,它在团聚体的形成中起重要作用[5],且相对于大团聚体,它们更多的是作用于微团聚体。钙键复合体、铁铝键复合体是土壤中主要的有机矿质复合体,对微团聚体的形成也起重要作用[6]。目前,国内外关于铁铝氧化物和化学结合态有机碳的研究主要集中在大团聚体,而对于红壤微团聚体关注较少。且不同形态铁铝氧化物结晶度、比表面积、官能团结构、表面电荷等特性,导致其对腐殖质和黏土矿物的胶结能力及团聚体影响均不同[7],在有机无机复合过程中发挥的作用也不同,因此,研究各粒级微团聚体中不同形态铁铝氧化物和化学结合态有机碳在土壤中的垂直分布对于揭示微团聚体形成及其稳定机制具有重要作用。
微生物是土壤的重要组成部分,它在团聚体的形成中也发挥重要的作用。微生物在代谢过程中产生的聚合物如多糖、多糖醛、氨基酸等能够对土壤颗粒产生黏结作用,有些微生物本身的菌丝会对土壤颗粒起到缠绕作用,且菌丝会产生分泌物,这些作用均有利于团聚体的形成[8-9]。虽然已有研究显示,微生物中细菌主要存在于微团聚体中,而真菌主要存在于大团聚体中[10],但是红壤中各级微团聚体中微生物群落的分布研究尚不足。且铁铝氧化物和化学结合态有机碳同样对团聚体的形成起重要作用,但是团聚体中铁铝氧化物和化学结合态有机碳与微生物群落的关系也尚未报道。
杉木林是中国南方红壤的典型林分类型之一,但是当前面临土壤退化、生物多样性降低、林分生产力下降等问题。近年来,众多学者围绕杉木林土壤展开了大量研究,包括土壤团聚体、铁铝氧化物、有机碳、微生物等[11-12],但是这些研究并未关注到微团聚体中的铁铝氧化物和微生物。因此,本研究以杉木人工林土壤为调查对象,通过测定杉木林不同土层不同粒级微团聚体胶结物中铁铝氧化物含量、化学结合态有机碳含量和微生物群落多样性,研究它们在微团聚体中的分布,并揭示铁铝氧化物含量和化学结合态有机碳含量与微生物群落多样性的关系,为土壤退化修复提供理论依据。
1 研究区概况
研究区位于江西赣江源国家级自然保护区,在武夷山脉西部洋地林场(26°2′56″N,116°19′29″E),海拔435 m,坡度20°~30°,坡向东南。土壤成土母质主要为石英岩类风化物,土壤类型主要为红壤,pH值为4.5~5.5。该区地处中亚热带与南亚热带过渡区,年平均气温18.5 ℃,年均降雨量2 180 mm,年均日照数1 993 h,全年无霜期249 d,四季分明,属典型亚热带湿润季风气候。样地设置在自然保护区的试验区,为实施水土保持生态修复的对象。选择的林分是杉木人促更新林,林龄约15 a。由于受人为干扰严重,立地条件较差,以及土壤养分限制,森林发育不良,林相残败,退化严重。土壤均呈酸性,沿土层深度增加而酸性略有减弱,pH值为4.58~5.34。林地内选取的样地平均坡度为28°,呈东南坡向,主要树种为杉木(Cunninghamialanceolata)和马尾松(Casuarinaequisetifolia),平均树高4.13 m,平均胸径5.78 cm,灌木层盖度为10%,草本层盖度为78%,乔木郁闭度为32%。
2 材料与方法
2.1 样品采集与处理
在杉木林分别设立3块20 m×20 m固定标准地进行采样,每个标准样地相隔500 m以上。于2019年4月在每个标准地坡面的上、中、下3部分布设3个取样点挖土壤剖面,分别用不锈钢饭盒采集0—10,10—20,20—30 cm土层原状土。该林地由于林龄达15 a,草本层覆盖度高,因此土层厚度达30 cm以上,且表层并未流失。
将3个采样点同一土层的原状土混合作为1个土样,共9个土样。将采回的原状鲜土沿土壤自然结构脆弱带用手轻轻掰成直径约为1 cm的小土块,剔除植物残体和石块等杂物,并混合均匀。采用四分法取土样过2 mm筛备用,一部分置于4 ℃冷藏冰箱密封保存待处理,剩余小部分土样自然风干后过1 mm筛用于测定土壤相关化学指标。测定土壤有机质、全氮、全磷、全钾、有效磷和速效钾作为背景值,结果见表1。

表1 洋地林场杉木人工林不同土层土壤化学指标背景值
2.2 指标测定
土壤化学指标采用常规方法测定[13],有机质用重铬酸钾外加热法测定,全氮用全自动凯氏定氮法测定,全磷用碱熔—钼锑抗比色法测定,有效磷用NaHCO3浸提—钼锑抗比色法测定,全钾用碱熔—火焰光度计的方法测定,速效钾则用乙酸铵浸提—火焰光度计的方法测定。
土壤微团聚体胶结物分离采用沉降虹吸法,分离出粒级为<20,20~50,50~200 μm的微团聚体胶结物用于铁铝氧化物、结合态有机碳和微生物群落多样性的测定。
土壤微团聚体铁铝氧化物中,游离态铁铝氧化物(Fed和Ald)用连二亚硫酸钠—柠檬酸钠—重碳酸钠提取,非晶型铁铝氧化物(Feo和Alo)用草酸铵—草酸提取,络合态铁铝氧化物(Fep和Alp)用焦磷酸钠提取,然后用试铁灵铁铝联合比色法测定波长在600,370 nm下各提取液的吸光值[13]。
土壤微团聚体结合态有机碳中,钙键结合态有机碳(Ca-SOC)用0.5 mol/l的Na2SO4溶液提取,铁铝键结合态有机碳〔Fe(Al)-SOC〕用氢氧化钠(NaOH)和焦磷酸钠(Na4P2O7)混合液提取[14],提取液中的碳用碳氮分析仪(MultiN/C3 100)测定。
土壤微团聚体微生物群落多样性的测定采用磷脂脂肪酸法(PLFA)[15],首先土壤各粒级微团聚体用体积比为1∶2∶0.8的氯仿:甲醇:柠檬酸缓冲液震荡提取总脂类,然后通过SPE硅胶柱分离纯化得到磷脂脂肪酸,接着进行碱性甲脂化,用正己烷(色谱纯,HPLC)溶解待测样品,最后在Agilent 7 890 AGC气相色谱仪上测定,采用MIDI Sherlock软件系统(Version 4.5,MIDI,Inc,Newark,DE)鉴定PLFA的成分,使用的内标为正十九烷酸甲酯Nonadecanoic acid(C19∶0,溶于色谱正己烷)[16]。命名采用Frostegard方法命名[17]。
采用辛普森指数Simpson(D)、香农指数Shannon-Wiene(H)、均匀度指数(E)3种群落多样性指数来表征微生物群落物种数及其分布均匀程度的综合指标[18],计算方法如下:
Simpson(D)辛普森指数计算公式为:
Shannon-Wiene香农指数(H)计算公式为:
H=-∑PilnPi
(2)
Shannon均匀度指数(E)计算公式为:
E=-H/(lnS)
(3)
式中:Pi=Ni/N,Ni是指测得土样中第i种磷脂脂肪酸含量(PLFAs),N为测得土样中各类磷脂脂肪酸含量的总和;S是磷脂脂肪酸生物标记种类的总数,即为丰富度。
2.3 数据处理
用SPSS 17.0软件对数据进行统计,用双因素方差分析(two-way ANOVA)检验土层和粒级对微团聚体铁铝氧化物、结合态有机碳、微生物数量以及微生物多样性指数影响的显著性。用Origin 8.5进行制图,用Canoco 5.0进行微团聚体铁铝氧化物、结合态有机碳与微生物数量、微生物多样性指数之间的冗余分析。
3 结果与分析
3.1 不同粒级微团聚体中铁铝氧化物在土层中的分布
由图1可知,从总体上看,土壤中铁铝氧化物含量呈现:游离结晶态(Fed,Ald)≫无定形(Feo,Alo)>络合态(Fep,Alp)。对于铁的氧化物,通过方差分析可知,土层仅对络合态铁(Fep)含量有显著影响,而粒级对游离态铁(Fed)含量、非晶型铁(Feo)含量和Fep含量均有显著影响(表2)。50~200 μm微团聚体中的Feo含量和各粒级微团聚体中的Fep含量均随土层深度的增加下降明显(图1b和1c)。表层土壤(0—10 cm)中,Fed在<20 μm微团聚体中的含量要远高于其他粒级微团聚体(图1a),Feo在<20 μm和50~200 μm微团聚体中的含量要明显高于20~50 μm,Fep在<20 μm微团聚体中的含量最大,而在50~200 μm微团聚体中的含量最小。10—20 cm和20—30 cm的土层中,各形态铁的氧化物含量随着微团聚体粒级的减少而增大,其中,增大幅度最大的是Fed。
对于铝的氧化物,土层对游离态铝(Ald)含量和络合态铝(Alp)含量有显著影响,而粒级对各形态铝的氧化物含量均有显著影响(表2)。随着土层的增加,各粒级微团聚体中的Ald含量有下降趋势,与此相反,Alp含量则有上升趋势,且50~200 μm微团聚体中的Ald含量和各粒级微团聚体中的Alp含量在土层增加至20—30 cm时与表层(0—10 cm)差异极大(图1d和1f)。表层土壤(0—10 cm)中,各粒级微团聚体中铝的氧化物含量特别是Alp含量差异不大,10—20 cm的土层中,Ald和非晶型铝(Alo)在<20 μm微团聚体中的含量远高于其他粒级微团聚体(图1d和1e),而在20—30 cm的土层中,各形态铝的氧化物含量随着微团聚体粒级的变化规律与铁的氧化物类似。
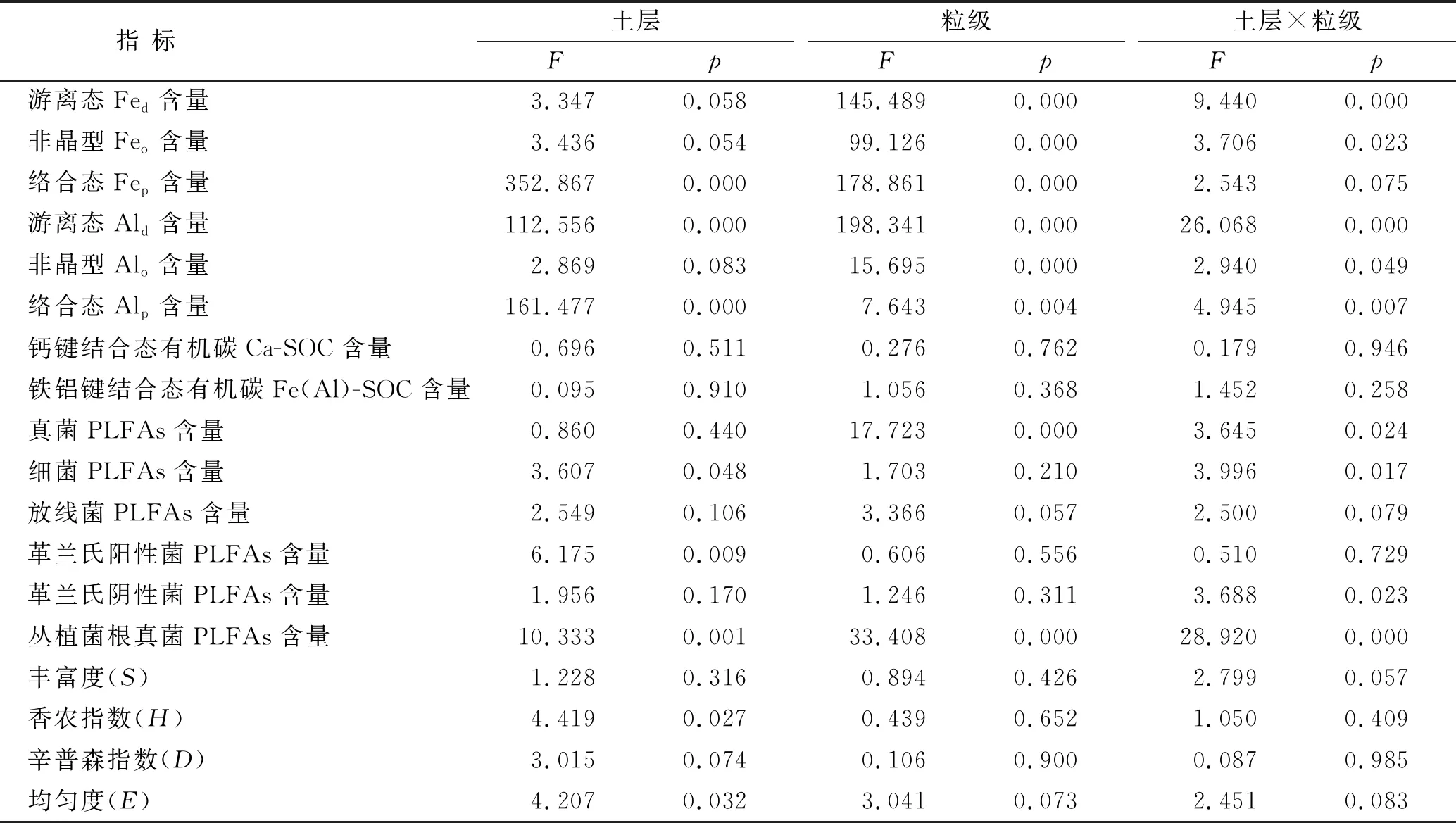
表2 杉木林土壤各指标的双因素方差分析
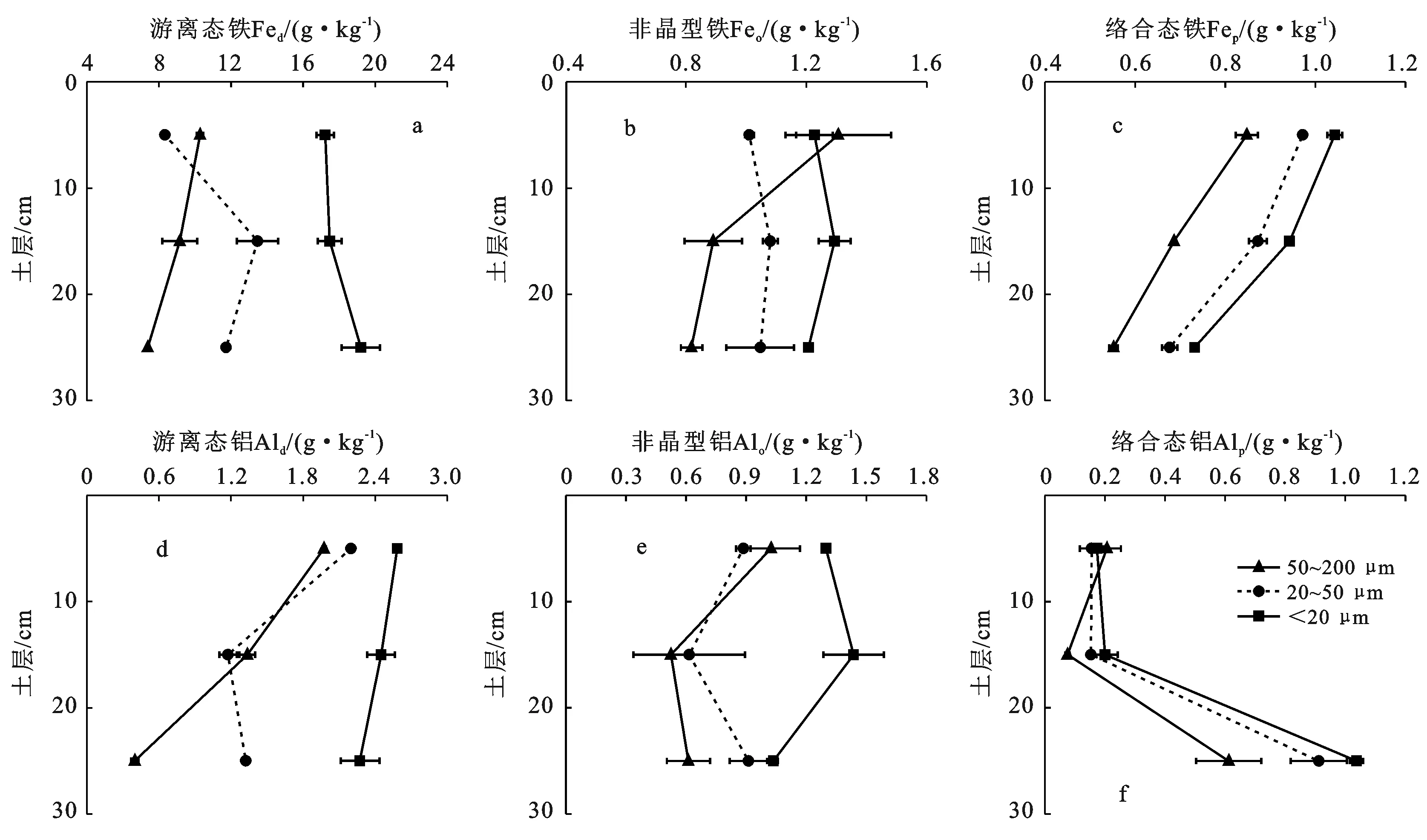
图1 杉木林土壤微团聚体中铁铝氧化物含量
3.2 不同粒级微团聚体中结合态有机碳在土层中的分布
由图2可知,从总体上看,土壤中铁铝键结合态有机碳〔Fe(Al)-SOC〕含量远远高于钙键结合态有机碳(Ca-SOC)。土层、粒级以及它们的交互作用(土层×粒级)对Ca-SOC和 Fe(Al)-SOC含量均无显著影响。各粒级团聚体Ca-SOC和Fe(Al)-SOC含量随着土层深度的增加无明显变化趋势。表层土壤(0—10 cm)中,<20 μm和20~50 μm微团聚体中的Fe(Al)-SOC含量要高于50~200 μm微团聚体(图2b)。
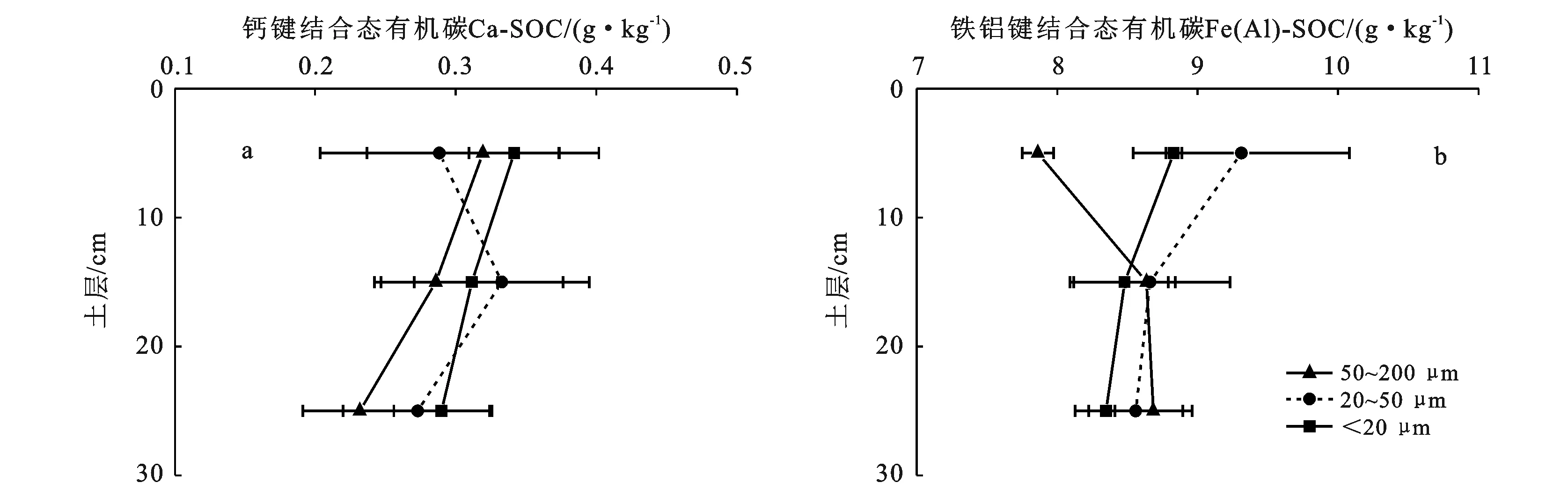
图2 杉木林土壤微团聚体中结合态有机碳的含量
3.3 不同粒级微团聚体中微生物PLFAs含量在土层中的分布
由图3可知,从总体上看,土壤中细菌和革兰氏阴性菌PLFAs含量最高,而真菌、放线菌和丛枝菌根真菌PLFAs含量最低。土层对细菌、革兰氏阳性菌和丛枝菌根真菌PLFAs含量均有显著影响,但粒级只对真菌和丛枝菌根真菌PLFAs含量影响显著(表2)。真菌PLFAs含量在各土层中尤其是在10—20 cm土层中随粒级的增加而增加(图3a)。细菌PLFAs含量随土层的变化与粒级有关,粒级为<20 μm的微团聚体中细菌PLFAs含量在土层增加至20—30 cm时大幅度降低,而粒级为20~50 μm和50~200 μm的微团聚体中细菌PLFAs含量在土层增加至20—30 cm时大幅度增加, 0—10 cm)和10—20 cm土层中,细菌PLFAs含量最高的是在50~200 μm微团聚体中,其次是<20 μm微团聚体中,最后是20~50 μm的微团聚体中,而20—30 cm土层中的细菌PLFAs含量随粒级的变化则刚好相反(图3b)。革兰氏阴性菌的变化规律与细菌相一致(图3b和3e)。各粒级微团聚体中的放线菌PLFAs含量只有在表层(0—10 cm)时差异较大,含量最高的是在50~200 μm微团聚体中,其次是<20 μm微团聚体中,最后是20~50 μm的微团聚体中,粒级为50~200 μm微团聚体中的放线菌PLFAs含量在土层下降至10—20 cm时就迅速下降(图3c)。各粒级微团聚体中的革兰氏阳性菌PLFAs含量在土层下降至10—20 cm时就迅速下降,表层(0—10 cm)土壤50~200 μm微团聚体中革兰氏阳性菌PLFAs含量远高于其他粒级(图3d)。同样的,表层(0—10 cm)土壤50~200 μm微团聚体中丛枝菌根真菌PLFAs含量远高于其他粒级,此粒级微团聚体中丛枝菌根真菌PLFAs含量随土层深度的增加迅速降低,而<20 μm微团聚体中丛枝菌根真菌PLFAs含量在土层增加至20—30 cm时略微增加(图3f)。
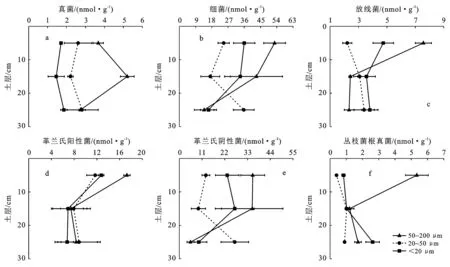
图3 杉木林土壤微团聚体中微生物PLFAs的含量
3.4 不同粒级微团聚体中微生物多样性指数在土层中的分布
土层对香农指数(H)和均匀度(E)均有显著影响,而粒级对微生物多样性指数均无显著影响(表2)。各粒级微团聚体中的微生物丰富度(S)只有在表层(0—10 cm)时差异较大,数值最高的是在<20 μm微团聚体中,其次是50~200 μm微团聚体中,最后是20~50 μm的微团聚体中(图4a)。表层(0—10 cm)土壤<20 μm微团聚体中的微生物香农指数(H)高于其他粒级,而20—30 cm土层20~50 μm微团聚体中的微生物香农指数(H)高于其他粒级,<20 μm和50~200 μm微团聚体中的微生物香农指数(H)在土层下降至10—20 cm和20—30 cm时降幅较大(图4b)。表层(0—10 cm)土壤各粒级微团聚体微生物辛普森指数(D)高于10—20 cm和20—30 cm土层(图4c)。表层(0—10 cm)土壤中,50~200 μm微团聚体中的微生物E要低于其他粒级微团聚体,其随土层深度的增加而有所增加(图4d)。
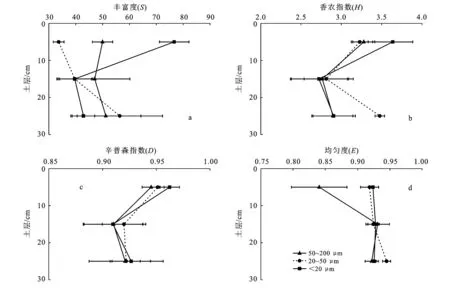
图4 杉木林土壤微团聚体中微生物多样性指数
3.5 土壤微团聚体中铁铝氧化物与微生物群落的关系
对土壤微团聚体中微生物群落及其多样性指数与各形态铁铝氧化物进行了冗余分析(RDA)(图5)。该模型pseudo-F=2.3,p=0.01<0.05,比较可靠,RDA轴1和轴2总共能解释所有变量的40.32%,说明微团聚体中各形态铁铝氧化物对微生物群落及其多样性的影响达到了40.32%。RDA结果显示,细菌、革兰氏阳性菌、革兰氏阴性菌是该林地的主要微生物。微生物种群与多样性的关系中,放线菌、革兰氏阳性菌PLFAs含量与香农指数、辛普森指数有较强的正相关关系,细菌PLFAs含量、真菌PLFAs含量、革兰氏阴性菌PLFAs含量与丰富度有较强的正相关关系,与均匀度指数有较强的负相关关系。微生物种群与铁铝氧化物相关指标的关系中,放线菌PLFAs含量、革兰氏阳性菌PLFAs含量与Fed含量显示出极强的负相关关系,细菌PLFAs含量与Ald含量、Alo含量显示出极强的正相关关系,与Alp含量显示出极强的负相关关系,丛枝菌根真菌PLFAs含量与Fe(Al)-SOC含量显示出极强的负相关关系,真菌PLFAs含量、革兰氏阴性菌PLFAs含量与Ald含量、Alo含量呈弱正相关关系。微生物多样性与铁铝氧化物相关指标的关系中,香农指数、辛普森指数与Fed含量显示出极强的负相关关系,丰富度指数与Ald,Alo,Fep和Ca-SOC含量呈强正相关关系,与Alp含量呈强负相关关系,而均匀度指数则与此相反。
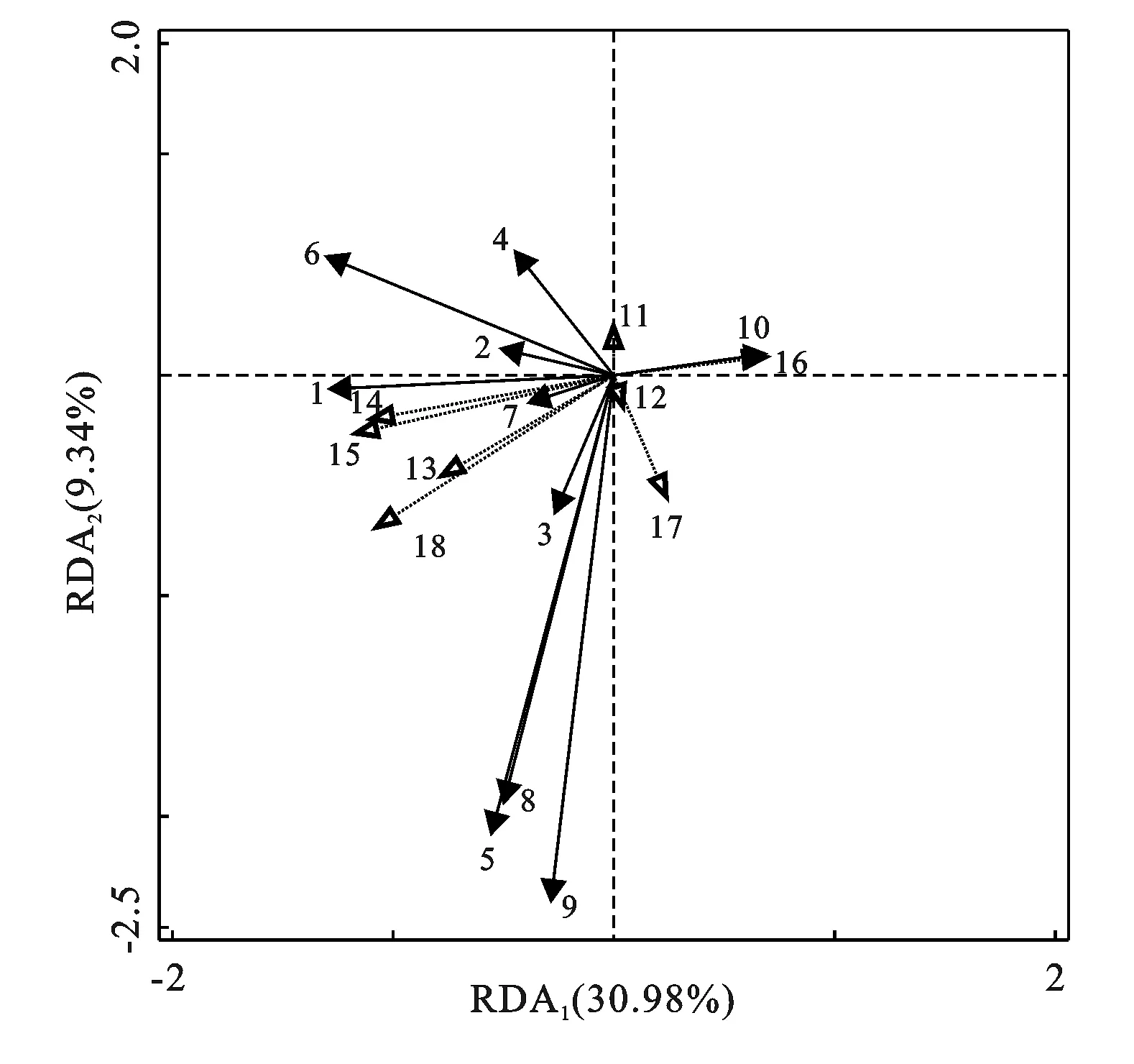
注:1细菌;2真菌;3放线菌;4丛枝菌根真菌;5革兰氏阳性菌;6革兰氏阴性菌;7微生物丰富度;8微生物香农指数;9微生物辛普森指数;10微生物均匀度;11游离态铁;12非晶型铁;13络合态铁;14游离态铝;15非晶型铝;16络合态铝;17铁铝键结合态有机碳;18钙键结合态有机碳;实心箭头是物种变量,空心箭头是环境变量。
4 讨论与结论
4.1 讨 论
一般而言,形成团聚体的胶结物质可分为有机胶体、无机胶体以及有机无机复合胶体,当土壤有机质含量较高且黏土矿物和铁铝氧化物含量较低时,有机胶体发挥主要作用,当有机质含量较低且黏土矿物和铁铝氧化物含量较高时,无机胶体和黏粒的内聚力发挥主要作用。章明奎等[19]认为大粒径团聚体的主要胶结物质是有机质等有机胶体,微团聚体的胶结物质是铁铝氧化物等无机胶体。铁铝氧化物在南方红壤团聚体的形成中扮演十分重要的角色。土壤铁铝氧化物中,游离态铁(Fed)铝(Ald)主要来源于成土母质,显著影响土壤理化特性各形态[20];土壤非晶形氧化铁(Feo)铝(Alo)相较于其他形态的铁铝氧化物拥有更大的比表面积和更密的羟基位点,对 SOC和有机氮的固持具有重要贡献;土壤络合态氧化铁(Fep)铝(Alp)参与土壤有机复合物的形成,引起金属离子,特别是铁、铝离子在土壤中的移动,在土壤剖面的发生和肥力的保持中具有重要的作用[21]。不同形态铁铝氧化物在土壤中的含量不同。王小红等[22]研究了中亚热带山区3种林分土壤铁铝氧化物含量的分布,发现3种林分土壤均呈现:游离结晶态(Fed,Ald)>无定形(Feo,Alo)>络合态(Fep,Alp)。与此类似,本研究中杉木林土壤游离态铁铝含量远远大于非晶型和络合态铁铝氧化物含量,这是由于南方红壤呈酸性,而酸性环境下的强还原条件更有利于结合态铁铝的大量溢出,导致游离态铁铝含量明显增加[23]。土壤中高含量的游离态铁铝说明杉木林土壤遭受的风化和淋溶的作用较强,土壤发育程度也较高[24]。铁铝氧化物在不同土层中含量有所不同。衡利沙等[25]研究黄棕壤中铁铝氧化物的分布发现铁铝氧化物含量一般随土层增加逐渐降低。本研究中,杉木林土壤Fep和Ald含量也随土层深度的增加而逐渐降低,表层土壤Ald含量更高的原因是表层土壤有凋落物及丰富的根系等作用,较深层土壤水分更多,有利于羟基铝的形成[26],而表层土壤Fep含量更高的原因则是由于络合态铁本身就是铁氧化物和有机质的胶结,表层土壤有机质含量更高,更加有利于络合态铁的形成[27]。而与此相反,20—30 cm的土层中的Alp含量远高于0—20 cm的土层,这可能的原因是0—20 cm的土层土壤酸性更高,导致结合态铁铝大量转化为Ald。铁铝氧化物含量一般随微团聚体粒径的减少而逐渐增大[25],本研究各形态铁铝氧化物含量总体趋势也遵循这一规律,但随土层深度增加,这一规律更加明显。
结合态有机碳通过钙键和铁铝键结合,一定程度上对有机碳起到保护作用,抵制微生物对有机碳的分解,减缓其在土壤中的周转速度,从而促进有机碳的积累[26],其中,铁铝键结合态有机碳〔Fe(Al)-SOC〕稳定性高于钙键结合态有机碳(Ca-SOC)。本研究中土壤中Fe(Al)-SOC含量远远高于Ca-SOC,这从侧面也印证了Fe(Al)-SOC稳定性高于Ca-SOC这一结论。研究表明,土壤微团聚体有机碳随微团聚体粒径的减少而增加[28-29],但是土壤微团聚体Fe(Al)-SOC和Ca-SOC随粒径的变化规律研究较少。本研究结果显示钙Ca-SOC含量在不同土层和不同微团聚体粒径下差异并不显著,而Fe(Al)-SOC在<20 μm和20~50 μm微团聚体中含量要高于50~200 μm微团聚体。
微生物在团聚体的形成和稳定中具有不可忽视的作用,细菌菌体本身的碳水化合物是团聚体形成的重要组成成分,其表面带着的负电荷可以通过静电引力吸附土壤颗粒,形成团聚体。菌根真菌会形成菌丝体,对土壤颗粒起到缠绕作用[30]。真菌及放线菌可以在土壤颗粒间形成菌丝桥,从而影响土壤团聚体的形成。微生物代谢产物如蛋白、孢外多糖、脂质、球囊霉素等是微团聚体胶结物的形成核心[31]。不同微生物类群在微团聚体中组成和分布不同,荣勤雷[32]研究发现菜田石灰性褐土微团聚体中真菌含量高于细菌,而本研究结果表明杉木林土壤微团聚体细菌PLFAs含量远高于真菌PLFAs含量,这与Gupta(2014)等的研究结果相一致。而在微团聚体的粒径层面上,根据张维等的报道,细菌和真菌主要分布在小于53 μm中,放线菌主要分布在53~200 μm粒级微团聚体中[33],本研究结果却显示细菌、真菌、放线菌、革兰氏阳性菌、革兰氏阴性菌和丛枝菌根真菌PLFAs含量均是在粒径为50~200 μm的微团聚体中含量最高,且不同粒径微团聚体中微生物的含量在表层土壤中差异最大。上述研究中不同种类微生物在土壤微团聚体中含量高低不一致的原因可能是受土壤酸碱性、通气透水性和土壤等的多重影响,因为不同种类微生物所需的酸碱环境、生存孔隙和碳源种类不同[34]。另外,杉木林土壤微团聚体中微生物丰富度、香农指数和辛普森指数总体上均是表层土壤最高,且在表层土壤中,粒径最小的微团聚体微生物丰富度、香农指数最高,主要原因是表层土壤通气性更好,有机碳含量更高,种类更丰富,为土壤微生物的繁殖创造了良好的条件[35]。
4.2 结 论
目前,关于土壤微团聚体中微生物群落多样性与土壤因子关系的研究报道较多,而与铁铝氧化物关系的研究报道较少。本研究中放线菌PLFAs含量、革兰氏阳性菌PLFAs含量与Fed含量显示出极强的负相关关系,细菌PLFAs含量与Ald含量、Alo含量显示出极强的正相关关系,这可能是由于放线菌、革兰氏阳性菌对酸性环境耐受力较弱,细菌对酸性环境耐受力较强,而红壤Fed和Ald含量越高,酸性也越强,因此,放线菌、革兰氏阳性菌PLFAs含量越低,细菌PLFAs含量越高。真菌、细菌、革兰氏阴性菌和丛枝菌根真菌PLFAs含量与Fe(Al)-SOC含量均呈负相关,说明这几类微生物菌无法利用与铁铝键结合而成的有机碳,这对于土壤中有机碳的积累有积极作用。
