无核葡萄杂交胚挽救后代花型与无核性状的遗传表现
2022-04-02罗燚佳张剑侠
罗燚佳,张剑侠
(西北农林科技大学园艺学院/旱区作物逆境生物学国家重点实验室/农业农村部西北园艺植物种质资源利用重点开放实验室,陕西杨凌 712100)
葡萄的花型有3种:两性花(完全花)、雌花和雄花[1]。野生葡萄一般为雌雄异株,在葡萄的长期进化和人类的选择作用下,栽培品种绝大多数为两性花,少数为雌花[2]。葡萄是自花授粉植物,也可异花授粉。对于两性花品种,由于不需要栽植授粉树,给生产带来了极大便利。而雌花品种,其产量每年依赖于花期气候条件而显著变动,在天气不良的年份会造成传粉困难,使产量大大降低[3]。无核葡萄由于无籽,食用方便,因而无核性状一直是葡萄育种的重要目标[4]。在常规的无核葡萄杂交育种中,只能以有核葡萄作母本,无核葡萄作父本杂交,后代中无核几率低,育种周期较长。1982年Ramming等[5]改变了传统育种模式,创立了无核葡萄胚挽救技术,解决了无核葡萄不能作母本的问题,提高了无核杂种比率,极大地缩短了育种年限。该技术已逐渐成为各国无核葡萄育种者采用的重要方法[6]。
关于葡萄的花型遗传[1,3,7-8]和无核遗传[9-16]有多种学说,但都存在有一定的局限性。在无核葡萄胚挽救育种中,为减少无核葡萄杂交胚挽救后代群体中雄株的数量及提高无核后代的比率,避免在亲本选择和选配中的盲目性,本文对本课题组前期获得的两个无核葡萄杂交胚挽救后代群体的花型及无核性状进行了田间调查,以期获得无核葡萄作母本杂交后代的花型及无核性状的遗传规律,为今后开展无核葡萄胚挽救育种提供理论依据。
1 材料与方法
1.1 试验材料
材料为2012年定植于西北农林科技大学葡萄种质资源圃中的两个无核葡萄杂交胚挽救后代群体,其中‘红宝石无核’为母本,与‘火焰无核’杂交的F1代46株;‘红宝石无核’为母本,与‘黑龙江实生’(雄花)杂交的F1代105株。
1.2 试验方法
2019年和2020年的4—5月对两个杂交组合的胚挽救后代进行多次花型调查,7—9月进行无核性状调查,第二年主要对上年未开花、未结果的杂种进行调查补充。花型鉴定参照贺普超的方法[1],雄花:花丝较长,子房不发育;雌花和两性花子房明显,雌花的花丝较短、花药位于柱头的下方,两性花花药明显位于柱头上方。无核葡萄的划分参照Mahanil等的方法[17],无核:葡萄胚珠质量在30 mg以下;残核:葡萄种痕30 mg至胚珠不变色、不硬化,为绿色的胚珠;硬核:葡萄中存在木质化褐色种子。根据调查结果统计分析花型和无核性状遗传,并进行X2测验和基因型的推定。
2 结果与分析
2.1 两个杂交组合胚挽救后代的花型遗传表现
结果(表1)显示,在双亲均为两性花的‘红宝石无核×火焰无核’杂交组合F1代46株中,两性花和雌花分别占63.04%和36.96%,未出现雄株。在母本为两性花、父本为雄花的‘红宝石无核×黑龙江实生’杂交组合F1代105株中,两性花、雌花和雄花分别占60.95%、36.19%和2.86%。
为分析花型遗传是否符合Levadoux的M、H、F三个等位基因学说,对花型表现结果进行了卡方检验[1](表1)。组合‘红宝石无核×火焰无核’为两性花与两性花杂交,后代出现性状分离,因此认为两者均为基因异质HF,且卡方检验结果表明理论值与实际值差异不显著,说明其符合三等位基因学说。另一组合‘红宝石无核×黑龙江实生’为欧亚种两性花品种与山葡萄雄株的种间杂交,‘红宝石无核’基因型为HF,‘黑龙江实生’基因型可能为MM、MH和MF,又由于后代中出现了雌花,因此推断‘黑龙江实生’基因型为MF,通过卡方检验发现其实际值与理论值差异显著,说明其不符合三等位基因学说。
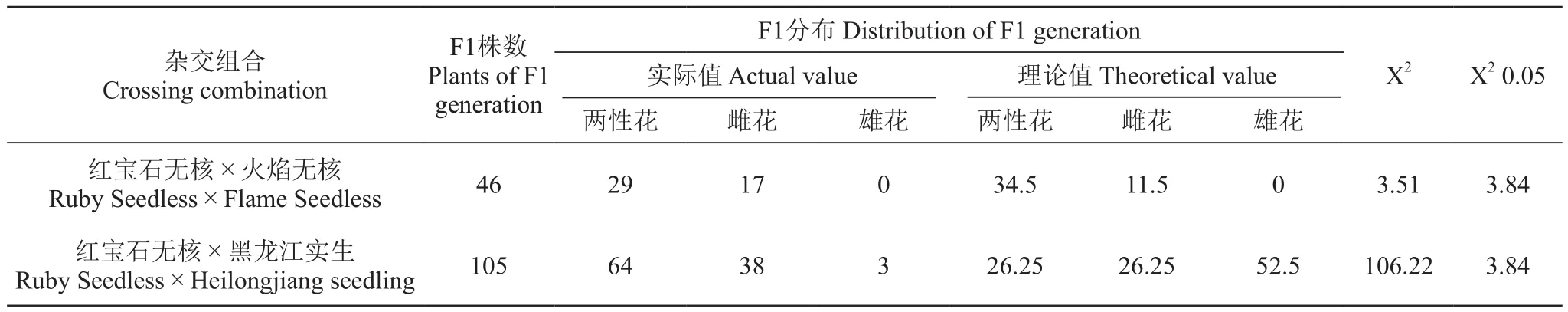
表1 两个组合后代花型遗传表现Table 1 The flower tуpe inheritance of progenies of the two combinations
2.2 两个杂交组合胚挽救后代的无核性状遗传表现
本研究依据杂种的胚珠质量和外观颜色与质地,确定核的性状为:无核、残核和有核。结果(表2)显示,两组合的F1代中,无核率分别达到50.00%和55.45%,说明以无核品种‘红宝石无核’作母本杂交,无论父本是否有核,无核性状都具有一定的遗传优势。
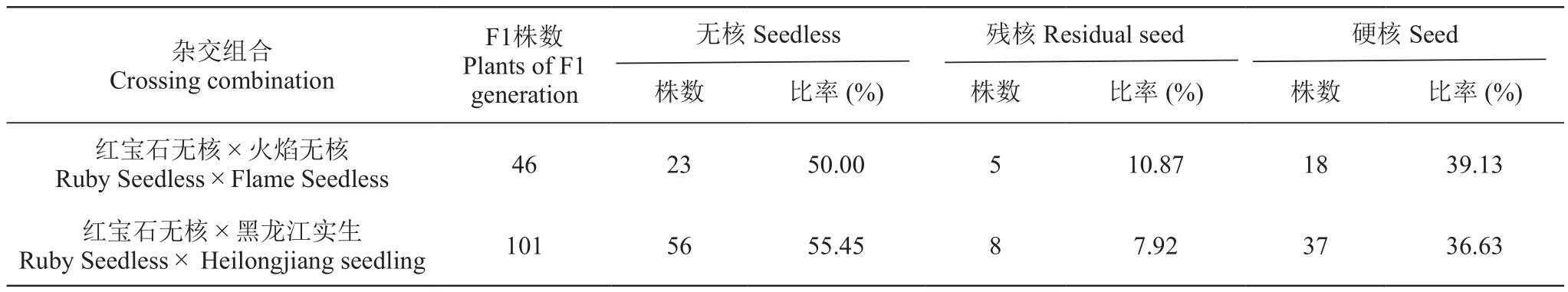
表2 两个组合后代的无核性状遗传表现Table 2 The seedless trait inheritance of progenies of two combinations
3 讨论
3.1 葡萄杂交后代的花型遗传
关于葡萄花型的遗传,贺普超[1]研究认为,欧洲葡萄品种间杂交,若父母本均为两性花,其后代大都为两性花,少数组合出现少量雌花;若为雌花×两性花,有的F1代为两性花,有的组合F1代有一部分雌花。用‘玫瑰香’为亲本,无论另一亲本性别如何,其后代都为两性花。这说明欧洲葡萄两性花性状具有一定的遗传优势。
Levadoux认为,葡萄花型受M、H、F三个等位基因控制。M决定雄花,H为两性花基因,F决定雌花。M对H、F为显性,H对F为显性。雄花基因型为MM、MH和MF,两性花基因型为HH和HF,雌花基因型为FF[1]。
本研究中,欧亚种品种间杂交‘红宝石无核×火焰无核’,为两性花×两性花,F1代两性花与雌花的比例为29∶17(2∶1),较为接近Levadoux的M、H、F三个等位基因学说,即两个杂合基因型的两性花亲本的杂交F1代中两性花与雌花比例接近3∶1。因此,推测‘红宝石无核’和‘火焰无核’两者基因型均为HF,F1代才会出现接近3∶1的分离比。
尹立荣等[7]对山葡萄花型遗传的研究表明:山葡萄不同花型杂交,子代中均出现雌花植株偏少的现象;山葡萄两性花与雄花植株杂交子代中,两性花植株数比理论值偏多。本研究中,‘红宝石无核×黑龙江实生’属于欧亚种两性花品种与山葡萄雄株的种间杂交,在后代中两性花植株偏多,这与尹立荣的结果相似。说明了以欧亚种两性花品种作母本与山葡萄雄株进行种间杂交时,后代中两性花和雌花占优势。
3.2 葡萄杂交后代的无核性状遗传
关于葡萄无核性状遗传假说有多种,包括:一隐性基因,认为无核性状是由单个隐性基因控制[9-10],但是解释不了无核×无核以及无核自交F1代中存在的有核性状。二隐性基因,Spiegel-Roy等[11]通过5年的试验发现,无核×无核的F1代中有核与无核的比例接近3∶1,所以假定两个互补的隐性因子决定了葡萄无核。几隐性基因,认为无核的产生受多个隐性基因的共同控制[12-13]。数量性状因子[14],解释不了无核与有核相互芽变的产生。五显性基因,来源于‘汤普森无核’或‘莫努卡’的品种无核性状由5个显性基因控制[15]。三显性基因,认为由3个优势互补因子组成的雌性无性系遗传模型,3个独立遗传的基因是产生无核表型所必需,有核性状的产生是由于3个基因中的任意一个处于隐性纯合[16],但是这一假说无法说明有核个体自交F1代会产生无核的现象。一显性基因,Stout[17-19]根据多重互补原理研究无核遗传规律,利用‘康可无核’的有核F1代相互杂交,认为无核性状是由一个显性基因控制。一显性调控基因与三隐性基因的复合调控,即由1个显性基因调控3个独立遗传的隐性基因表达来控制葡萄的无核性状[20],随后Lahogue等[21]通过无核基因分子标记证实1个主效基因和其它互补的隐性基因的存在。Doligez等[22]使用两个无核亲本杂交的139个F1代个体,构建遗传图谱,提出有3个微效基因控制和1个主效基因控制葡萄的无核性状,将种子质量等性状定位在一个连锁群上,证明了由微效基因和主效基因相互协同控制葡萄无核。此外,Cabezas等[23]对种子的鲜质量及浆果的种子数和种痕数进行分析,认为无核是数量性状遗传。无核性状的形成原因多种多样,其中包括:花粉不育、胚囊败育、胚乳败育[24]。但迄今尚未有一个完美的学说解释清楚葡萄无核性状的遗传。
王勇等[25]通过调查‘红宝石无核×SP6164’杂交F1代134株无核性状,发现75株为无核,约占调查后代中的56%,并认为杂交组合中的无核遗传优势较强。董新平[26]调查了4650株有核×有核和有核×无核的F1代,在有核×无核组合中后代无核占比最高不超过9.7%。王勇等[27]以有核品种‘红地球’为母本,无论其与有核还是无核父本杂交,其后代无核率均低于10%。
本研究中,‘红宝石无核×火焰无核’为无核与无核杂交,F1代中无核率为50.00%;‘红宝石无核×黑龙江实生’为无核与无果(按有核)杂交,F1代中无核率为55.45%。这表明当母本为无核葡萄时,父本无论是有核或者无核,其F1代都表现出较高的无核率,说明无核品种作母本无核性状具有遗传优势,这与王勇等[25]的调查结果接近。对于‘红宝石无核×火焰无核’F1代无核率低于‘红宝石无核×黑龙江实生’,可能是由于该组合的样本较小,后代有所缺失导致。
4 结论
欧亚种无核葡萄种内杂交‘红宝石无核×火焰无核’胚挽救F1代花型遗传基本满足Levadoux的M、H和F三个等位基因学说,且双亲基因型均为异质HF;‘红宝石无核’与山葡萄‘黑龙江实生’种间杂交F1代花型遗传不符合三个等位基因学说,两性花和雌花占有优势。以欧亚种无核品种为母本,无核性状在杂交F1代中具有一定的遗传优势。
