核麦间作条件下小麦光合特性及核桃果实品质的变化
2022-04-01朴涵琪余晓娥王宝庆虎海防
朴涵琪,余晓娥,欧 源,王宝庆,陈 虹,虎海防
(1.新疆农业大学林学与园艺学院,乌鲁木齐 830052;2.新疆林业科学院,乌鲁木齐 830000)
0 引 言
【研究意义】阿克苏地区是新疆核桃(JuglansregiaL.)的主产区[1],占全疆核桃总种植面积的37.5%,该地区也是小麦、棉花等农作物的主要栽植地。间作在时间和空间上实现集约化种植的栽培,能提高单位面积的光能利用率,并充分利用到温,水,肥等资源,提高单位面积产出效率[2]。随核桃树龄不断增加,树冠结构发生变化,林木与农作物之间的资源竞争也更加明显[3]。植物通过光合作用,利用太阳能合成自身所需的碳水化合物[4],光合同化产物也是植物果实产量与品质形成的物质基础[5],植物受到不利的光照势必会影响植物的产量[6-7]。研究阿克苏地区核桃林内光环境变化、间作物光合生理指标及单作和核麦2种模式下核桃果实的品质差异,对核桃与小麦丰产增收有实际意义。【前人研究进展】间作系统内光环境变化所导致的间作物生长发育受阻以及主作物的光合性能产生差异是间作系统发生不利竞争的关键因素[8,9]。【本研究切入点】测定单作园与间作园内光环境的变化、间作物的光合生理指标以及分析单作与间作两者间核桃果实品质的差异。【拟解决的关键问题】使用LAI-2200C冠层分析仪和LI- 6400便携式光合仪,研究2种种植模式下核桃的果实品质以及对比单间作小麦生理指标差异,为核桃与小麦的丰产增收提供理论依据。
1 材料与方法
1.1 材 料
试验地选择在新疆阿克苏地区温宿县佳木试验站(E80°31′58″,N41°15′22″),地处天山中部托木尔峰南麓,塔里木盆地西北的边缘,台兰河灌区上游,属于大陆性暖温带干旱气候,年降水稀少,季节分配不均,昼夜温差大。春季升温快而不稳,秋季短暂降温迅速,光照充足。4~8月中旬,平均气温22.3℃,日最高气温36.1℃,日最低气温6℃,08:00~20:00,总降水量为51.3 mm,平均相对湿度为35.7%,平均风速达到3.79 m/s,平均气压为872.5 hpa。
供试对象为核麦间作园复合系统与核桃单作园单一生产系统,核桃品种为早实丰产的温185号(J. regia Wen185)树龄6a,平均株高6.48 m,东西冠幅5.57 m,南北冠幅6.03 m,株行距(4×5) m,南北行向栽植。冬小麦品种为当地主栽品种新冬2号,播种行距15 cm。实测间作物冬小麦不同生育时期的光合生理参数以及2种生产系统下核桃树的叶面积指数(LAI)与核桃果实的外观性状、营养成分。间作园与单作园均按当地常规田间管理方法进行施肥及灌溉管理。
1.2 方 法
1.2.1 叶面积指数测定
在间作园与单作园内分别选取两列长势基本一致,林相整齐的核桃树,使用LAI-2200冠层分析仪进行大田内核桃树的叶面积指数(LAI)的测定。将探头放置在空旷地矫正,测定A值,之后放于林下距离底层树叶大致树叶4倍大的距离,保持探头上水平泡平衡,按下测定按钮,听到仪器发出两声蜂鸣后即可,测量点首先选择2行8株核桃树(每行4株),以对角线为主测量4次,然后以最后1株核桃树为起点测此行剩余3株核桃树,则仪器自动测出群体叶面积指数(LAI)。图1

图1 叶面积指数测定示意Fig.1 schematic diagram of leaf area determination
1.2.2 冬小麦光合生理数据
采用美国Li-COR公司生产的便携式光合仪Li-6400XT及其配备的LED人工光源测定冬小麦的光合生理数据。分别于冬小麦的抽穗期(5月上旬)、扬花期(5月中旬)、灌浆期(5月下旬)、成熟期(6月中旬)的10:00~12:00(晴朗无云)测定冬小麦旗叶在光合光量子通量密度13个梯度(1 800、1 500、1 200、1 000、800、600、400、250、200、150、100、50、0单位)下的净光合速率Pn、Ci、Gs、WUE等参数,并绘制出Pn光响应曲线。设定3个重复,取平均值。
1.2.3 核桃果实性状
待果实成熟后,分别采集单作园、间作园果实。以“Z”字形为标准选取20株核桃树,每株采摘10颗果实,游标卡尺测量干果三径,并测定脂肪、蛋白质、氨基酸、皂化值、过氧化值、出仁率、单宁,其中,氨基酸采用GB 5009.124-2016、脂肪采用GB 5009.6-2016、皂化值采用GB/T 5534-2008、过氧化值采用GB 5009.227-2016、蛋白质采用GB 5009.5-2016、出仁率采用GB/T 20398-2006、单宁采用GB/T 15686-2008(比色法)[10-11]。
1.3 数据处理
冬小麦光合作用光响应曲线拟合采用非直角双曲线模型[12-13]。
(1)
式中:Pn为净光合速率[μmol/(m2· s)];l为冬小麦叶片接受到的光量子密度,也称作初始量子密度(μmol ·CO2/μmol·PPFD),α为表征光合作用最大光能转化率的表观初始量子效率(μmol ·(CO2)/μmol);Pmax 为冬小麦的最大净光合速率[μmol/(m2· s)];Rd为暗呼吸速率;θ为非直角曲线的凸度。通过冬小麦的光响应曲线可以得出冬小麦的光饱和点和光补偿点[14]。
采用Microsoft Excel 2010,SPSS17.0处理试验中的数据。
2 结果与分析
2.1 不同种植模式下叶面积指数的差异
研究表明,不同模式下叶面积指数存在极显著差异(P<0.01),间作园与单作园叶面积指数整体变化趋势都是先增大后逐渐变小,单作园叶面积指数均高于间作园叶面积指数。二者在速生生长期叶面积指数最大,其中单作园叶面积指数为4.242,间作园叶面积指数为3.055。单作园叶面积指数在展叶期为最低,仅为2.928,而间作园则在近成熟期表现为叶面积指数最低,为2.118。图2

注:不同字母表示为差异显著(P<0.01)
2.2 冬小麦不同生育时期光合生理参数
研究表明,冬小麦在抽穗期、扬花期、灌浆期、成熟期的光响应曲线趋势基本保持一致,并且不同生育时期的净光合速率(Pn)差异较大,在相同的环境条件下,抽穗期的净光合速率值(Pn)较高。当光合有效辐射强度处于0~300 μmol/(m2·s)时,净光合速率(Pn)随光合有效辐射强度的增加呈直线性增长。当光合有效辐射强度达到300 μmol/(m2·s)以上时,随着PAR的增加,间作物冬小麦的抽穗期、扬花期、灌浆期、成熟期的净光合速率上升趋势变缓。当光合有效辐射强度达到1 300 μmol/(m2·s)时,净光合速率呈现一种稳定状态。图3

图3 冬小麦不同生育时期光响应曲线Fig.3 Changes of net photosynthetic rate of winter wheat at different growth stages
2.2 冬小麦不同生育时期光饱和点和光补偿点
研究表明,冬小麦不同生育期的光饱和点大小差异明显。总体呈现逐渐升高又减小的变化趋势。在各个生育期中,抽穗期、灌浆期光饱和点存在极显著差异(P<0.01);而扬花期与成熟期的光饱和点差异不显著。其中抽穗期的光饱和点仅为818.581 2 μmol/(m2·s);灌浆期的光饱和点可达2 592.455 1 μmol/(m2·s);光补偿点的变化趋势则呈现出先增大后减小再增大的特点。4个生育时期差异不显著,其中抽穗期的光补偿点最低,为20.685 5 μmol/(m2·s),成熟期的光补偿点为47.151 6 μmol/(m2·s)。表1

表1 冬小麦不同生育期光饱和点与光补偿点Table 1 Light saturation point andlight compensation point of winter wheat at different growth stages
2.3 不同生育时期冬小麦的光合生理参数变化
研究表明,不同生育时期冬小麦的光合参数存在明显的变化趋势,不同生育期的胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)、水分利用效率(WUE)均存在规律性的变化。胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)是影响植物叶片净光合速率的主要原因之一,净光合速率Pn的大小直接决定了植物进行光合作用的程度。Ci、Gs、Tr等相互配合,协同作用从而使光合作用顺利进行。不同生育时期冬小麦的胞间CO2浓度、气孔导度发展呈相反趋势,气孔导度(Gs)随着光合光量子通量密度的增大而增大,胞间CO2浓度则随着光合光量子通量密度增大而减小。灌浆期气孔导度值略大于其他时期,成熟期气孔导度最小。抽穗期、扬花期、成熟期的Ci变化不太明显,灌浆期的胞间CO2浓度则略低于其他3个时期。图4~5

图4 不同生育期冬小麦气孔导度变化Fig. 4 Changes of stomatal conductance of winter wheat at different growth stages

图5 不同生育期冬小麦胞间CO2浓度变化Fig. 5 Changes of Ci of winter wheat at different growth stages wheat in different growth stages
研究表明,冬小麦不同生育期的蒸腾速率与水分利用效率变化趋势同样大体一致,不同时期冬小麦的蒸腾速率随着光合光量子通量密度的增加而呈现上升趋势,不同的是抽穗期在光合光量子通量密度到达500 μmol/(m2·s)时,蒸腾速率趋于平稳,在扬花期光合光量子通量密度在1 000 μmol/(m2·s)时,蒸腾速率出现短暂下降并于1 500 μmol/(m2·s)时又出现了缓慢的上升。水分利用效率(WUE)的变化则呈现先增大后平稳的趋势,扬花期水分利用效率最大,并且扬花期光合光量子通量密度在800 μmol/(m2·s)时出现下降在达到1 000 μmol/(m2·s)时又出现升高趋势,最终平稳。图6~7
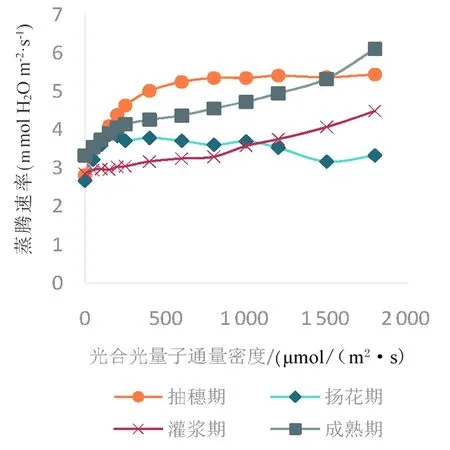
图6 不同生育期冬小麦蒸腾速率变化Fig. 6 Changes of transpiration rate of winter wheat in different growth stages

图7 不同生育期冬小麦水分利用效率变化Fig.7 Changes of water use efficiency of winter wheat in different growth stages
2.4 核桃果实的外观型状与品质性状
研究表明,单作园与间作园模式下核桃果实的外观性状差异较不明显,只有鲜果重、纵径、去皮后纵径以及去皮后的横径存在显著性差异,其余差异均不显著。表2
核桃果仁品质在不同栽植模式下存在差异,单作园核桃果仁的棕榈酸、硬脂酸、油酸、亚油酸、亚麻酸、皂化值、蛋白质以及氨基酸的含量要极显著高于间作园果仁的含量,单作园果实脂肪含量显著高于间作园脂肪含量,而间作模式下的果仁单宁含量则极显著高于单作模式。表3

表2 不同种植模式下核桃果实外观性状变化Table 2 Effects of different planting patterns onthe appearance and characters of walnut fruit

表3 不同种植模式下种仁营养成分变化Table 3 Effects of different planting patterns on the nutritional composition of seed kernels
3 讨 论
间作条件下,间作物与主作物同在一个种植系统内,因生态位的交叠,势必导致该系统内的资源配置与单作环境存在差异。叶面积指数(LAI)是群体结构的重要量化指标[15]。通常叶面积指数(LAI)有一个最适值,该值是当光通过叶冠被均匀吸收时的叶面积。当该植物测定的LAI大于其最适值时,叶面积指数过大,透光率变小,光截获密度则变小,相应的会影响植物进行光合作用[16]。当该植物测定的LAI小于其最适值时,LAI越小则证明其叶面积越小,叶面积大小直接关系到林分同化光能的数量,关系到生产力。研究中,间作模式与单作模式下核桃叶面积指数(LAI)存在极显著差异(P<0.01),各个时期的叶面积指数均表现为单作条件高于间作条件。核桃的LAI最适值在3~6[17]。研究中间作条件下的核桃的叶面积指数在2~3,而单作条件下的叶面积指数则处在2.5~4.5。单作叶面积指数处在LAI的最适值范围内,单作核桃园可以最有效的通过叶片进行光合作用,积累有机物。间作的叶面积指数小于核桃LAI的最适值,间作园内核桃的叶面积较小,不能完全进行光合作用,对光合同化产物的产生造成一定影响,进而影响了果实品质[18]。核桃的速生生长期和油脂转化期的叶面指数高于其余两时期,此阶段核桃的LAI值迅速增长,此时期为核桃产量形成的重要时期,光合作用强,营养物质积累迅速。而进入成熟期后,LAI值下降,这可能与核桃进入成熟期,光合作用减弱,叶片产生的营养物质发生转移,叶片脱落或枯萎有关[19]。单作条件下果实的脂肪、蛋白质、棕榈酸、硬脂酸、油酸、亚油酸、亚麻酸、氨基酸的含量要极显著或显著高于间作条件下果实内的含量。而间作模式下的果实单宁含量则极显著高于单作种植模式,通常单宁含量越多,果仁口感越涩[20]。孙桂丽等[21]在研究光照对库尔勒香梨品质的影响时表明,光照的不同就会带来果实的品质的差异;周先艳等[22]同样认为,由于树体本身受到的光照差异,相对的使水晶蜜柚的含糖量出现了变化。研究中,核桃的单作条件下的LAI值显著高于间作条件,加之核桃间作物冬小麦的存在,减少了一定程度光的反射[23],相应的降低了核桃树体对光的利用率,影响了碳物质的积累,从而影响了核桃果实品质的变化。果实的品质与光照有着紧密的关系[24]。
林粮间作会带来一定程度的遮阴,张宏芝等[25]进行遮阴程度对小麦光合特性的结果中指出,无遮阴条件的冬小麦在扬花期及灌浆期的净光合速率应在30 μmol/(m2·s)以及25 μmol/(m2·s),而研究中测定的间作条件下的小麦的扬花期与灌浆期净光合速率仅为14和12 μmol/(m2·s),林粮间作会降低小麦的光合能力。植物叶片的光饱和点与光补偿点反映了植物对光照强度条件的要求[26-27]。张宏芝[28]等在研究核麦间作下冬小麦的冠层光分布时表示仅冬小麦在冠上部的PAR值大于冬小麦的光补偿点,冠层下部的PAR只在很短的时间中略大于光补偿点。研究中,运用Lambert-Beer定律进行叶面积指数与光合有效辐射值计算后,到达冬小麦的光合有效辐射值仅为230 μmol/(m2·s),虽然值高于光补偿点,却远低于光饱和点,冬小麦在与核桃间作时,由于光照不足,限制了光合作用的快速进行进而影响有机物的积累。王世伟[29]在核桃与粮棉间作系统光环境特征中指出,间作条件下的冬小麦对比单作冬小麦蒸腾速率明显降低,并且随着PAR的降低,间作冬小麦的光合速率和气孔导度也在降低,但是胞间CO2浓度却有所增高,是间作导致光能供应不足,使光合作用受到限制。研究中,不同时期的冬小麦Pn、Gs、Tr均随光合有效辐射的增加而增加,胞间CO2浓度则随着光合有效辐射增加而减小,并未得到与王世伟等[29]研究相同的结论,可能是由于间作的遮阴使小麦旗叶光合速率与蒸腾速率降低,以及气孔导度的失控,使得叶片内外CO2不能正常平衡交换,导致细胞间隙CO2过量积累,进一步影响光合速率及其相关参数[30]。在种植核桃时,应尽量避免间作,如必须间作则需要增加核桃的株行距。
4 结 论
4.1间作园内核桃的叶面积指数小于单作园内的核桃叶面积指数,并且间作园果实品质低于单作园果实品质,由于光环境的改变使间作园核桃不能完全进行光合作用,对光合同化产物的产生造成一定影响,进而影响了果实品质。应改善核桃间作园内的光环境,以避免因光环境差异,而带来的影响。
4.2间作小麦的扬花期及灌浆期的净光合速率值为14和12 μmol/(m2·s),单作小麦同时期净光合速率为30和25 μmol/(m2·s),间作带来的遮阴的确影响了小麦的光合作用,增大主作物核桃的株行距或者在6 a生以上树龄的核桃园不再进行间作。
