不同大小入侵小飞蓬资源分配特性
2022-04-01张宝成孙晴黄小桐曾钢白艳芬
张宝成 孙晴 黄小桐 曾钢 白艳芬
摘要:小飞蓬是一种常见入侵植物,它会采用不同的策略适应入侵地新环境。采用测量、称质量等方法研究不同大小个体小飞蓬的不同器官分配模式,以期探讨小飞蓬的入侵机制。结果表明,3个不同质量阶段的小飞蓬采取了不同的分配策略。整株质量为0~10 g的小飞蓬,其根质量与根幅和根长呈极显著相关关系,根质量增加1 g,其根幅和根长分别增加为4.28、4.25 cm;在整株质量>15 g的小飞蓬,根质量增加1 g,其根幅和根长仅分别增加0.39、0.41 cm。整株质量在>10~15 g的小飞蓬的根质量与根长和根幅无明显关系。整株质量为0~10 g的小飞蓬,其整株质量增加1 g,其根质量和叶质量增加分别为0.15、017 g。整株质量>10~15 g的小飞蓬,根质量投入和叶质量随着整株质量增加1 g,它们的增加速度分别为0.22、0.21 g;整株质量>15 g的小飞蓬,其增加速度分别为0.24、0.12 g。0~10 g和>15 g的小飞蓬,花的质量随着株高增加而增加,但是>10~15 g的小飞蓬,花质量随株高的增加而降低。>15 g的小飞蓬,整株质量增加1 g,繁殖器官花的投入增加0.08 g;而整株质量为0~10 g的小飞蓬繁殖器官花的投入却增加0.39 g。这表明,整株质量越小的小飞蓬其有性繁殖的投入越大,这可能是小飞蓬入侵成功的关键。
关键词:小飞蓬;构件;繁殖器官;生物入侵;相关关系;分配策略
中图分类号: S451文献标志码: A
文章编号:1002-1302(2022)05-0107-06
收稿日期:2021-05-08
基金:贵州省科技厅合作项目[编号:黔科合LH字(2017)7063];贵州省科技厅项目[编号:黔科合支撑(2018)2364];遵义师范学院博士基金[编号:遵师BS(2014)07号]。
作者简介:张宝成(1978—),男,陕西南郑人,博士,副教授,从事生态功能及资源生态研究,E-mail:bczhang@fudan.edu.cn;共同第一作者:孙 晴(1999—),女,江苏连云港人,从事生态学研究,E-mail:1808805040@qq.com。
通信作者:白艳芬,博士,讲师,从事生态学研究。E-mail:823523593@qq.com。
入侵植物随着贸易和人们旅游及全球气候变化而扩散,进而入侵新领地[1]。物种入侵每年给我国造成的经济损失高达1 198.76 亿元[2],对当地生态功能和结构带来严重影响。如互花米草入侵影响食物链降低了当地生态系统的服务功能[3]。研究发现,入侵植物通过化感物质抑制植物的萌发和生长以增加竞争力[4-6],入侵植物分泌的化感物质可改变土壤的化学性质,影响到土壤动物的数量,尤其是弹尾目动物[7-8]。土壤动物影响地表凋落物的分解和营养物质的矿化供给可利用状态。
由于植物器官的可塑性,入侵植物在与土著植物竞争时,通过自身的可塑性改变器官分配。枝、叶、花、根是植物重要器官,都是植物光合作用产物的重要组成部分,它们对植物维持生长、繁殖和资源获取和防御等众多功能始终存在如何优化分配,即资源分配的权限使其功能适应不同的环境[9-10]。因此,植物资源分配在一定程度上反映了其生活史的特点[11]。植物资源分配有繁殖分配,即资源分配到繁殖体中的营养比例[12]。植物在生长中分配到茎、 叶、根、花等器官的生物构件,反映的是植物对环境的适应。目前关于这些器官分配模式在小飞蓬上的研究较少。外来植物能否成功入侵不仅取决于其本身的生物学特性,还与环境的可入侵性密不可分,掌握其生活史对策及生态幅范围对其入侵機制的研究至关重要。
小飞蓬原产于北美洲,现已入侵我国各地,它能够破坏入侵地的生物多样性,也是害虫的中间宿主,有利于害虫的繁衍。它还通过分泌化感物的方式来抑制邻近其他植物的生长发育。近年来不少学者将植物器官组成引用到入侵生态学中,利用种群构件理论研究外来入侵植物根、茎、叶、花等各器官的数量特征及其相互关系,通过对入侵植物表型的可塑性研究,探索小飞蓬的入侵能力、入侵机理、对外界环境的适应性[6]、表型可塑性[8]。本研究对小飞蓬的株高、整株质量、根质量、茎秆质量、花质量、叶质量和根幅质量等进行测定,分析小飞蓬的适应策略,以期为研究小飞蓬的表型可塑性提供科学积累,为小飞蓬的利用和防治提供有效依据。
1 材料与方法
1.1 试验材料
以遵义师范学院校园池塘小山下的小飞蓬为研究对象。遵义市新蒲新区为亚热带季风气候,年均降水量为1 000 mm。在遵义师范学院小湖旁小飞蓬生长分布的典型区域。
1.2 试验方法
用卷尺测量小飞蓬的株高,将植株整株用小铲慢慢地全部挖出根系同时测定最大根幅(小飞蓬水平根系分布最大直径)及根系垂直最深深度(根长)[13],分别把不同器官(花、叶)分袋,洗去根部泥土后吸去水进行编号,放在干燥烘箱60℃烘至恒质量后,称其质量。
1.3 数据处理
用Excel、SPSS 11.5、Sigmplot 10.0进行数据处理并作图。
2 结果与分析
2.1 小飞蓬数量特征
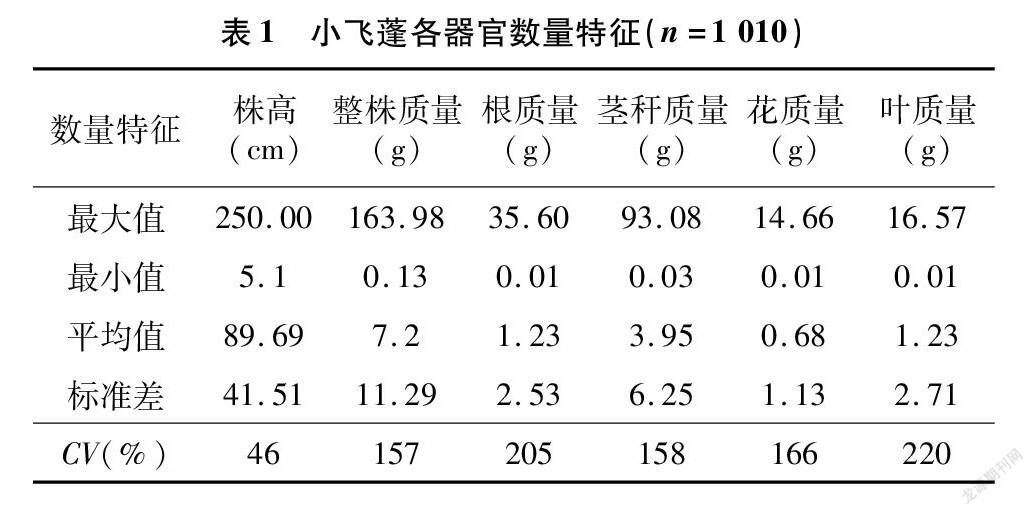
由表1可知,小飞蓬整体样本株高的变异系数最小,为46%,最大值为 250.00 cm,最小值为 5.10 cm。整株质量变异系数为157%,最大值为163.98 g,最小株仅为0.13 g,平均值为7.20 g。根质量变异系数为205%,最大值为35.60 g,最小为0.01 g,均值为1.23 g。茎秆质量、花质量、叶质量变生物量是生物从环境中所获取资源的综合体现,按照频次图研究的1 010个样本的整株质量可分成4个段:0~10、>10~15、>15~30、>30 g。由于>30 g的样本数小于30个,因此对其合并划分为0~10、>10~15、>15 g等3个阶段,并对其数据特征进行处理。
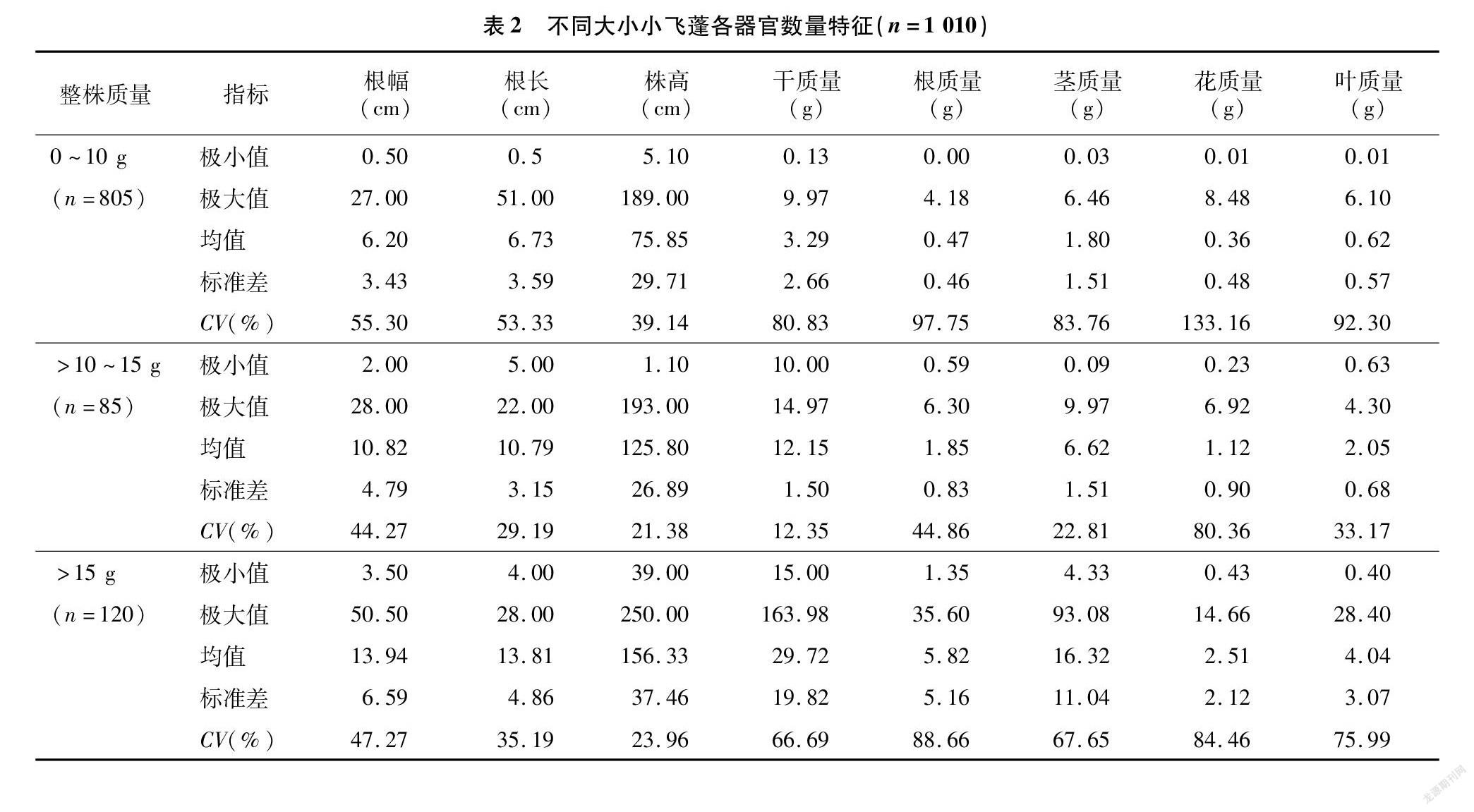
由表2可知,根幅和根长在>10~15 g变异系数最小,其次是>15 g阶段,0~10 g的变异最大,为55.30%。株高的变异系数最大的为0~10 g阶段,>15 g与>10~15 g阶段的变异系数接近,分别为23.96%、21.38%。干质量的变异系数也呈现出与株高类似的变化规律,整株0~10 g阶段的小飞蓬变异系数最大,达80.83%;其次是>15 g阶段小飞蓬的变异系数,为66.69%;最小的为整株质量>10~15 g 阶段的小飞蓬,仅为12.35%。
与整株干质量相比,根质量的变异系数较大,茎质量变异系数也是0~10 g阶段的最大,其次是整株质量>15 g,>10~15 g阶段的最小,依次为83.76%、67.65%、22.81%。0~10 g阶段的小飞蓬繁殖器官花质量的变异系数最大,达133.16%,>10~15 g阶段与>15 g阶段小飞蓬的变异系数较接近,分别为80.36%、84.46%。与繁殖器官花相比,叶质量变异系数较小,0~10 g阶段小飞蓬的变异系数为92.30%;其次是>15 g阶段,为75.99%,>10~15 g 阶段的变异系数为33.17%。
2.2 小飞蓬根指标与根质量的关系
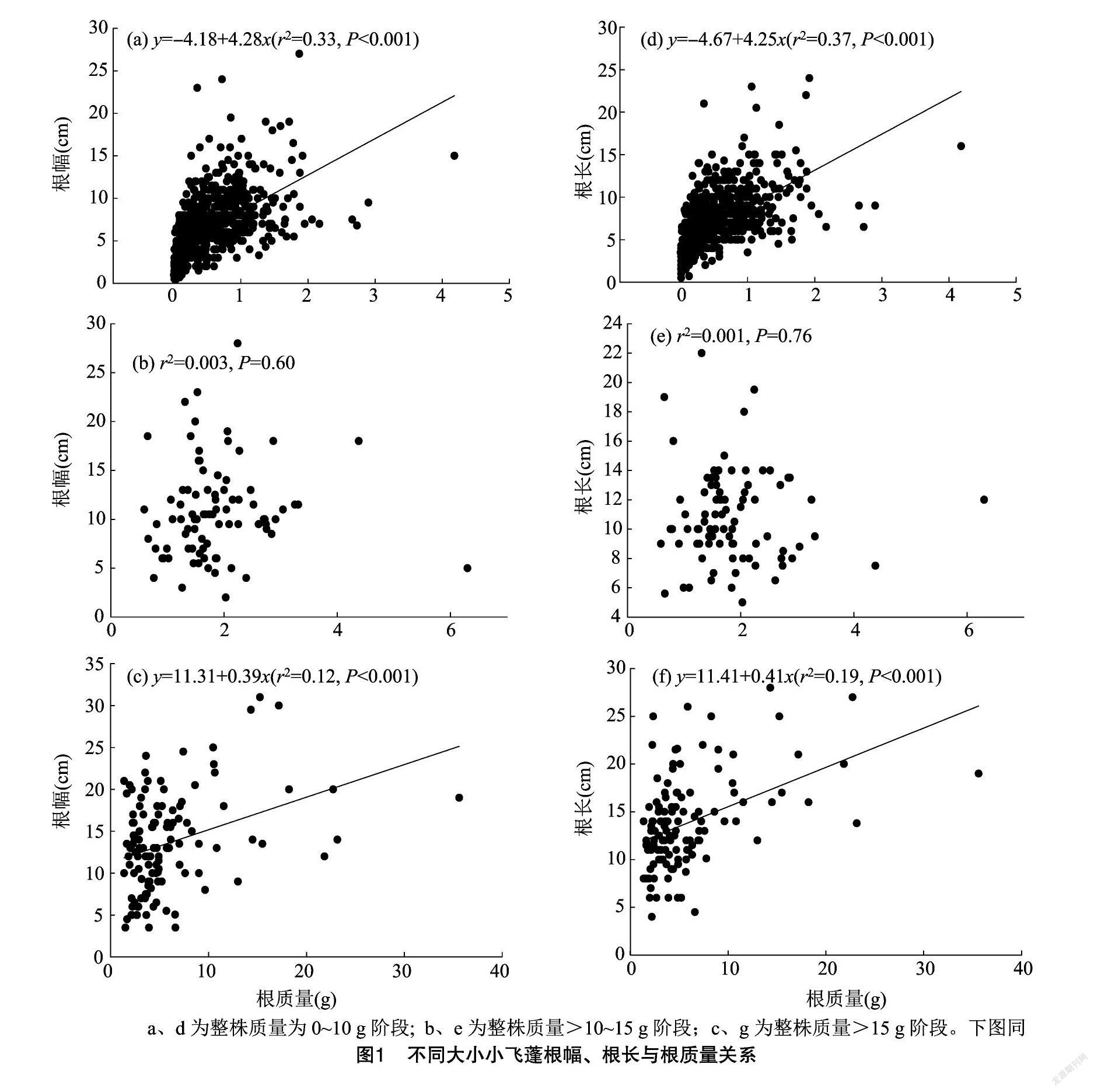
由图1可知,3种不同阶段的小飞蓬表现了不同的投资策略模式。在个体低矮,质量较小的小飞蓬根部营养主要投向根幅和根长,根的质量与根幅呈现出线性相关。0~10 g阶段的小飞蓬根质量增加1 g根幅增加4.28 cm(P<0.001),根的质量能够解释该方程的33%(r2);根质量增加1 g根长度增加4.25 cm(P<0.001),根的质量能够解释该方程的37%。质量>15 g的小飞蓬,根质量增加投入在根幅和根长的增加速度分别为0.39、0.41 cm (P<0.001),根质量分别能够解释该方程的12%、19%。整株质量在>10~15 g阶段的小飞蓬, 其根质量与根幅和根长没有显著相关性,P值分别为060、0.76。
2.3 小飞蓬根、叶质量与整株质量的关系
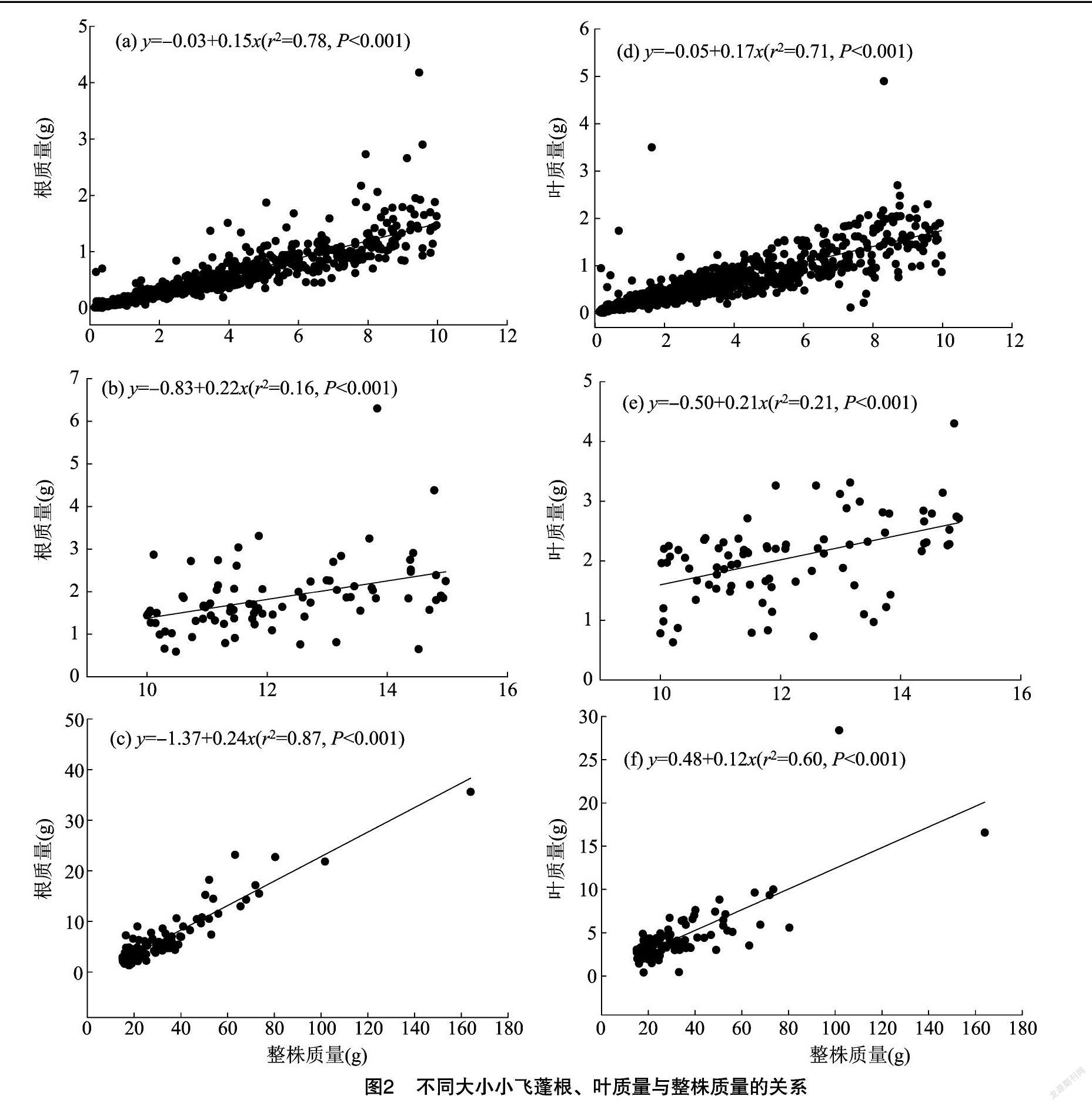
叶和根是植物生长的重要器官。由图2可知,3种不同大小阶段的小飞蓬表现出不同的投资策略模式。在整株质量较小的小飞蓬(0~10 g阶段),整株质量每增加 1 g,根系质量增加0.15 g,最小;其次是>10~15 g阶段, 整株质量每增加1 g, 根系质量增加0.22 g;整株质量>15 g的小飞蓬,整株质量每增加1 g,根系质量增加0.24 g。
整株质量>15 g的小飞蓬,整株质量每增加 1 g,叶片质量增加0.12 g;其次,整株质量较小的小飞蓬(0~10 g阶段),整株质量每增加1 g,叶片质量增加0.17 g;>10~15 g阶段,整株质量每增加 1 g,叶片质量增加0.21 g。
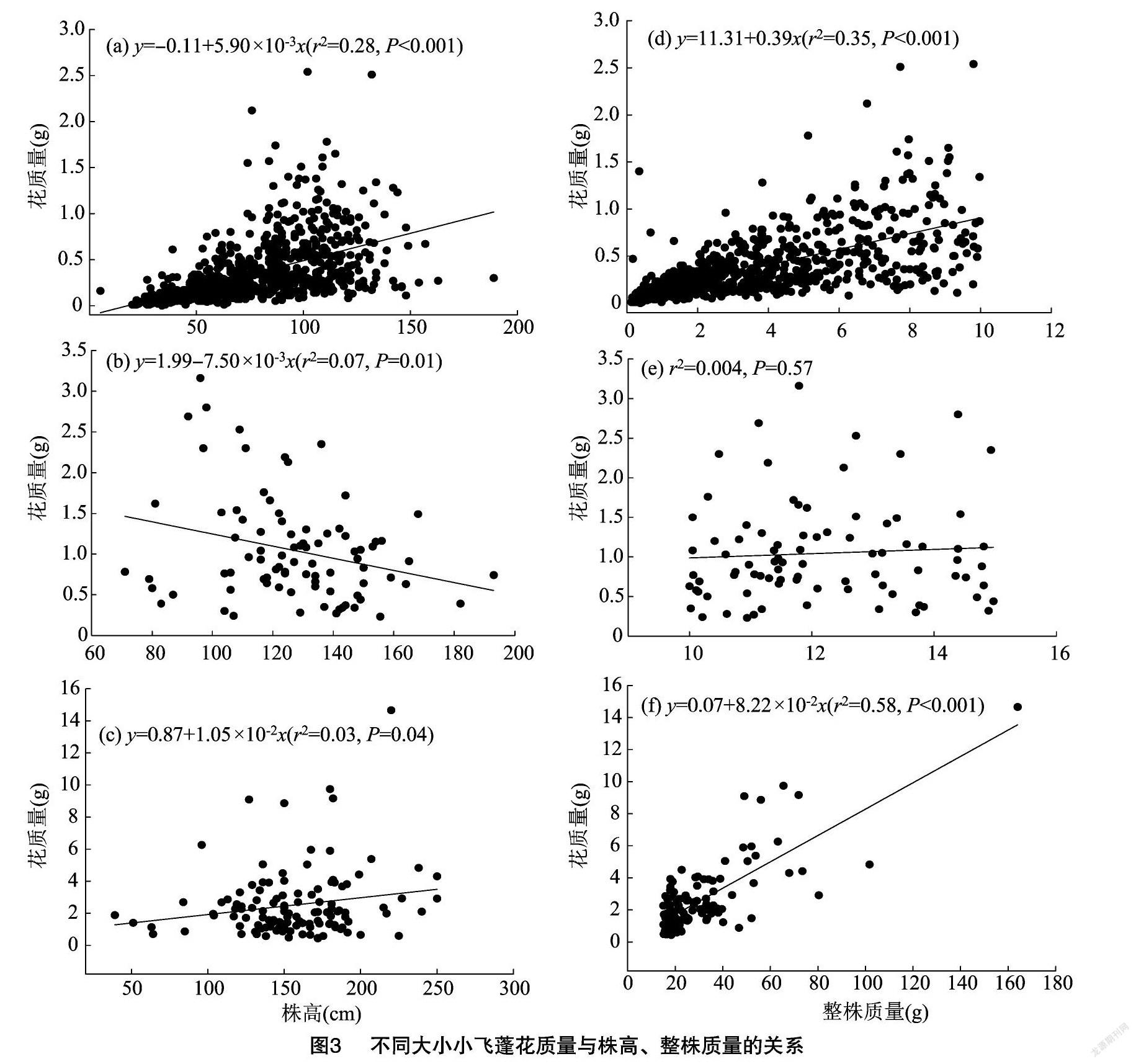
2.4 小飞蓬花质量与株高、整株质量的关系
由图3可知,小飞蓬的株高与其繁殖器官花的质量呈显著相关性(P<0.05)。整株质量>15 g的小飞蓬,株高增加1 cm,其繁殖器官花质量增加1050 mg。处于中间阶段,整株质量>10~15 g的小飞蓬随着株高增加,繁殖器官花的质量呈现降低趋势,高度增加1 cm,其花质量减少750 mg。对于个体小整株质量在0~10 g阶段的小飞蓬,高度增加1 cm,其花质量增加 590 mg。
如果从小飞蓬整株质量角度考虑,处于中间阶段整株质量在>10~15 g范围内的小飞蓬,其整株质量与繁殖器官花的质量无显著关系(P=0.57)。处于质量较大个体的小飞蓬(整株质量>15 g),质量增加1 g,繁殖投入增加82.20 mg;而整株质量为0~10 g的小飞蓬,质量增加1 g,繁殖投入增加390 mg。
3 讨论
由于植物的器官根、茎、叶、秆和花在植物的生长中扮演着不同的角色。叶片是植物光合作用重要的器官,植物吸收CO2合成有机物分配于根、茎、叶、秆等组织以便与周围其他植物竞争中处于优势竞争地位。
植物的生物量投资于根系的比值为对根的投入比,如整株质量>15 g的小飞蓬投入比为0.24 g/g,整株质量为0~10 g的小飞蓬投入比为0.15 g/g。根据方程的决定系数(r2)对模拟的曲线方程进行解释,仅仅质量>15 g的小飞蓬和0~10 g小飞蓬分别能够解释87%、78%。而質量在10~15 g的小飞蓬解释率仅为16%。可能是整株质量>15 g的小飞蓬,竞争压力大,根系投入比值大,拟合解释率高,株较大,需要吸收较多的营养来促进生长适应环境[14-15]。根据解释率高的拟合曲线,小飞蓬整株生物量投入效果最高的是>15 g阶段的小飞蓬,投入叶比为0.24 g/g,比0~10 g的小飞蓬高36%。这可能是株质量小的飞蓬高度与周围的植物竞争压力较小利用了环境中的生态位空缺,高大的小飞蓬(>15 g)高度高,需要与周围的灌木进行竞争,因此投入更多的生物量以在竞争中获得优势[15]。
根系在土壤中的分布情况会影响植物在其生长的土壤中获取营养和水,且起着支撑作用。根系沿着土壤水平生长,可以占据较多的肥沃土,对其长势、地下竞争和群落结构具有影响,根幅分布影响到土壤资源的占有和周围根系的错位复杂程度[16]。主根长度影响到植物须根数量和株高[17],主根长度主要担负着支撑和萌发更多的次生根作用。0~10 g阶段小飞蓬的根系质量与根幅和主根长度的拟合曲线决定系数分别为33%、37%,远高于>15 g的小飞蓬的12%、19%。而整株质量 >10~15 g 的小飞蓬根质量与根幅和根长无明显关系(P≥0.60),从另外一方面反映出更多的营养投入到了根系和叶等器官中。
植物的花是植物的重要繁殖器官,根据株高与花质量的关系,个体质量小的小飞蓬呈现显著的相关关系,拟合方程的解释率也高,系数贡献较大。整株生物量与花质量关系极显著相关的是整株小和整株大的小飞蓬阶段,尤其是0~10 g的小飞蓬是>15 g小飞蓬投入繁殖系数的4.74倍。意味着较小个体的小飞蓬有较多的繁殖投入。高寒草甸系统研究表明,个体越大越高投入的繁殖量约大[10]。本研究与其不一致,但是本研究支持刘左军等的研究结果[11]。适当的压力胁迫可以促进植物的繁殖投入[18]。郭水良关于北美车前地研究也表明,在逆境压力下增加繁殖投入,北美车前在逆境环境中,增加了其种子投入[19]。利用大量子代数量优势增加子代在环境中的数量提高繁殖[19]。小飞蓬单株产种子量为4×105~1×106粒,种子的质量为2×105~3×105 g/粒,如果按照这个比例[20],小飞蓬在整株质量为0~10 g阶段,按照花器官投入平均值计算,整株质量增加1 g,繁殖的种子投入增加1.08倍,这意味着整株小飞蓬会增加4.32×105~1.08×106粒种子。在适宜的环境下90%的种子均可萌发,可见小飞蓬是典型的r繁殖策略。
在小飞蓬的繁殖器官花、叶、根质量以及根长等器官与植物其他相关器官的拟合方程中,r2为拟合方程的决定系数。r2可用来估算生物遗传的贡献,用1-r2估计环境因子的作用[21]。在根幅与根质量的关系中,决定系数最小的为0.003,最大的为0.330;根长与根质量拟合中也出现决定系数类似的情形,在0.001~0.370之间波动,也就是说环境因子的作用在0.630~0.999之间波动,可见入侵植物根系的水平分布和垂直分布受环境的作用较大,也就是说其在根器官的分配获取土壤资源的可塑性很大。
整株质量与根质量的决定系数在0~10 g和 >15 g 这2个阶段的小飞蓬的遗传生物因子贡献率分别为0.78、0.87,而与小飞蓬叶器官分配决定系数分别为0.71、0.60;在>10~15 g的小飞蓬的决定系数为0.16,可见在不同大小其可分配的决定系数不同。在小飞蓬繁殖器官花的质量与整株质量关系中,遗传因子的作用较小,在个体比较小的和比较大的阶段,遗传因子贡献率分别为0.35、0.58。但是结合单位质量的增加与繁殖器官的投入,个体较小阶段的投入很大。可能是环境竞争压力较大,因此小飞蓬投入较大的比例在繁殖器官。可能是生物固有的生存策略,尤其是环境压力较大,其r繁殖策略更加凸显。
4 结论
(1)整株质量为0~10 g的小飞蓬,其根质量与根幅、根长呈显著关系,根质量增加1 g,其根幅和根长分别增加为4.28、4.25 cm;整株质量>15 g的小飞蓬,根质量增加1 g,其根幅和根长仅分别增加039、0.41 cm。整株质量为>10~15 g的小飞蓬的根质量与根长、根幅无明显关系。
(2)整株质量为0~10 g的小飞蓬,其整株质量增加1 g,根质量和叶质量增加分别为0.15、0.17 g。整株质量为>10~15 g的小飞蓬,根质量投入和叶质量随着整株质量增加1 g,它们的增加速度分别为0.22、0.21 g;整株质量>15 g的小飞蓬,其增加速度分别为0.24、0.12 g。
(3)0~10 g和>15 g的小飞蓬,花的质量随着高度增加而增加,但是>10~15 g的小飞蓬,花质量随株高的增加而降低。>15 g的小飞蓬,整株质量增加1 g,繁殖器官花的投入增加0.08 g;而整株质量为0~10 g的小飞蓬繁殖器官花的投入却增加039 g。这表明,整株质量越小的小飞蓬其有性繁殖的投入越大,这可能是小飞蓬入侵成功的关键。
另外,整株质量小(0~10 g)的小飞蓬产生种子的能力较强,因此需要加强控制。
参考文献:
[1]于 飞,吴海榮,吴福中,等. 潼湖生态智慧区外来入侵杂草调查研究[J]. 杂草学报,2020,38(4):20-25.
[2]王睿芳,冯玉龙. 叶物候、构建消耗和偿还时间对入侵植物碳积累的影响[J]. 生态学报,2009,29(5):2568-2577.
[3]Li B,Liao C Z,Zhang X D,et al. Spartina alterniflora invasions in the Yangtze River estuary,China:an overview of current status and ecosystem effects[J]. Ecological Engineering,2009,35(4):511-520.
[4]张宝成,陈通丽,李德辉,等. 喜旱莲子草浸提液对萝卜种子萌发和幼苗生长的影响[J]. 种子,2018,37(1):84-87.
[5]王有兵,杨建军,李子光,等. 我国自然保护区入侵植物种类概况[J]. 杂草学报,2020,38(3):1-6.
[6]张宝成,王 平,金 星. 观赏植物与生物入侵[J]. 北方园艺,2018(5):178-183.
[7]范建军,乙杨敏,朱珣之. 入侵杂草一年蓬研究进展[J]. 杂草学报,2020,38(2):1-8.
[8]陈 雯,李 涛,郑荣泉,等. 加拿大一枝黄花入侵对土壤动物群落结构的影响[J]. 生态学报,2012,32(22):7072-7081.
[9]Cheplick G P. Life history trade-offs in Amphibromus scabrivalvis (Poaceae):allocationtoclonalgrowth,storage,andcleistogamous
reproduction[J]. American Journal of Botany,1995,82(5):621-629.
[10]梁 艳,张小翠,陈学林. 多年生龙胆属植物个体大小与花期资源分配研究[J]. 西北植物学报,2008,28(12):2400-2407.
[11]刘左军,杜国祯,陈家宽. 不同生境下黄帚橐吾(Ligularia virgaurea)个体大小依赖的繁殖分配[J]. 植物生态学报,2002,26(1):44-50.
[12]李 博,张大勇,王德华. 生态学——从个体到生态系统[M]. 4版.北京:高等教育出版社,2016.
[13]张玉芹,杨恒山,高聚林,等. 施钾方式对高产春玉米根系分布及其活力的影响[J]. 水土保持通报,2015,35(6):64-69.
[14]Wilson J B. Shoot competition and root competition[J]. The Journal of Applied Ecology,1988,25(1):279.
[15]王 平,王天慧,周道玮,等. 植物地上竞争与地下竞争研究进展[J]. 生态学报,2007,27(8):3489-3499.
[16]陈 伟,薛 立. 根系间的相互作用——竞争与互利[J]. 生态学报,2004,24(6):1243-1251.
[17]章 敏,金方伦. 柑桔苗木主根长度与其地上部分特征变化的相关性[J]. 天津农业科学,2017,23(5):89-93.
[18]古 琛,赵天启,王亚婷,等. 短花针茅生长和繁殖策略对载畜率的响应[J]. 生态环境学报,2017,26(1):36-42.
[19]郭水良,方 芳,黄 华,等. 外来入侵植物北美车前繁殖及光合生理生态学研究[J]. 植物生态学报,2004,28(6):787-793.
[20]张 帅. 外来植物小飞蓬入侵生物学研究[D]. 上海:上海师范大学,2010.
[21]杨允菲,李建东. 松嫩平原全叶马兰种群不同龄级分株的生长策略[J]. 应用生态学报,2003,14(12):2171-2175.
