木莲(Manglietia fordiana)的适生分布、影响因子与保护空缺研究
2022-03-30朱祥福陈伏生曾菊平
邹 武,王 渌,赖 童,朱祥福,陈伏生,曾菊平,*
1 鄱阳湖流域森林生态系统保护与修复国家林业和草原局重点实验室,江西农业大学林学院, 南昌 330045 2 江西九连山森林生态系统定位观测研究站, 龙南 332900 3 江西九连山国家级自然保护区管理局, 龙南 332700
生境保护是维持生物多样性的基本途径[1],而自然保护区是生境保护最有效方式之一[2—3]。当前,我国生物多样性保护体系仍不完善,一些重要生态系统和生物资源尚未得到有效保护[4—5],生物多样性下降的总体趋势仍待有效遏制[6]。而优先区域是生物多样性保护重点和关键区域[7],当前急需开展优先区域内现有自然保护区的保护效果评估,同时,更需分析优先区域保护空缺[8],优化自然保护区空间布局[9]。
山地阔叶林是我国南方典型森林生态系统,维持着横断山脉、武陵山脉、南岭山脉、华东山区(如武夷山脉)、海南岛(如五指山脉)等生物多样性热点[10],是山地生物多样性保护优先区域。然而,受长期人类活动影响及其自身山地分布特征(如孤立、隔离等),山地阔叶林生境已急剧减少,呈现明显破碎化,使得自然保护区也呈现片段化分布[4]。因而,急需在保护空缺分析基础上,扩增、新增自然保护区[6],并通过小保护区、生物廊道、跳脚石等建设,完善自然保护区规划、设计,优化山地阔叶林保护网络,提高整体保护水平。
木莲Manglietiafordiana是典型的山地阔叶林树种,主要在我国武夷山脉、南岭等南方中山山脉发生[11],研究木莲发生、分布特征与保护需求,可一定程度反应我国南方山地阔叶林保护状况,提出优化方向。木莲又称黄心树,树干通直圆满、木材结构细、文理直,且耐腐性好、易加工,是建筑、家具和胶合板优良用材[12—14],野生种群易因过度利用、生境丧失等受到威胁,地理种群下降甚至消失。这种消失不仅代表生物多样性(如遗传多样性)的丧失,也同时降低了该地区的科学、生态和社会价值[1]。基于系统保护规划(如空缺分析,GAP analysis)[15]思想来保护生物多样性,成为当前测定保护地有效性的重要方法[16—17],通过将生物多样性优先区与现有保护区进行比较分析[18],找出生物多样性保护的空缺,并在空缺地带建立保护区[1]。而关注地理种群的保护空缺,同样有利于提高物种种下多样性保护的有效性。
本研究根据木莲典型的山地、阔叶林分布特征[19],基于生态位理论[19—20],在适生分布预测基础上,叠加阔叶林生境要素,并考虑海拔偏好,获得木莲的保护优先区,进行空缺分析。同时,基于地理山脉的隔离作用,选择、判定木莲各地理种群的保护空缺,为优化山地阔叶林保护网络提供参考。本次致力于(1)木莲的全球分布与适生范围;(2)木莲保护优先区的确定;(3)木莲地理种群的空缺分析;(4)木莲九连山种群案例分析,结合大尺度适生分布与空缺分析、地方适宜生境与功能区保护,探讨木莲保护现状与空缺,提出优化方向,为山地阔叶林优先区保护提供参考。参考植物智(http://www.iplant.cn)将乳源木莲Manglietiayuyuanensis归为木莲的异名等分类方法[13—14],且由于两者形态特征重叠、野外识别困难,本次不区分乳源木莲与木莲,而均记录为木莲。
1 材料与方法
1.1 研究区域与方法
1.1.1研究区域
木兰科间断分布于亚洲与美洲热带至温带地区,而木莲只分布在亚洲,故本次选择木兰科的亚洲分布区作为木莲适生分布研究区域。通过查询有关木兰科地理起源、分布文献或著作[21—23],获得木兰科在亚洲的大致分布后,将其与全球生态地理区划[24]叠加,并选定有重叠的生态区作为本次木莲适生分布研究区域。从图1可知,该区域覆盖东亚区、喜马拉雅区、印度区、印度支那区、马来西亚区及部分热带地区等。
1.1.2地理发生点
为获得木莲地理发生点,用于预测其在研究区的适生范围,从中国植物数字标本馆(CVH)、全球生物多样性信息网络(GBIF)采集有关木莲或乳源木莲的具有经纬度或地名信息发生点,去除部分信息不确定或地名模糊数据后,再据地名信息在Google地球或百度地图查询补充经纬度信息,最后获得有效木莲发生点296个,将其保存为CSV文件,以备后用。
1.1.3木莲适生分布预测
从WorldClim(http://www.worldclim.org)采集研究区范围的19个生物气候变量(Bio1至Bio19,1950—2000年平均值)[25—26]栅格,分辨率为30弧秒(近似等于1 km×1 km)。将分辨率一致的DEM数据导入ArcGIS,用栅格计算生成研究区坡向、坡度、海拔高度3个地形栅格。将以上22个环境栅格保存为ASC格式,以备后用。
用R语言Hmisc数据包计算22个环境变量的Spearman相关系数,对相关系数> 0.7的变量进行筛选后,选定对木莲分布更具生物学意义的bio14(最干月降水量)、Ele(海拔高度)、bio4(温度季节性变化标准差)、bio18(最暖季降水量)、bio6(最冷月最低温度)、bio12(年降水量)、bio16(最湿季度降水量)、bio15(降水量季节性变化)、bio2(昼夜温差月均值)作为适生分布预测变量。
适生分布建模与预测用当前被广泛用于物种潜在分布的MaxEnt[27]方法完成。参考朱耿平和乔慧捷等[19]有关MaxEnt参数优化方法,用ENMeval数据包调整模型调控倍频(regularization multiplier, RM)和特征组合(feature combination, FC),选择AIC(Akaike information criterion correction)[28]值最小时的RM参数(1.5)、特征组合(1.5)与LQHPT模型预测木莲适生分布。在MaxEnt界面,导入以上选定的环境变量(ASC格式)与木莲发生点(CSV格式),选定25%作为测试集(test data)、75%作为训练集(training data),设置重复运行100次。用AUC(Area under the curve)评估各模型的精确性[29],其值> 0.8表示达到精确要求[30],而值越大预测越精确[31],因而将AUC值最大的模型作为最优模型,并用刀切法(Jackknife)评估模型中各变量对预测适生分布增益的相对贡献(表1)。
将最佳模型预测分布以avg.asc格式输出,导入Arcgis,按概率值(P值)重分类,将0≤P<0.2定义为非适生、0.2≤P<0.4为低适生、0.4≤P<0.6为中适生、0.6≤P≤ 1为高适生,获得木莲适生分布(Potential Distribution, D)后,计算各适生等级栅格数与比重。以上过程在R语言与Arcgis10.2(http://www.arcgis.com)完成。
1.2 案例研究:九连山种群
为了解木莲在种群水平的发生、分布与生境需求特点,选择木莲九连山种群为研究案例,在九连山保护区范围内开展野外种群调查。九连山跨广东、江西两省,在江西省龙南县一侧残存着一小片原生性常绿阔叶林,成为当地生物庇护所。自1957年,地方性保护政策开始在该片原生林实施,并于2003年围绕该原生林设立“江西九连山国家级自然保护区”(24°29′18″—24°38′55″N,114°22′50″—114°31′32″,图3)[32],进一步加强保护力度。九连山在区划上处于中亚热带湿润常绿阔叶林与南亚热带季风常绿阔叶林过渡带[32],地带植被属典型常绿阔叶林。本次调查覆盖大部分保护区范围,并覆盖原生林及其周边次生林、人工林等多种森林生境。
1.2.1木莲植株分布调查
在2018—2019年间,采用样线法沿道路、林缘、山脊、林间小道等行进,调查沿线木莲(包括乳源木莲)植株,用GPS仪记录遇到植株(含苗株)的经纬度、海拔、生境等信息。行进中,先借助肉眼或望远镜(10×42)查看沿线植物,根据树形、树干、顶芽、花、果实等初步判定为木兰科植株后,再在树下捡拾或采集少量枝叶、果实、花等,参考《中国木兰》[23]与中国在线植物志http://www.iplant.cn确定为木莲或乳源木莲植株后做好记录。
本次调查样线长度共计134km(含部分非林区开阔地的行车路线),覆盖保护区海拔范围(300m至1400m),并贯穿当地常绿阔叶林、常绿针叶林、常绿灌丛、常绿针阔混交林、竹林等林区,以及农田、荒地等非林区,共记录木莲534株(包括苗株),以此统计分析九连山木莲种群发生对海拔、生境的需求特征,为后续有关木莲生境、海拔的优先保护提供参考。
1.2.2九连山种群的适宜生境
在九连山保护区范围内,用30弧秒(近似于1 km × 1 km)栅格作为取样单位,每个栅格选取一个位点,从以上调查获得的534个木莲植株位点中稀释、筛选出17个点(以降低取样偏差,避免过度拟合[19]),作为当地木莲种群适宜生境分布预测的发生点。同样地,将含有经纬度信息的17个发生点保存为CSV文件,连同九连山保护区的22个环境变量栅格(ASC格式),一起导入MaxEnt界面,构建最佳模型后,预测获得木莲在九连山保护区的适宜生境分布。
1.2.3核密度分析
将以上534个木莲植株位点导入Arcgis,采用核密度分析方法,获得核密度分布图,并根据核密度中心、次中心位置,判断木莲九连山种群的发生或分布中心,再与九连山保护区功能区划图叠加,分析当地木莲种群的受保护状况(如是否在保护功能上有缺失或空缺?)。
1.3 木莲的保护与空缺分析
考虑到野外木莲常在阔叶林发生[11],且本次九连山案例也显示出类似选择,以及本次采集的木莲发生点均位于中国,本文基于以下步骤进行木莲的保护空缺分析:(1)从以上木莲的预测适生区,选择中国范围作为木莲的主体适生区(Main-part of Potential Distribution, Dm);(2)将Dm与中国2015年自然保护区(Nature Reserve in China, NRc)分布矢量图叠加,掩膜提取获得木莲的适生区保护分布(Protected Dm, pDm);(3)同时,将Dm与中国阔叶林分布图叠加,掩膜提取获得木莲的阔叶林适生区分布(Broad-leaved forest Dm, BDm);(4)将BDm与中国自然保护区分布叠加,掩膜提取获得木莲的阔叶林适生区保护分布(Protected BDm, pBDm);(5)分别计算Dm、pDm、BDm与pBDm的高、中、低适生等级的栅格数及其所占比重;(6)基于以上4个图层数据进行空缺比重(Gap,G)计算,方法包括:
(1)基于Dm的保护空缺:GDm=1-(pDm栅格数/Dm栅格数)
(2)基于BDm的保护空缺:GBDm= 1-(pBDm栅格数/BDm栅格数)
而 “木莲一般生长在海拔700—1200m的阔叶林…”[11],且从本次九连山案例也获得类似支持依据(如核密度在700—900m最高等)。为此,本文进一步在BDm与pBDm基础上,去除低适生与非适生区后,沿海拔梯度将其分割出<400m、400—700m与 >700m三个海拔高度图层,分析木莲的适生区保护与空缺,而将>700m的阔叶林适生区BDm700作为木莲的优先保护区。不同海拔的空缺比重计算如下:
(3)海拔高度<400m的保护空缺:G400= 1-(pBDm400栅格数/BDm400栅格数)
(4)海拔高度400—700m的保护空缺:G400—700= 1-(pBDm400—700栅格数/BDm400—700栅格数)
(5)海拔高度>700m的保护空缺:G700=1-(pBDm700栅格数/BDm700栅格数)
其中,BDm400、BDm400—700、BDm700分别为不同海拔梯度的阔叶林适生区(不含低适生区),pBDm400、pBDm400—700、pBDm700分别为不同海拔梯度的受保护阔叶林适生区(不含低适生区)。同时,分别基于BDm与BDm700适生分布(不考虑低适生区)进行空缺识别。首先,叠加自然保护区矢量,将图层中的适生区划分成保护区与空缺区两类;其次,据以上296个发生点与中国自然保护区分布的位置关系,将发生点也划分为保护点(保护区内)与空缺点(保护区外)两类;最后,根据空缺或保护点、空缺或保护区分布,参考山脉位置、走向,以及木莲地理种群的保护要求,识别保护空缺。
另外,在九连山案例研究中,本文将自然保护区的功能区划图与获得的木莲九连山种群适宜生境分布图叠加,计算获得核心区、缓冲区与实验区的低、中、高适宜生境栅格数及所占比重,案例分析木莲适宜生境及其等级与各功能区的保护管理强度的匹配性或一致性。
1.4 数据分析
用列联表(Crosstable)与Pearson卡方检验各适生等级分布比重在不同类型适生区(如D、Dm、BDm)的分布差异。用Friedman检验方法比较Dm、BDm各等级适生区的比重。用Wilcoxon Rank Sum Test分析不同植被生境的木莲多度、核密度与适宜度的差异。以上数据分析均在SPSS17.0完成。
2 结果与分析
2.1 木莲的发生范围与适生分布
基于本次采集的296个木莲发生点,可知其主要分布在我国大别山以南至海南岛之间,向东至雁荡山脉,向西达高黎贡山,而集中在武夷山脉、南岭、苗岭等南方中山山地发生,在全球分布上属东亚山地特有种(图1)。

图1 木莲发生点与适生范围(D)及其主体适生区(Dm)与阔叶林提取主体区(BDm)
用MaxEnt获得最佳模型(测试AUC=0.974、训练AUC=0.976),预测木莲适生范围(D),结果显示木莲适生区主要位于亚欧大陆东部沿海山地(图1),包括台湾岛、济州岛、日本群岛南部岛屿,在喜马拉雅南面也存在一个狭长地段(如长山山脉、帕特凯山脉等)。但其主体适生区位于中国南部、西南部中山山地,包括戴云山、南岭、罗霄山、武夷山等,向北延伸至大别山南坡,向西达高黎贡山。而叠加考虑阔叶林分布后,可见其适生区明显呈零散分布,除了在武夷山、南岭等区域集中连成小片外(图1)。
2.2 影响木莲适生分布的生态因子与生态位宽度
表1列出了构建最佳适生模型的生物气候、地形变量,据其贡献度可知,bio14(最干月降水量,贡献度为34.6%)、Ele(海拔高度,19.2%)、bio4(温度季节性变化标准差,15.6%)、bio18(最暖季降水量,12.0%)、与bio6(最冷月最低温度,7.0%)是影响木莲适生分布的关键生态因子,尤其,前4个生态因子的联合解释度达88.4%(表1)。
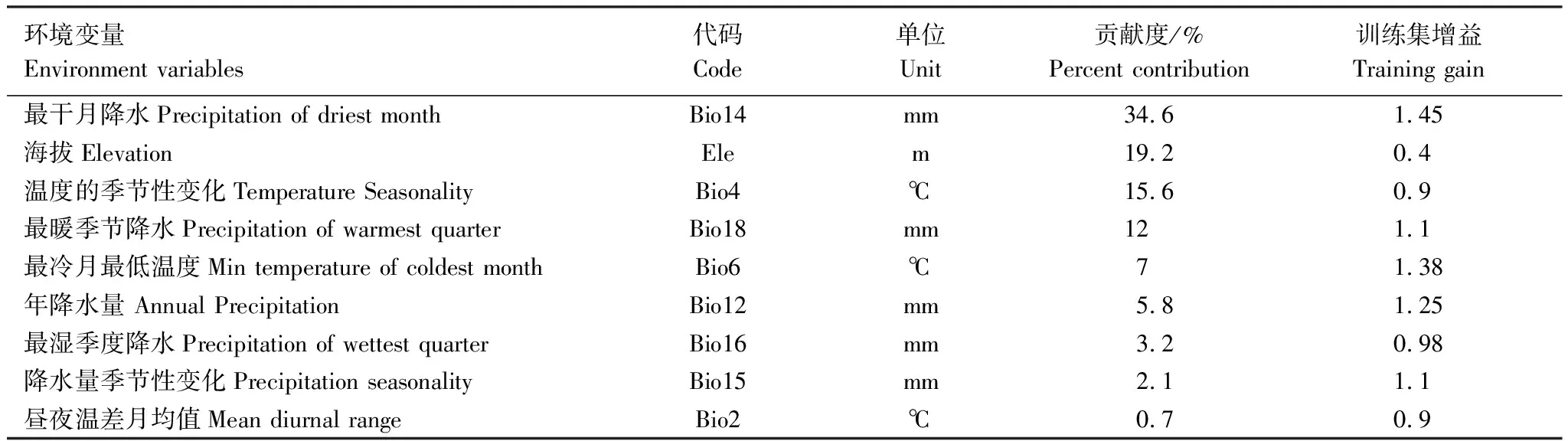
表1 构建木莲适生分布预测模型的环境变量与贡献度
参照≥60%的适合度,计算木莲适生分布生态位宽度,可知木莲的bio14生态位宽度为30—55mm,海拔生态位宽度为400—1500m,bio18生态位宽度为500—800mm(图2)。

图2 影响木莲适生分布的4个关键生态因子(bio14、Ele、bio4、bio18,见表1)响应曲线(绿色短线表示适合度≥60%的生态位宽)
2.3 木莲的山地分布特征:以九连山种群为例
在九连山保护区范围内构建最佳模型,预测木莲的适生分布(平均测试AUC=0.845,平均训练AUC=0.88),分析当地适宜生境。结果显示,九连山种群的中、高适宜生境主要位于保护区东南部(图3),而核密度图也显示多数植株在该地段聚集,形成中心(图3),这与当地海拔高度、植被类型分布有关。调查显示木莲九连山种群海拔分布最高线为1000m左右,个体多度(n=534)在800m左右最高,核密度也在700—900m范围达最大。而在1000m最高界线以内,木莲的生境适宜度伴随海拔而升高,两者呈显著正线性关系(图4)。不仅如此,调查也发现木莲植株仅在当地常绿阔叶林与常绿落叶混交林集中出现,尤其后者多度、核密度均显著高于前者。当然,常绿针叶林也能为木莲提供适宜生境(4图4),但实际调查却鲜能遇见木莲植株(见图4),说明除了海拔要求外,野外木莲发生对植被或生境类型也有要求。
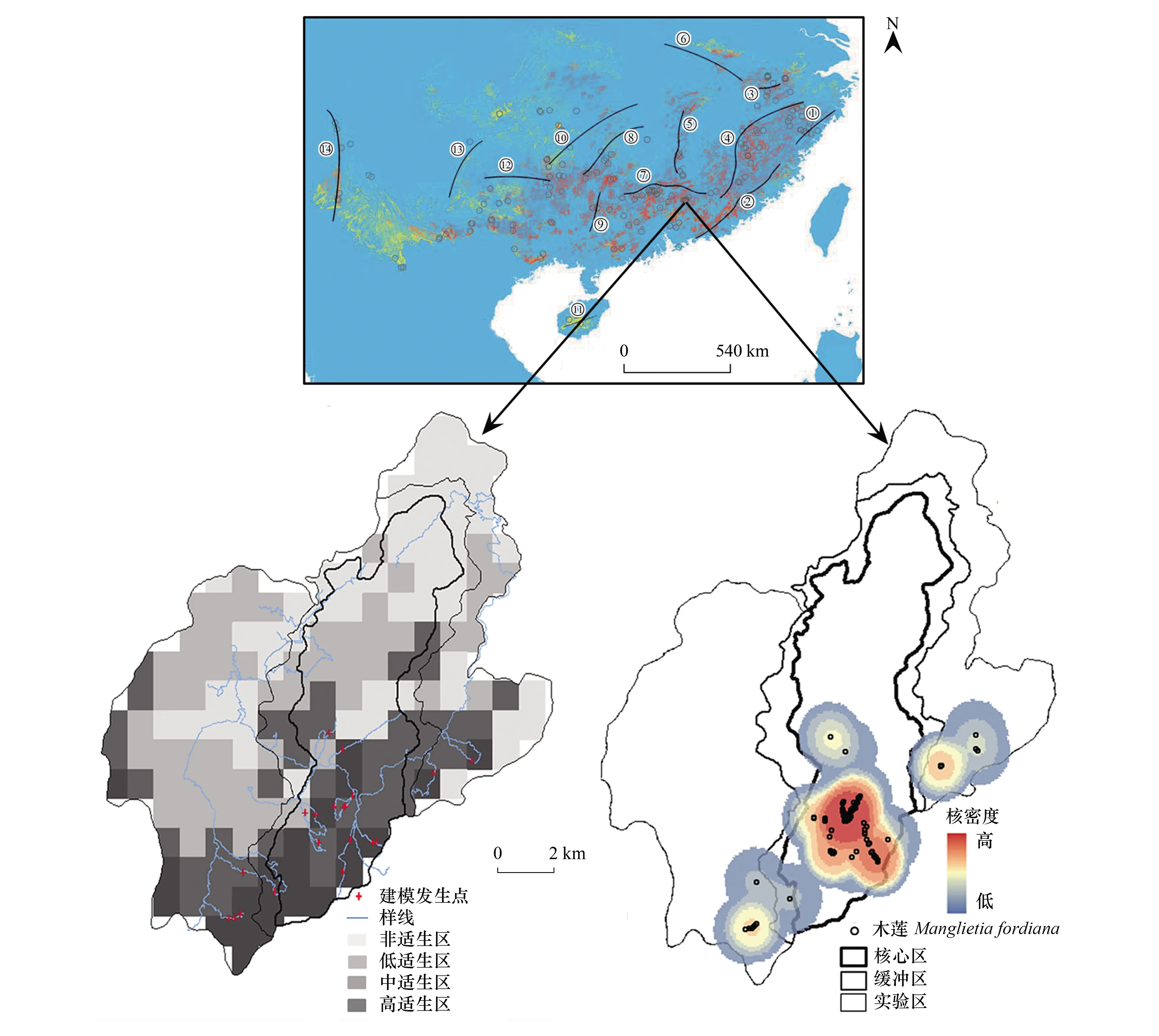
图3 木莲M.fordiana九连山种群调查样线、植株位点及其适宜生境预测与核密度中心分布

图4 海拔高度、植被类型影响木莲M.fordiana九连山种群的多度、核密度与生境适宜度(以气泡大小表示值的高低)
2.4 基于木莲适生分布的保护与空缺分析
木莲预测适生区(D)中高、中、低适生区占比分别为35.8%、23.2%、41%(表2),选取的主体区(Dm,占D的83.9%)及其阔叶林提取部分(BDm)的适生等级占比均与D一致(x2=3.188,P=0.527)。然而,Dm高适生占比(39.9%)升高,尤其BDm的高适生占比(48.2%,表2)明显提升,意味着经阔叶林提取后的木莲适生分布BDm较Dm更接近实际,对保护空缺分析有利(如误差下降)。

表2 木莲预测适生区、主体适生区及其各适生等级的栅格数与占比
2.4.1木莲的保护空缺分析
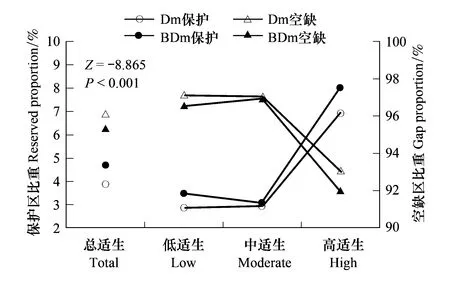
图5 木莲预测适生主体区(Dm)与阔叶林提取主体区(BDm)各等级适生区的保护与空缺比重
从图5可知,分别基于Dm与BDm的保护空缺分析,所得结果差异显著(Z=-8.865,P<0.001,图5),基于BDm的保护比重均高于基于Dm的保护比重,尤其高适生区受保护比重从Dm的6.95%提高至BDm的8.03%,空缺比重则正好相反(图5)。不仅如此,从图6可知,保护或空缺分析也受海拔梯度影响,海拔越高保护比重越高(或空缺比重越低)。例如, 基于BDm(不含低适生区)的木莲适生区保护比重在海拔>700m的区域保护比重最高(8.7%),其次为海拔400—700m区域(4.4%),而<400m区域的保护比重最低(2.3%),空缺比重与之相反(图6)。事实上,从受保护的木莲发生点分布来看,大部分保护点也位于海拔>700m区域,而仅少数位于低海拔地带(图6)。
结合木莲保护空缺点与BDm包括空缺区的分布,考虑山脉隔离下的地理种群保护覆盖的需要,大致识别出7个保护空缺区,即雁荡山区、黄山-天目山区、苗岭东段、高黎贡山北段与南段、中越边界山区与广东西南山区。而若基于木莲的>700m优先区保护原则,仍有黄山-天目山区、苗岭东段、高黎贡山北段与南段、中越边界山区与广东西南山区6个空缺区需保护覆盖(图6)。
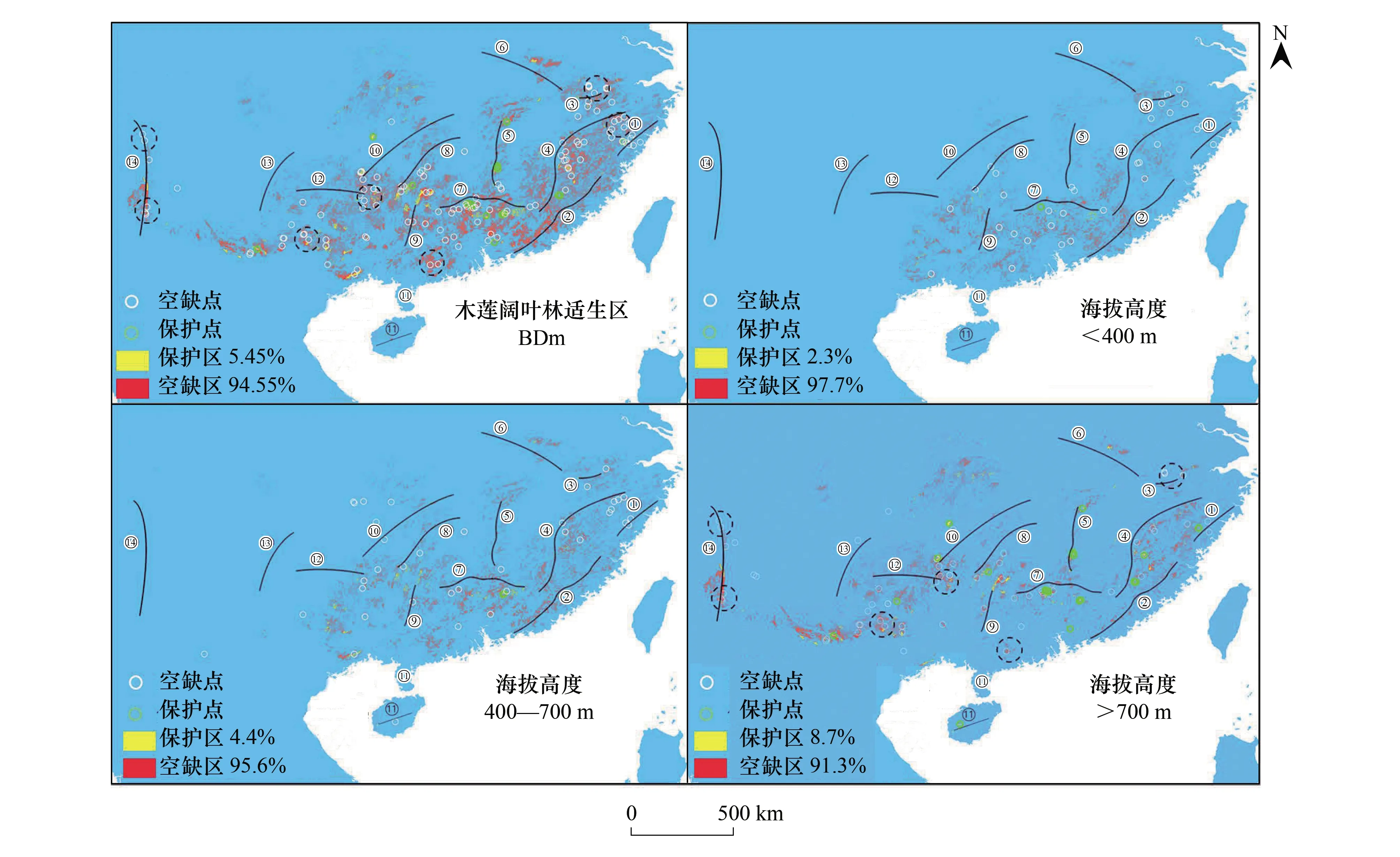
图6 不同海拔高度的木莲阔叶林适生区及其发生点的保护/空缺分布与识别
2.4.2保护区的生境保护:以九连山种群为例
从表3可知,九连山保护区内70%为木莲适宜生境,其中高适宜生境占14%、中适宜占24%、低适宜占32%。适宜生境在核心区、缓冲区、实验区3个功能区的分布比重差异显著(Friedman检验:x2=221.9,P<0.001),例如,51.4%的高适宜生境位于核心区;而其余适宜等级则均以实验区占比最大,包括47.1%的中适宜生境、68.8%的低适宜与55.3%的非适宜生境均在实验区。所以,总体上,实验区适宜生境占比最高(55.4%)(图3,表3)。

表3 木莲适宜生境在九连山自然保护区核心区、缓冲区、实验区的分布比重
3 结论与讨论
我国南方生物多样性主要依赖于山地阔叶林生境,优先保护山地阔叶林才能维持现有的生物多样性热点[10]。当前对这类重要生态系统的有效保护不足,自然保护区在优先区域的覆盖比重偏低[34]。例如,本次研究结果显示,自然保护区对典型南方山地阔叶植物木莲的适生范围的覆盖率为4.1%至6.0%,需分析其优先区域的保护空缺,以优化自然保护区在山地阔叶林的空间布局[6]。
多数木兰科种类的发生依赖于山地阔叶林生境(尤其是常绿型)[32],而呈现出局部聚集或区域零散状发生模式,这可能与一些特定的生境条件需求有关,由此限制了它们的地理分布。从本次采集的发生点可知,木莲为东亚山地特有种,尤其集中在武夷山脉、南岭、苗岭等南方中山地带发生,似乎与木莲属的云南分布中心[11—12]不一致,但可能都受益于喜马拉雅山脉抬升运动对大气环流与区域气候的影响。南方中山地带常年降水丰沛,受湿润性季风气候影响(如最暖季降水量bio18贡献度较高)。尤其,干季仍能满足木莲所需的30—55mm/月的生态位需求,对其种群发生可能至关重要,是影响木莲适生分布的最关键生态因子(如最干月降水量bio14贡献度最高)。此外,这些地带特定的中山地形对于古老植物木莲的长期保存也至关重要。尤其,当人类活动日益频繁、生境丧失加快[35],较高的海拔与起伏的地势,能减缓人类活动强度,降低破坏频次,从而保存更多木莲植株或种群发生所需的生境。九连山案例显示,当前木莲种群的发生中心受当地海拔高度及植被类型分布影响显著,表现出人为干扰后的选择迹象。例如,当地木莲种群偏向于在700m以上海拔高度聚集发生,而大量植株个体出现在干扰强度更低的常绿阔叶林与常绿落叶混交林发生,尤其是后者的原生性,使其成为当前木莲的核密度中心。近些年,物种分布模型或生态位模型(如MaxEnt)在保护热点/优先区、空缺分析上应用较广[33,36—38],基于气候、地形等环境变量的生态位模型及其适生区分布预测,能在一定程度上代表物种基础生态位[39],反应其适生性特征,并获知影响物种分布的关键或约束性环境因子。例如,木莲适生性受最干月降水量(bio14)、海拔高度、温度季节性变化标准差(bio4)、最暖季降水量(bio8)以及最冷月最低温度(bio6)联合影响,本次揭示其bio14生态位为30—55mm、海拔为400—1500m、bio18为500—800mm等,意味着北方冬季的干燥寒冷限制了它的扩散、分布,而南方温暖、湿润的山地气候则为其提供了适宜生境。当前,80%木兰科物种聚集在亚洲南部、东南部[14,21],也类似地反应了对这种温暖湿润山地气候的依赖,而这很大程度上归功于当地特有的喜马拉雅山抬升运动,促使形成了特殊的湿润性夏季季风模式[40]。
然而,生态位模型预测结果反映的是基础生态位[12],并非实际生态位[20]。而且,根据保护管理的成本效益[41]、“最小种群”[42]或“最小面积”[43]要求,进行保护空缺分析前,需在适生区基础上,叠加考虑植被类型、物种分布与热点图,再与现有土地所有权、管理状态图叠制[38,44],获得优先区及其保护覆盖后,再进行空缺分析。考虑到当前多数野外木兰科植物限制在阔叶林生境发生[12],因而我们将阔叶林分布与木莲适生区分布叠加,以考虑实际生境分布。事实上,考虑实际生境后,其适生区面积缩减了75.6%,有利于提高后续空缺判断、识别的针对性。通常,自然保护区设计目标是以最小的面积来保护当地最大的生物多样性[45],因而需首先考虑生物多样性优先区、热点区的分布[46]。从木莲的各适生等级的保护比重来看,现有自然保护区在规划、设计上遵从优先区、热点区原则,其保护比重在高适生区(相当于优先区或热点区)最高(空缺最低,见图6)。同样地,沿海拔分布梯度的保护比重也呈现类似情况。九连山案例显示木莲集中分布于海拔700m以上区域,400m以下几乎不发生(见图2),这与大尺度下适生与海拔生态位(如400—1500m)较一致。比较<400m、400—700m与>700m海拔组,也发现>700m组的保护比重最高,说明现有保护区倾向于保护山地环境,而实际上,该区域的生物多样性热点生态区(如横断山脉、南岭山脉、中国东部山脉等)也多数分布在这类生境[10]。
但是,木莲适生区总体保护覆盖有限(<10%),即便在>700m的发生热点区,保护比重也<10%(如8.7%)。主要原因可能在于该区域山地面积广阔,分布着武夷山、南岭、罗霄山、苗岭、大别山等14座以上中山山脉,而通常单一的保护区在面积上很难做到山脉全覆盖,比如九连山横跨广东与江西两省,保护区划分直接采用山脊行政界线方法,未能考虑完整山体的保护要求。山地保护区的规划、设计常面临许多挑战,如“最小面积”[43]如何界定等问题,然而我们仍可通过新增保护区的形式,不断优化区域保护网络。
运用系统保护规划概念、方法测定保护地有效性[16—17],首要步骤是确定生物多样性优先区,之后与现有保护区比较分析[18],找出保护空缺,在空缺地建立保护区[1]。而山地阔叶林优先区的确定,或许要考虑海拔最低线,并根据保护对象,筛选出最适海拔范围。以木莲为例,本次将>700m作为其最适海拔要求,即保护优先区,并借助主要山脉走向、位置,通过将具有地理山脉隔离的地理种群作为保护空缺识别参考,判定获得区域内阔叶林保护空缺6个,如黄山-天目山区、苗岭东段、高黎贡山北段与南段、中越边界山区与广东西南山区等。本次未基于单元网格保护覆盖探寻空缺,而是将发生点与优先保护区结合,并从地理种群保护层面,识别空缺所在的大致位置、区域,这对于大尺度上的保护规划与保护区网络建设、优化,同样具有启示作用。不仅如此,对于物种多样性保护而言,也有利于提高物种种下多样性保护的有效性。
根据生物圈的自然保护区概念,保护区常包括核心区、缓冲区与实验区或过度区3部分[47],承担各自功能、目标,管理措施方法也不同。如核心区一般采用严格控制措施,而实验区则采用适度控制等方法。而根据适应性管理要求[48],高适宜生境归入核心区严格管理,九连山案例显示 >50% 木莲高适宜生境在核心区,符合适应性管理要求,生境保护总体有效。但实验区仍有较高比重的中适宜生境及相对比重的高适宜生境,与适应性管理不相称,如可能不利于低海拔生境恢复与木莲扩散及其种群扩增等。
本次以木莲的保护与空缺为例,探讨我国南方山地阔叶林的保护现状。保护覆盖沿海拔梯度增加而增加,意味着前期保护区规划、设计较好地遵从了优先区概念。然而,在保护区布局上来看,呈现片段化分布,在保护网络设计上考虑不足,这可能土地利用矛盾妥协的产物。多数保护区设计未遵从最佳方案,如保护区形状(圆形优于带状)、缓冲区面积偏小等,使得功能区之功能不易发挥出来。结合木莲 >700m优先区与山脉分布,从地理种群保护需求识别空缺,为我国南方山地阔叶林保护网络建设提供优化方向。
