黄土高原藓结皮土壤呼吸速率对降雨量变化的响应
2022-03-30窦韦强田乐乐姚小萌李胜龙
窦韦强,田乐乐,肖 波,姚小萌,3,李胜龙
1 中国农业大学土地科学与技术学院农业农村部华北耕地保育重点实验室, 北京 100193 2 中国科学院水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100 3 河北师范大学资源与环境科学学院环境演变与生态建设实验室, 石家庄 050024
土壤呼吸是指土壤由于代谢作用释放CO2的过程,是陆地生态系统碳循环的重要环节之一,约占整个生态系统呼吸的60%—90%,每年释放的CO2高达到68—100 Pg[1—3]。因此,土壤呼吸的微小变化都会显著影响大气CO2浓度和土壤碳的累积量,进而影响陆地生态系统的碳源/汇功能[4]。干旱和半干旱地区作为陆地生态系统的重要组成部分,约占全球陆地总面积的41%[5]。由于该区域降水稀少,维管束植物生长受限,而由藻类、地衣、苔藓等隐花植物和土壤微生物等通过假根、菌丝体、胞外分泌物与土壤表层颗粒胶结形成的具有代谢活性的生物结皮广泛发育[6—7],覆盖度最高可达60%以上[8]。
作为干旱和半干旱地区重要地表覆被,生物结皮可影响土壤水分循环、养分循环、碳氮固存、水土流失、生物多样性以及植被恢复等地表生态过程[9—12],特别是对土壤呼吸的影响不容忽视。通常认为,生物结皮的光合自养成分(如蓝藻、地衣和苔藓)能促进自养呼吸增加生物结皮覆盖土壤CO2的排放,其还会改变土壤温度、含水量、酶活性、微生物群落组成和结构等,间接影响土壤呼吸[13—14];此外,生物结皮层的致密结构还会堵塞土壤孔隙,阻碍结皮下层土壤与大气CO2的交换,进而影响土壤呼吸[15—16]。可见,干旱和半干旱地区生物结皮对土壤呼吸具有显著影响。然而,与单位面积的维管束植物相比,尽管生物结皮土壤呼吸速率相对较低,但其覆盖了地球陆地表面的12%[17],相对较高的覆盖面积可能会显著增加其对干旱和半干旱地区土壤呼吸的总贡献。因此,评估陆地生态系统土壤碳动态平衡时,应该充分考虑干旱和半干旱地区生物结皮对土壤呼吸的影响。
土壤水分是调控干旱和半干旱地区生物结皮土壤呼吸的首要限制因子[18],而降雨是该区土壤水分补充的主要途径。降雨导致的土壤水分变化使土壤发生频繁的干湿交替过程,而这一过程将改变土壤团聚体、微生物活性和群落结构等,进而影响生物结皮土壤呼吸[19]。研究表明[20—21],当无降雨供给时,生物结皮表层受干燥胁迫,土壤微生物处于休眠状态,生物结皮土壤呼吸速率较低;当降雨发生时,土壤水分增加会迅速激活土壤微生物从而打破休眠状态,同时还会破坏团聚体结构,释放出大量呼吸底物,致使生物结皮土壤呼吸速率迅速升高;而当降雨过大时(如极端降雨),降雨造成土壤水分饱和或积水淹没生物结皮层,阻碍O2进入使土壤从好氧转变为厌氧环境,从而抑制微生物活动,降低生物结皮土壤呼吸速率[22—24]。可见,生物结皮土壤呼吸速率对降雨量变化的响应异常敏感。而根据气候模型预测,未来干旱和半干旱地区降雨模式将发生变化,极端降雨和干旱延长事件的频率与幅度将不断升高[25]。因此,明确未来降雨模式下生物结皮土壤呼吸的响应规律对干旱和半干旱地区土壤碳循环和碳通量研究具有重要意义。
迄今,有关干旱和半干旱地区降雨量改变对生物结皮土壤呼吸的影响已有部分报道,但基于该区土壤碳排放的重要性、土壤呼吸影响因素的复杂性以及时空变异性等[26—27],生物结皮土壤呼吸对降雨量改变的响应仍需进一步深入探究。黄土高原作为我国典型的干旱和半干旱地区,自20世纪90年代起实施大面积退耕还林(草)工程,其后放牧减少,生物结皮大面积发育,部分地区覆盖度可达60%—70%[28—29]。基于此,本研究针对黄土高原风沙土发育的藓结皮,以自然降雨量为对照,分别进行增雨10%、30%、50%和减雨10%、30%、50%的处理,并利用便携式土壤碳通量分析仪测定增雨和减雨下的藓结皮土壤呼吸速率以及表层土壤温度和含水量,对比分析增雨和减雨下藓结皮土壤呼吸速率的响应规律及其与土壤温度和含水量的数量关系,以期为黄土高原地区未来降雨模式变化情景下土壤碳的动态响应提供参考依据。
1 材料与方法
1.1 研究区概况
研究区位于黄土高原北部陕西省神木市六道沟流域(110°21′—110°23′E,38°46′—38°51′N),海拔1094.0—1273.9 m,面积大约6.9 km2,属中温带半干旱气候,雨热同季,年均气温8.4℃,年均降雨量和蒸发量分别为408.5和1337.0 mm,其中6—9月降雨量约占全年降雨量的80.9%。该区域属黄土丘陵区向鄂尔多斯高原过渡的水蚀风蚀交错带,是黄土高原水土流失最为严重的地区[30]。土壤质地为砂土和砂质壤土,多年退耕还林(草)过程中,植被恢复良好,柠条锦鸡儿(CaraganakorshinskiiKom.)、油松(PinustabuliformisCarr.)、沙柳(Salixcheilophila)及苜蓿(MedicagosativaLinn.)等分布广泛,同时生物结皮大面积发育,且以藓结皮为主,受人为干扰较小。
1.2 降雨量处理实验设计
实验分别于2018-08-19—9.23和2019-06-25—8-25进行。根据野外实际调查结果,于2018年7月在研究区风沙土上选择地势较为平坦、远离植物灌丛、藓结皮发育良好的开阔样地,采用随机区组实验设计,共设定21个小区(2 m×2 m),每个小区间隔3 m。为防止地表径流流入,小区四周被嵌入地下20 cm的铁皮形成隔离带包围,样地概况见表1。设定的21个小区共包括7种降雨量处理:自然降雨量(对照)、减雨10%、30%、50%和增雨10%、30%、50%,每种处理3个重复。

表1 生物结皮样地概况
实验采用自制减雨架和人工喷洒的方式进行减雨和增雨处理。如图1所示,减雨架主要由夹角60°和直径5 cm的瓦面状透明PVC板凹槽挡雨面、夹角15°和直径20 cm的半圆状PVC管集雨槽、高70 cm和外上口径56 cm的集雨桶以及两侧高度分别为1.0 m和1.5 m的金属支架组成。根据减雨梯度,实验在减雨处理小区上方均匀朝上设置一定数量直径5 cm的瓦状透明PVC板凹槽挡雨面达到减雨效果(挡雨面与样方面积之比即为减雨梯度)。每次降雨前,将集雨桶的盖子固定并留出与集雨槽的连接口,其余封闭,以免雨水进入;每次降雨结束后,依据减雨梯度将收集在集雨桶中的雨水通过手提式喷雾器在样方50 cm高度来回均匀喷洒到对应的增雨处理小区,形成增雨处理,喷洒速度以避免产生地表径流为准。实验中由多人配合对增雨处理样点同时喷水和测量,以避免时间先后差异对结果的影响。

图1 增雨和减雨处理实验小区
1.3 测定指标及方法
1.3.1降雨量
采用翻斗式自动雨量计(TE525M)测定。
1.3.2土壤呼吸速率、表层土壤温度和含水量
土壤呼吸速率:在各处理样方中心随机安装1个直径为20 cm、高度12 cm的土壤呼吸环,插入土壤中9 cm,露出地面3 cm。为减少土壤呼吸环安装对土壤结构的影响,安装完成后稳定一个月再进行测量。每种降雨处理结束后等待5 min,使用便携式土壤碳通量分析仪(LI-8100A)对每个样点的土壤呼吸速率依次测定(测定时间2 min)。测量完成后定期清理样方内植物和枯枝落叶,以保证不干扰后续测定。
表层土壤温度和含水量:测定土壤呼吸速率的同时,使用便携式土壤碳通量分析仪自带的土壤温湿度探头同步测定表层5 cm的土壤温度和含水量,每次测量时插入土壤,测量完成后取出。
1.3.3藓结皮基本特性
各处理结皮厚度采用游标卡尺测定;藓植株密度利用镊子夹取单位面积内的苔藓植株计数测定;藓生物量采用烘干法测定;结皮盖度采用网格法测定。不同降雨量处理下藓结皮的基本特性见表2。

表2 不同降雨量处理下藓结皮的基本特性
1.4 数据处理与分析
采用Excel 2016对数据进行整理分析,使用SPSS Statistics 22进行t检验和单因素方差分析,利用OriginPro 9.2进行图表绘制和数据拟合。
生物结皮土壤呼吸速率与土壤温度的关系利用指数回归模型拟合[13],形式如下:
Rs=R0ebT
式中,Rs表示土壤呼吸速率(μmol m-2s-1);R0表示温度为0℃的土壤呼吸速率(μmol m-2s-1);T表示表层5 cm土壤温度(℃);b表示温度反应系数。
生物结皮土壤呼吸速率与土壤含水量的关系使用线性模型拟合[31—32],形式如下:
Rs=aW+b
式中,Rs表示土壤呼吸速率(μmol m-2s-1);W表示表层5 cm的土壤含水量(%),a、b均为常数。
2 结果与分析
2.1 不同处理的降雨量统计分析
如表3所示,整个实验周期共有24次自然降雨事件出现,其中2018年10次,7种处理的总降雨量范围为37.3—111.9 mm,平均降雨量范围为3.7—11.2 mm,最大降雨量范围为10.3—30.8 mm,最小降雨量范围为0.7—2.0 mm,整体变化幅度较小;2019年降雨14次,7种处理的总降雨量范围为68.2—204.5 mm,平均降雨量范围为4.9—14.6 mm,最大降雨量范围为24.0—72.0 mm,最小降雨量范围为0.35—1.05 mm,整体变化幅度较大。总体来看,整个实验周期不同降雨处理以小降雨(<10 mm)事件为主,占总降雨次数的约72%。

表3 不同处理的降雨量统计分析
2.2 土壤温度和含水量对降雨量变化的响应
如图2所示,不同降雨量处理下土壤温度整体呈先升高后降低(2018)和“多峰”(2019)变化趋势;土壤含水量整体呈“多峰”(2018和2019)变化趋势。整个实验周期(2018和2019)不同降雨量处理下土壤温度表现为减雨50%>减雨30%>减雨10%>自然降雨量>增雨10%>增雨30%>增雨50%,与自然降雨量相比,虽然增雨(增雨10%、30%、50%)和减雨(减雨10%、30%、50%)分别会降低(降幅分别为0.5%—2.8%和0.4%—2.6%)和升高土壤温度(增幅分别为2.8%—3.7%和1.7%—2.6%),但单因素方差分析表明,增雨和减雨下土壤温度均无显著差异,说明降雨量变化对土壤温度的影响不显著。此外,整个实验周期不同处理下的土壤含水量表现为增雨50%>增雨30%>增雨10%>自然降雨量>减雨10%>减雨30%>减雨50%。与自然降雨量相比,增雨和减雨分别提高(增幅分别为17.9%—48.2%和27.1%—54.2%)和降低了(降幅分别为1.8%—26.8%和5.2%—20.8%)土壤含水量,且单因素方差分析表明增雨和减雨下土壤含水量差异显著,其中2018年增雨50%显著高于减雨30%、50%,2019年增雨50%、30%显著高于自然降雨量、减雨10%、30%和50%。另外,2018年的土壤含水量显著低于2019年。

图2 不同降雨量处理间的土壤温度和含水量差异
2.3 藓结皮土壤呼吸速率对降雨量变化的响应
由图3可得,不同降雨量处理下藓结皮土壤呼吸速率整体呈“多峰”变化趋势。整个实验周期不同降雨量处理下土壤呼吸速率为减雨10%>减雨30%>自然降雨量>增雨10%>减雨50%>增雨30%>增雨50%。与自然降雨量相比,增雨和减雨对土壤呼吸速率的影响存在明显差异,其中增雨会抑制土壤呼吸速率(降幅分别为7.8%—31.7%和14.7%—39.4%),且随梯度增大抑制作用越明显;而减雨取决于减雨梯度,减雨10%和30%会促进土壤呼吸速率(增幅分别为27.5%、9.6%和23.6%、9.7%),减雨50%会抑制土壤呼吸速率(降幅分别为15.6%和18.5%)。单因素方差分析表明,增雨和减雨下土壤呼吸速率存在显著差异,其中2018年减雨10%的土壤呼吸速率显著高于增雨30%和50%,2019年减雨10%的土壤呼吸速率显著高于自然降雨量,增雨30%、50%和减雨50%显著低于自然降雨量。此外,除增雨30%、50%外,其余降雨量处理下2018年的土壤呼吸速率显著低于2019年。
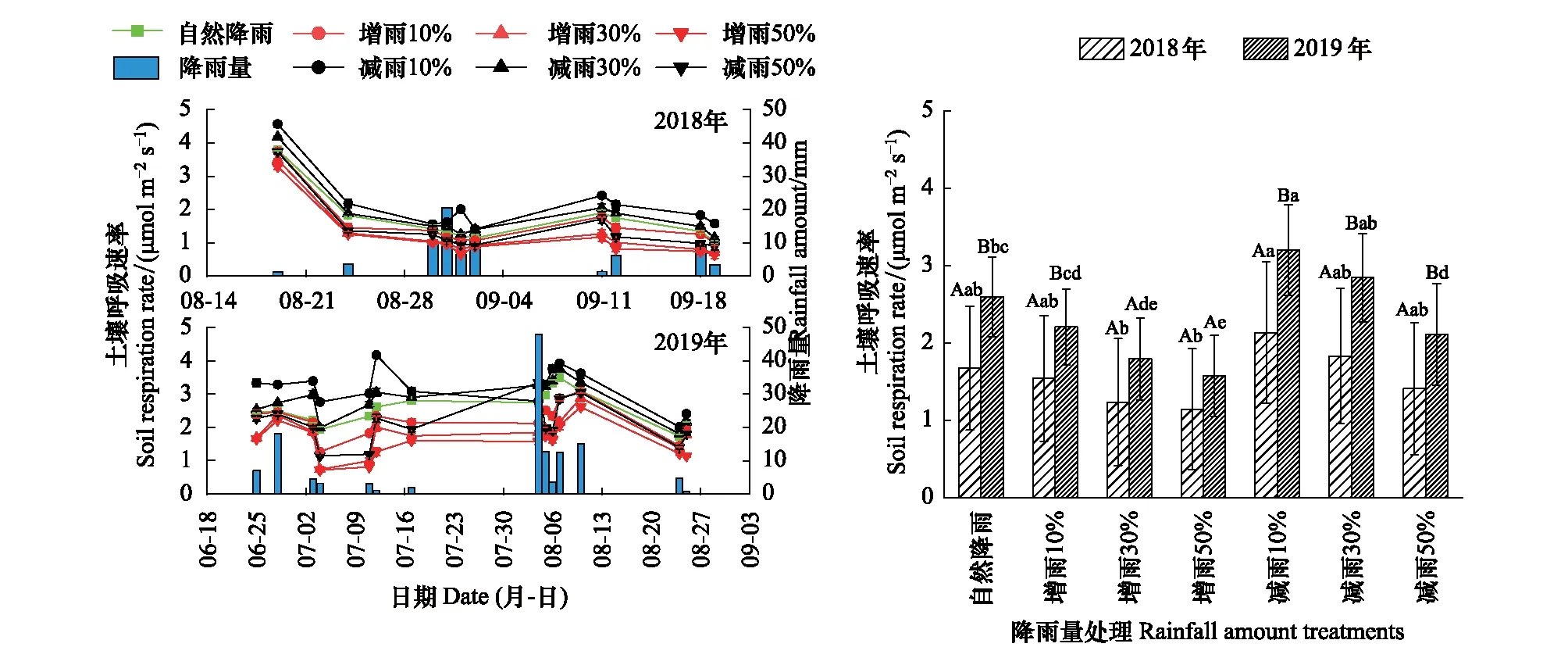
图3 藓结皮土壤呼吸速率对降雨量变化的响应
2.4 降雨量变化下土壤温度与藓结皮土壤呼吸速率的关系
如图4所示,采用公式(1)的指数模型对7种降雨量处理下的土壤温度和藓结皮土壤呼吸速率拟合发现,7种处理下藓结皮土壤呼吸速率均随土壤温度的升高呈指数增加,二者成显著指数正相关关系,增雨和减雨下的土壤温度可分别解释藓结皮土壤呼吸速率92.6%—95.0%和85.7%—95.9%的变化(表4)。可见,增雨和减雨下土壤温度增加对藓结皮土壤呼吸速率具有正效应,且降雨量变化对土壤温度和藓结皮土壤呼吸速率的关系无影响。

图4 不同降雨量处理下藓结皮土壤呼吸速率与土壤温度的关系

表4 不同降雨量处理下藓结皮土壤呼吸速率与土壤温度的拟合方程
2.5 降雨量变化下土壤含水量与藓结皮土壤呼吸速率的关系
由图5可得,当土壤含水量小于约15%时,7种降雨量处理下土壤含水量与藓结皮土壤呼吸速率的关系表现出一致性,均成显著线性正相关关系,即藓结皮土壤呼吸速率随土壤含水量的增加而快速线性增加,增雨和减雨下的土壤含水量可分别解释藓结皮土壤呼吸速率88.9%—98.7%和86.2%—92.4%的变化(表5);当土壤含水量超过约15%时,增雨下藓结皮土壤呼吸速率随土壤含水量的增加迅速降低,二者成显著线性负相关关系,土壤含水量可解释藓结皮土壤呼吸速率73.1%—99.2%的变化(表5)。当土壤含水量约等于15%时,增雨下藓结皮土壤呼吸速率达到峰值,分别为3.55、3.26和2.89 μmol m-2s-1。可以看出,低含水量时增雨和减雨下土壤含水量增加对藓结皮土壤呼吸速率具有正效应,而高含水量时增雨下具有负效应,最适含水量阈值约为15%。

图5 不同降雨量处理下藓结皮土壤呼吸速率与土壤含水量的关系

表5 不同降雨处理量下藓结皮土壤呼吸速率与土壤含水量的拟合方程
3 讨论
3.1 降雨量变化对藓结皮土壤呼吸速率的影响
土壤水分是限制干旱和半干旱地区生命活动的主要非生物因素,降雨不仅是土壤水分的主要来源,而且对地下生物化学过程具有重要的调控作用[33]。降雨模式的变化会直接影响土壤水分有效性进而影响包括土壤呼吸在内的陆地生态系统碳循环。本研究结果表明,藓结皮土壤呼吸速率对降雨量变化具有明显响应,但其规律不同。与自然降雨量相比,增雨会抑制土壤呼吸速率,且随梯度增加抑制作用越明显,这可能与增雨前的土壤水分状态和增雨导致的土壤水分含量显著增加有关。研究表明,土壤水分相对亏缺时对土壤呼吸速率具有促进作用而饱和时具有抑制作用[34—35]。本研究增雨是在自然降雨的基础上进行,使得增雨前土壤水分含量相对较高,土壤处于湿润状态,而增雨又显著增加了土壤含水量(图2),造成的土壤水分饱或积水可能会填充土壤孔隙并淹没生物结皮层,一方面使土壤CO2不能轻易释放,限制O2进入土壤使其处于还原状态,抑制微生物活动[22];另一方面还会降低其有效光合面积,影响光合产物在根系的分配,从而降低土壤呼吸速率[36]。如赵洋等[37]研究表明,腾格里沙漠东南缘极端降雨(44.7 mm)后生物结皮土壤呼吸会受到明显抑制,其呼吸速率会下降40.0%—58.3%;肖波等[38]研究发现,黄土高原藓结皮土壤呼吸速率在6—40 mm降雨量范围随降雨量增加而迅速降低;Davidson等[39]也得到大的降雨事件后土壤呼吸速率会受到明显抑制。此外,增雨还可能与自然降雨形成连续降雨,吴林等[40]研究认为初次降雨会消耗更多的土壤呼吸底物导致二次降雨的土壤呼吸速率下降。
减雨对藓结皮土壤呼吸速率的影响取决于减雨梯度,减雨10%和30%会促进土壤呼吸速率,而减雨50%会抑制土壤呼吸速率(图3)。本研究整个实验周期以小降雨事件(<10 mm)为主(约72%),减雨10%和30%处理下降雨量整体减小,但干旱半干旱区生物结皮的土壤微生物在长期干湿交替过程中已经形成了一种快速而强烈的响应机制,一旦降雨发生,土壤微生物会迅速打破休眠机制从而恢复活性,加速有机质等底物的分解和释放;此外,减雨前土壤处于相对干旱状态,因水分缺乏导致土壤微生物大量死亡,土壤团聚体中积累了丰富且易被分解的有机物,减雨导致的土壤湿化过程可以破坏团聚体结构,使以前被保护而不可利用的有机底物暴露,并加速其分解从而提高土壤呼吸速率[19]。如管超等[41]认为小于5 mm的降雨能刺激生物结皮中土壤微生物的新陈代谢活动,增加其生物量,从而提高土壤呼吸速率;肖波等[38]研究表明黄土高原地区小于6 mm降雨范围内藓结皮土壤呼吸速率随降雨量增加呈直线上升;韩海燕[42]在高寒沙区模拟1、2、5 mm降雨后发现藓结皮土壤呼吸速率较降雨前分别升高了11.0、6.2、4.1倍。但降雨量过低时可能不会诱发上述过程,如彭信浩等[43]和周世兴等[44]研究认为短期的极端减雨会明显抑制土壤呼吸。另外,本研究减雨10%和30%下的藓结皮基本特性(厚度、植株密度、生物量以及盖度)明显高于自然降雨量(表2),表明适当的减雨更利于生物结皮的发育,而Miralles等[5]研究表明物种丰度的增加将进一步增加土壤碳的释放量。相关研究表明,未来黄土高原地区降雨量整体呈波动减少的趋势[45],这使生物结皮可能成为一潜在碳源,未来降雨量变化下准确评估该地区土壤碳源/汇功能时应考虑生物结皮土壤呼吸的影响。
3.2 降雨量变化下土壤温度与藓结皮土壤呼吸速率的关系
研究证实,黄土高原地区藓结皮土壤呼吸速率日变化曲线与土壤温度日变化曲线具有高度一致性,说明土壤温度与土壤呼吸速率高度相关[46]。一般而言,土壤温度与土壤呼吸速率成正相关关系,土壤呼吸速率随土壤温度的升高而快速增加,如Yao等[13]研究得到藓结皮土壤呼吸速率随土壤温度的升高而显著增加,二者成显著指数正相关关系;Oberbauer等[47]研究认为温度升高显著促进了干旱苔原生态系统的呼吸作用;Maestre等[48]研究也表明半干旱区增温显著提高了生物结皮覆盖的土壤呼吸速率。而本研究也得到了类似结果,增雨和减雨下藓结皮土壤呼吸速率与土壤温度均成显著指数正相关关系(图4),即藓结皮土壤呼吸速率随土壤温度的升高呈指数型增加,这是因为温度升高能刺激土壤酶的活性从而加速有机质分解,促使土壤呼吸速率升高。然而,当土壤水分受限时,土壤温度升高反而会降低土壤呼吸速率,这是因为干旱和半干旱地区受水分制约明显,土壤水分几乎是所有生态过程的第一驱动力,增温会降低土壤水分的有效性进而抑制微生物活性,从而影响土壤呼吸速率。如管超等[41]在腾格里沙漠东南缘的研究发现,随土壤温度升高藓结皮和藻—地衣结皮的土壤呼吸速率呈下降趋势;徐冰鑫等[49]在沙坡头天然植被区的研究也得到了类似结论。另外,由于荒漠生态系统有机质贫乏,缺少根系呼吸等自养呼吸,增温还会通过降低异养呼吸来影响土壤呼吸速率[50]。
3.3 降雨变化下土壤含水量与藓结皮土壤呼吸速率的关系
本研究结果表明,当土壤含水量低于约15%时,增雨和减雨下藓结皮土壤呼吸速率与土壤含水量均成显著线性正相关(图5),但二者斜率存在一定差异,即相同土壤含水量时增雨和减雨下的土壤呼吸速率不同,这可能与增雨和减雨下藓结皮的自身特性变化有关。如Lange等[51]研究表明,即使在给定的湿度水平下生物结皮对土壤碳通量的影响也可能因物种的内在因素而异,如生物结皮结构、生物量、颜色以及热性质等,而本研究中不同降雨量处理下的藓结皮基本特性存在一定差异(表2),因此,可能导致了土壤呼吸速率的差异。当土壤含水量超过15%时,增雨下二者成显著线性负相关,表明增雨对藓结皮土壤呼吸速率的影响存在最适含水量阈值,约为15%。由于干旱和半干旱地区降水稀少,蒸发强烈,土壤长期处于干旱状态,水分是限制土壤呼吸的主要因子,减雨会显著降低土壤含水量,同时增雨下的土壤含水量低于最适含水量阈值时土壤水分仍是影响土壤呼吸的关键因子,因此最适土壤含水量阈值范围内,增雨和减雨下藓结皮土壤呼吸速率均随土壤含水量增加线性增加。如董雄德等[31]和Huang等[52]研究得到土壤含水量与土壤呼吸成显著线性正相关关系;Xu等[53]和胡宜刚等[54]研究表明随土壤含水量增加土壤呼吸速率呈快速线性增长。然而,超过最适含水量阈值范围时,增雨下土壤含水量与藓结皮土壤呼吸速率成显著线性负相关,此时土壤水分已不是限制土壤呼吸的主要因子,甚至还会通过堵塞土壤孔隙形成厌氧环境显著抑制土壤呼吸,如肖波等[38]研究得到随土壤含水量增加藓结皮土壤呼吸在低含水量时具有正效应,但在高含水量时具有负效应。另外,还有研究认为土壤含水量与土壤呼吸速率并非线性相关关系,如李新鸽等[55]和吴林等[40]研究表明二者成二次函数关系;与此类似,张鹤等[56]和辜晨等[24]认为成对数函数关系。可见,土壤含水量与土壤呼吸速率并非简单的二元函数对应关系,而是受诸多生物和非生物因子的共同影响,仍需进一步深入探究。
4 结论
增雨和减雨显著影响了土壤含水量,但对土壤温度的影响不显著。与自然降雨量相比,增雨和减雨下藓结皮土壤呼吸速率大多差异显著,其中增雨抑制了藓结皮土壤呼吸速率,且随梯度增大抑制作用越明显;减雨则取决于减雨梯度,减雨10%和30%具有促进作用而减雨50%具有抑制作用。增雨和减雨下藓结皮土壤呼吸速率与土壤温度均成显著指数正相关关系,同时,低含水量(小于约15%)时与土壤含水量均成显著线性正相关关系,而高含水量(大于约15%)时增雨下成显著线性负相关关系。总体而言,降雨量变化会显著影响该地区生物结皮土壤碳的排放过程,尤其是降雨量整体波动减小的趋势下其可能成为一潜在碳源。因此,未来黄土高原土壤碳源/汇功能研究中应考虑降雨量变化对生物结皮土壤呼吸的影响。
