新质源两系杂交早稻10个性状的配合力及遗传力分析
2022-03-21谢芳腾刘海平张红林汪雨萍欧阳春荣谢建萍欧阳真程杨福权张家健钟晓英
谢芳腾, 刘海平, 张红林, 汪雨萍, 蒋 龙, 张 璞, 欧阳春荣,谢建萍, 欧阳真程, 杨福权, 张家健, 何 胜, 章 萍, 钟晓英
(1.赣州市农业科学研究所/国家水稻产业技术体系赣州综合试验站, 江西 赣州 341000;2.赣州市农业农村局, 江西 赣州 341000)
目前,全球杂交水稻种植面积已累计超过5 亿hm2,累计增产粮食6.25亿kg,为世界粮食安全做出了重要贡献[1]。两系杂交水稻应用快速提升,已成为杂交水稻重要的组成部分[2]。然而,随着杂交水稻种植面积的扩大,审定通过的杂交水稻品种也越来越多,突破性品种匮乏,杂交水稻单产徘徊不前已成为限制杂交水稻发展的重要因素[3]。以江西为例,江西是水稻种植大省,2018年江西省水稻播种面积仅次于湖南省和黑龙江省,位列全国第三位。2012—2017年江西审定通过的198个水稻品种中186个为杂交水稻品种,其中杂交早稻品种29个,平均单产7 585.53 kg/hm2,比对照增产3.45%。杂交中稻品种52个,平均单产9 019.78 kg/hm2,比对照增产4.40%。杂交晚稻品种105个,平均单产8 209.88 kg/hm2,比对照增产3.41%。在审定通过的190个早中晚稻品种(不含特种稻)中仅有1个品种的穗颈瘟抗性为5级,6个品种的穗颈瘟抗性为7级,剩余183个品种的穗颈瘟抗性均为9级,高感稻瘟病[4]。造成上述问题的主要原因是遗传基础狭窄,相似性品种普遍存在,不仅在产量上难以突破,还导致新育成的品种对病虫害和不良环境的抵抗能力降低。若想进一步提高杂交水稻品种的产量和抗性,当务之急是进行杂交稻亲本的种质创新,丰富水稻种质资源的遗传多样性。本研究采用聚合育种与核辐射诱变育种相结合的方法创制新种质资源,并对其进行配合力分析,旨在为水稻种质创新提供参考。
1 材料与方法
1.1 试验材料
1.1.1不育系
田丰S和广占63 S在不育时期分别与三系保持系D 57杂交,F2进行辐射诱变,在M2选择白化转斑叶型突变体,在短日照低温条件下自交结实,经定向单株选择,聚合水稻早熟、高产等优良基因,最终育成具有白化叶色标记的标田丰S和标广占63 S。需要特别说明的是,广占63 S是典型的晚稻型两用核不育系,但经与D 57杂交、辐射处理后抽穗期提前,成为早籼型两用核不育系。
1.1.2恢复系
R 76、鹏超1号、R 189和R 9311都是中晚稻类型的恢复系,于2012年9月在赣州市农业科学研究所试验基地进行人工去雄杂交,共获得R 76/鹏超1号、R 189/R 9311等24个恢复系改良组合,2012年冬南繁加代,2013年4月成熟后分单株考种,选择优良单株进行核辐射诱变处理,后代采取定向单株选择的方式直至稳定,育成了早籼恢复系R 76/鹏超1号、R 189/R 9311、R 9311/R 76和R 76/R 9311。
2018年6月,以3个光温敏核不育系为母本与6个恢复系(表1)按不完全双列杂交(NCⅡ)遗传设计,配制了18个杂交组合。
1.2 试验方法
2019年3月21日,在赣州市农业科学研究所育种基地播种,4月17日移栽。随机区组设计,3次重复。每小区种10行,每行40株,株行距16.7 cm ×20.0 cm,双本栽插。田间管理按照常规管理进行。
1.3 性状测定
水稻成熟后,连续取小区中间3株考种,考查株高、穗长、单株有效穗、穗总粒数、穗实粒数、结实率、千粒重等7个性状,并实割测产。
长宽比和垩白粒率测定参照郑轶等[5]的方法。
1.4 数据处理
采用DPS 7.05数据处理系统进行方差分析和F测验,配合力分析根据NC-Ⅱ交配设计模型进行。
2 结果与分析
2.1 主要农艺性状的配合力方差分析
由表2可知,除单株有效穗外,其余9个性状组合间的差异均达极显著水平,说明9个性状的组合间存在显著的遗传差异。恢复系除单株有效穗和小区产量外,其余性状的一般配合力方差均达显著水平;不育系株高、穗长、单株有效穗、小区产量和长宽比的一般配合力方差达显著水平,其余性状的一般配合力方差差异不显著。组合间穗总粒数、穗实粒数、结实率、长宽比和垩白粒率的特殊配合力方差达显著水平,其余性状未达显著水平。因此,株高、穗长、千粒重和小区产量的基因型方差全是亲本的加性方差,穗总粒数、穗实粒数、结实率、长宽比和垩白粒率的基因型方差是亲本的加性方差和父母本交互作用产生的非加性方差。
2.2 一般配合力和特殊配合力的相对效应分析
由表3可知,同一亲本不同性状及同一性状不同亲本之间的一般配合力相对效应值具有差异,表明不同亲本不同性状的基因加性效应遗传复杂。对不育系而言,标广占63 S在穗长、穗总粒数、穗实粒数、结实率、小区产量、长宽比和垩白粒率性状上的一般配合力相对效应较好,如要配制穗大、粒多、结实率高、产量高、长宽比大、垩白粒率低的杂交组合,标广占63 S是较好的母本;虔S在株高、单株有效穗和千粒重等性状上的一般配合力相对效应较好,标田丰S在株高、穗长、穗总粒数、穗实粒数、结实率、千粒重、长宽比、垩白粒率的一般配合力效应值均处在第二位,一般配合力中等。从以上结果可以看出,通过辐射诱变与常规杂交相结合选育出的带有叶色标记的不育系与正常不育系虔S相比,一般配合力并无明显降低,甚至标广占63 S大多数性状的一般配合力均处于第一位。对恢复系而言,R 76/鹏超1号在穗长、穗总粒数、穗实粒数、结实率、小区产量、长宽比和垩白粒率等性状上的一般配合力效应较好,是6个父本中最优秀的一个。中早39在株高、穗实粒数、结实率和千粒重上的一般配合力相对效应较好;P 27在株高、结实率、千粒重等性状上的一般配合力相对效应较好;R 189/R 9311在单株有效穗上一般配合力效应最好,R 9311/R 76和R 76/R 9311大部分性状的一般配合力效应无优势。
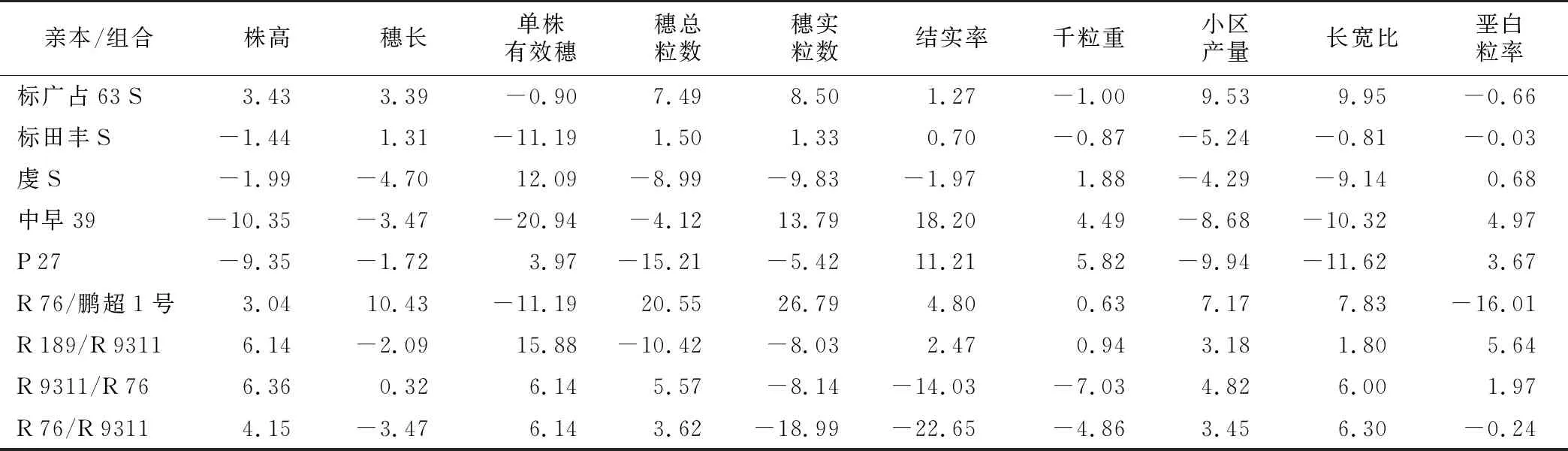
表3 亲本的一般配合力相对效应值
从以上分析可以发现,利用典型晚籼恢复系杂交后,再通过核辐射的方式创制的恢复系对早稻不育系的恢复能力较强,其中R 76/鹏超1号在多数性状上均具有较大的一般配合力,甚至超越了配合力较强、应用广泛的恢复系中早39,利用该恢复系容易选育出穗大粒多、结实率高、产量好、垩白粒率低的组合。
从表4可以看出,株高、穗长、单株有效穗、穗总粒数、穗实粒数、结实率、千粒重、小区产量、长宽比、垩白粒率的特殊配合力最高的组合分别是标田丰S/中早39、标田丰S/(R 76/鹏超1号)、虔S/(R 9311/R 76)和虔S/(R 76/R 9311)、标广占63 S/(R 76/鹏超1号)、标广占63 S/(R 76/鹏超1号)、虔S/中早39、虔S/中早39、标广占63 S/(R 9311/R 76)、标广占63 S/(R 76/鹏超1号)和标广占63 S/(R 76/鹏超1号),特殊配合力最低的组合分别是标广占63 S/(R 189/R 9311)、虔S/(R 189/R 9311)、虔S/中早39、虔S/(R 189/R 9311)、虔S/(R 76/R 9311)、虔S/(R 76/R 9311)、标田丰S/(R 9311/R 76)、标田丰S/P 27、虔S/P 27和虔S/P 27。从结果看,在各性状配合力最高的组合中,用到最多的不育系为标广占63 S,其次为虔S,标田丰S最少。使用最多的恢复系为R 76/鹏超1号,其次为中早39。在各性状配合力最低的组合中,使用最多的不育系为虔S,其次为标田丰S,标广占63 S用得最少。使用最多的恢复系为P 27和R 189/R 9311,其次为R 76/R 9311。在同一性状不同不育系组合间特殊配合力表现不同,同一不育系组合不同性状间特殊配合力效应也不相同,说明基因互作具有多样性。对每个组合而言,虔S/中早39的株高、穗实粒数、结实率和千粒重的特殊配合力较高,但单株有效穗、小区产量和长宽比的特殊配合力又很低。虔S/(R 76/R 9311)穗实粒数和结实率的特殊配合力效应值均较低,但单株有效穗的特殊配合力效应值最高。另外,标广占63 S和R 76/鹏超1号分别是一般配合力表现最好的不育系和恢复系,但标广占63 S/(R 76/鹏超1号)仅穗总粒数、穗实粒数、长宽比和垩白粒率的特殊配合力表现最佳,其余性状的特殊配合力仅为中上等水平。标田丰S和P 27的一般配合力表现一般,但标田丰S/P 27株高、结实率和千粒重的特殊配合力均较高。由此可见,一般配合力与特殊配合力并无一一对应关系。
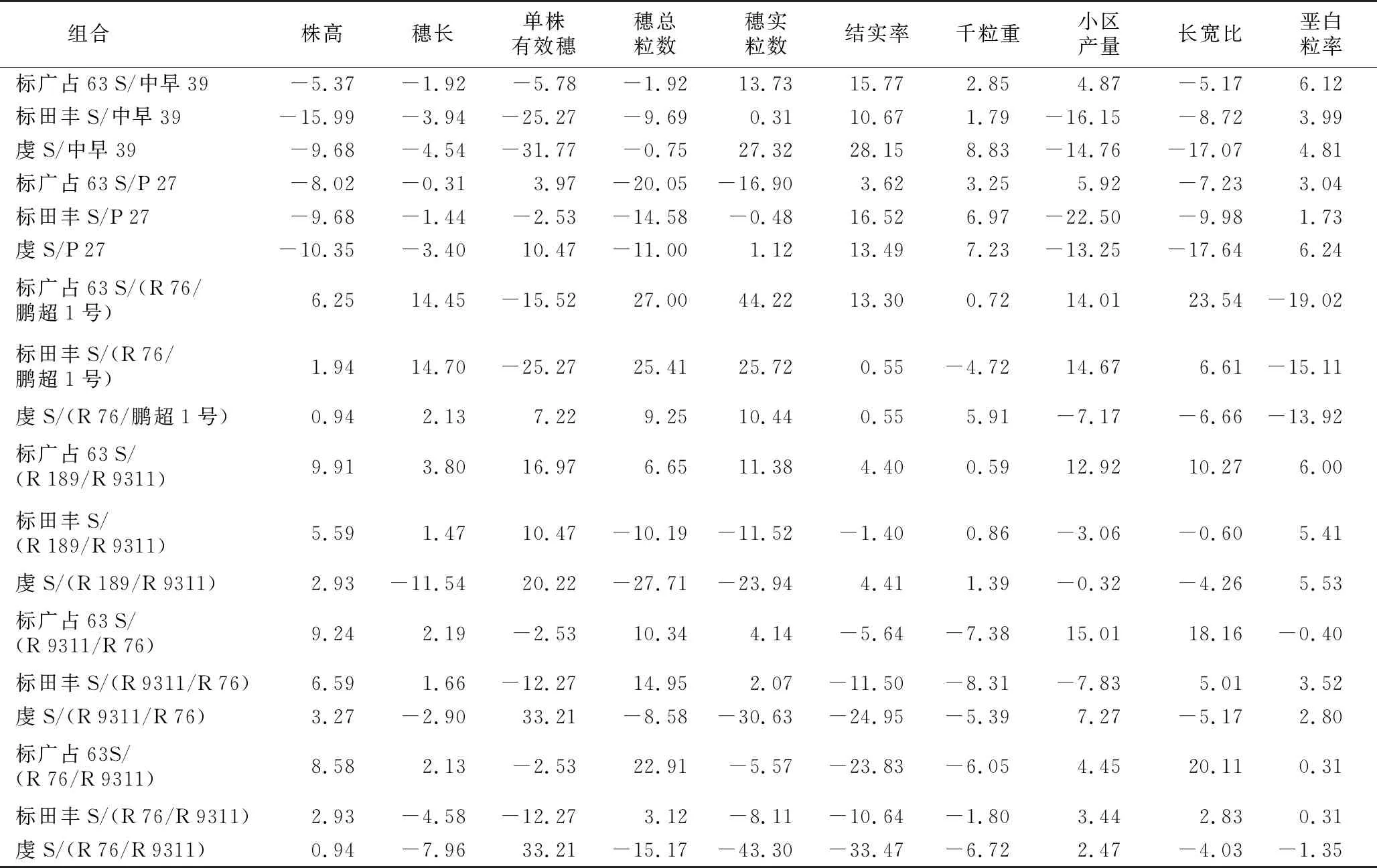
表4 各组合性状特殊配合力相对效应值
从一般配合力方差Vg(%)和特殊配合力方差Vs(%)分别占配合力总方差的比率看,10个主要农艺性状的一般配合力方差占较大比例(表5),说明这些性状以基因加性作用为主,在这些性状上对亲本的选择是主要的。
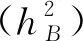
表5 配合力方差分量及遗传力估值
3 小结与讨论
3.1 核辐射诱变育种与聚合育种在早稻种质创新中的应用价值
核辐射诱变技术是作物育种中重要的技术之一,由于核辐射诱变育种的生物安全性高和独特的“创造性”,对解决目前水稻育种中亲本材料过于集中和单一、遗传多样性降低等问题具有重要的作用[6]。本研究采用亲本材料杂交后再进行核辐射处理,其辐射后代的变异范围扩大,变化更为丰富,提高了选择优良基因型的频率。
目前,聚合育种在水稻抗病虫性和品质上运用较多,本研究的目的是采用核辐射诱变与聚合育种相结合的方法,将晚稻材料改良为相对早熟的早稻材料,进一步拓宽早稻育种的亲本遗传基础。本研究选用的亲本材料广占63 S、R 76、鹏超1号、R 189和R 9311都是中晚稻类型,采用先杂交后核辐射处理的方法聚合水稻优良基因,选育出的4个恢复系与标广占63 S、标田丰S、虔S杂交配制的12个两系杂交早稻组合中,有11个组合播始历期比对照早丰优402(播始历期79 d)短,1个组合比对照中早35短(播始历期76 d),1个组合与中早35相当(表6),早熟特性明显。研究表明,随着水稻品种生育期的缩短,其生物学产量和经济产量并不一定降低[7-9]。创制的新质源不育系标广占63 S和恢复系(R 76/鹏超1号)在产量相关性状和米质上都有很好的表现,具有较好的应用价值。
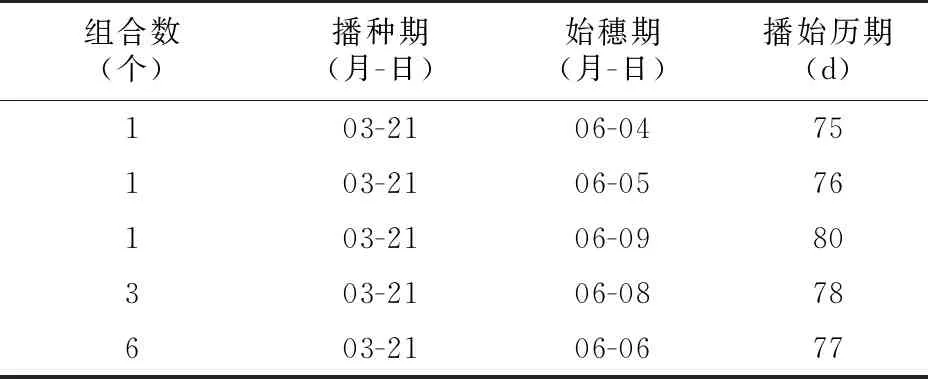
表6 12个杂交组合播始历期
3.2 亲本配合力与杂种优势的关系
杂交水稻主要是利用双亲间的杂种优势,将性状优良的亲本通过合理的组配,使其优良性状在后代中得到体现和加强,从而达到高产、优质和多抗的目的。在育种实践过程中选育高配合力的亲本极其重要,杂交水稻配合力的研究是个复杂的问题,国内外许多研究表明,配合力是可以遗传的,一般配合力和特殊配合力并非一一对应的关系,一般配合力高的亲本,其组合的特殊配合力并不一定高,杂种优势不一定强[10-11]。只有亲本或亲本之一的一般配合力高,组合特殊配合力也高的杂交组合才具有较高的增产潜力[12-13]。一般配合力高,特殊配合力方差大的亲本较容易选育出强优势组合。一般配合力高,特殊配合力方差小的亲本能将其优良性状整齐地遗传给后代,是最好的亲本类型[14-15]。
本研究结果表明,杂交水稻的农艺性状和产量的表现同时受不育系和恢复系的一般配合力和特殊配合力的共同作用,但不同性状不育系和恢复系的作用不同。只有双亲或双亲之一的一般配合力高,其组合的特殊配合力高时才能有高产或超高产的潜力,如标广占63 S/(R 76/鹏超1号)、标田丰S/(R 76/鹏超1号)等。如用双亲一般配合力低的亲本配组,其组合的特殊配合力也较低,如虔S/中早39、虔S/P 27等,其增产潜力较弱。
3.3 亲本配合力测定的应用前景
配合力测定研究是为了评价亲本,预测亲本的应用前景,只有亲本自身的一般配合力较高才有可能组配出特殊配合力高的杂交组合,才有广阔的应用前景[16-17]。在本研究考察的10个性状中,标广占63 S的穗长、穗总粒数等7个性状的一般配合力较高;标田丰S的株高、穗长等6个性状的一般配合力较高;虔S仅在株高、单株有效穗和千粒重上的一般配合力较高。恢复系(R 76/鹏超1号)在穗长、穗总粒数等7个性状上的一般配合力较高,而(R 9311/R 76)和(R 76/R 9311)多数性状的一般配合力较低,有待进一步选择优良单株配组观察。
综上所述,创制的新质源不育系标广占63 S和恢复系(R 76/鹏超1号)的一般配合力较高,其所配制的杂交组合的特殊配合力也较高,是较理想的亲本材料。这也说明核辐射诱变育种与聚合育种相结合的方法是水稻种质资源创新的重要途径。
