马铃薯叶片内生细菌的分离鉴定及其对马铃薯块茎蛾的毒力研究
2022-03-18杨金睿李俊逸祝春月王文倩肖关丽
杨金睿,吉 泽,李俊逸,姚 遥,祝春月,王文倩,肖关丽*
(1云南农业大学农学与生物技术学院,云南 昆明 650201;2云南农业大学植物保护学院,云南 昆明 650201)
0 引言
【研究意义】马铃薯是我国第四大粮食作物,除自然资源条件外,虫害是制约我国马铃薯单产进一步提升的重要因素(徐进等,2019)。马铃薯块茎蛾(Phthorimaea operculella)是一种世界性农业害虫,对马铃薯植株和块茎均有危害,严重影响马铃薯的产量和品质,已在我国南方马铃薯主产区大范围发生,尤其是在云南等地发生严重(闫俊杰等,2019)。在我国大力推进绿色农业发展的今天,为促进马铃薯产业的可持续发展,采取以微生物等为基础的绿色生物防治技术是马铃薯块茎蛾防控的重要举措。植物内生细菌作为一种新型微生物资源,其丰富的微生物种类为寻找新型抗虫微生物及其活性物质提供了可能。因此,从不同马铃薯品种中探寻对马铃薯块茎蛾有防治作用的内生细菌具有重要的研究开发价值。【前人研究进展】植物内生菌(Endophyte)是指生长在植物各种不同组织内部但又不会引起植物表观症状的微生物(Sturz et al.,2000),主要包括内生细菌、放线菌以及真菌3大类,普遍存在于不同的植物中(张鑫,2018),在植物不同组织及其器官内广泛分布(贾颜等,2021)。当前文献报道的马铃薯内生细菌研究大多集中在马铃薯块茎方面,研究人员通过传统分离培养和高通量测序发现的马铃薯内生细菌主要包括假单胞菌属(Pseudomonas)、微球菌属(Micrococcus)、芽孢杆菌属(Bacillus)、红球菌属(Rhodococcus)、肠杆菌属(Enterobacter)和鞘氨醇单胞菌属(Sphingomonas)等(屈青松等,2016;戴超,2017;Liu et al.,2020;Shi et al.,2021)。在马铃薯内生细菌功能的研究中,研究人员通过内生细菌菌悬液浸泡马铃薯种薯块茎后,根据其生长状况筛选出对马铃薯植株生长发育或致病产生影响的菌株(Istifadah et al.,2018);也有采用平板培养法发现对马铃薯病原真菌有高抑制率的芽孢杆菌属内生细菌——萎缩芽胞杆菌(B.atrophaeus)和贝莱斯芽胞杆菌(B.velezensis)(陈星伊等,2020),而关于内生细菌防治马铃薯块茎蛾的研究则鲜有报道。微生物防治已成为近年来绿色防控马铃薯块茎蛾的研究热点,如苏云金杆菌(Bacillus thuringiensis)已被应用于防治马铃薯块茎蛾,B.thuringiensisssp.Kurstaki(Btk)是一株商用苏云金杆菌菌株,可制成可湿性粉剂作为化学杀虫剂的替代品,在马铃薯收获前施用以抵御马铃薯块茎蛾幼虫在田间对块茎的侵害,降低储藏过程中块茎被侵染的风险(Arthurs et al.,2008);以浸虫法测定球孢白僵菌(Beauveria bassiana)菌株SD对马铃薯块茎蛾1龄幼虫的毒力作用,10 d后致死率高达98%,处理后的幼虫产卵量、生长发育期和后代的净繁殖率均显著降低,对马铃薯块茎蛾造成潜在的长期影响(Yuan et al.,2017);采用金龟子绿僵菌(Metarhizium anisopliae)、莱氏野村菌(Nomuraea rileyi)和细脚拟青霉(Paecilomyces tenuipes)的孢子悬浮液浸泡处理马铃薯叶片和块茎,对取食的马铃薯块茎蛾幼虫均有杀虫作用,还可降低成虫羽化率和卵孵化率,其中莱氏野村菌对防治初孵幼虫侵入马铃薯块茎的效果最好(Khorrami et al.,2018);通过浸叶法测定发现黏质沙雷氏菌(Serratia marcescens)菌株ML-1不同成分对马铃薯块茎蛾幼虫均有较强的杀虫活性(苏造堂,2020)。【本研究切入点】目前用来防治马铃薯块茎蛾的生防菌较少,对马铃薯内生细菌的开发利用仍处于早期阶段,且尚未有通过马铃薯可培养内生细菌防治马铃薯块茎蛾的研究报道。【拟解决的关键问题】以云南省2种马铃薯主栽品种丽薯6号和会-2为材料,采用LB培养基分离培养马铃薯叶片内生细菌,根据内生细菌的形态特征和16S rDNA序列进行种类鉴定;通过浸叶饲喂法测定内生细菌的发酵液、发酵上清液和菌悬液对马铃薯块茎蛾的杀虫活性,以期为马铃薯块茎蛾的生物防治提供菌种资源和理论依据。
1 材料与方法
1.1 试验材料
供试马铃薯品种:丽薯6号和会-2,由云南省曲靖市会泽县农技中心提供。供试虫源:云南农业大学植物保护学院昆虫学实验室饲养繁殖的马铃薯块茎蛾稳定种群,待马铃薯块茎蛾幼虫羽化为成虫后用直形管收集雌、雄成虫并在封口纱布上沾10%蜂蜜水喂食后使其产卵,待纱布上的幼虫孵化后用合作88马铃薯块茎饲养至3龄,选择大小一致、健康和具有活力的马铃薯块茎蛾3龄幼虫作为试验虫源。
培养基:LB固体培养基(NaCl 10 g、胰蛋白胨10 g、酵母浸粉5 g、琼脂15 g,蒸馏水1000 mL,pH 7.0~7.2);LB液体培养基(NaCl 10 g、胰蛋白胨10 g、酵母浸粉5 g、蒸馏水1000 mL,pH 7.0~7.2)。
主要试剂及仪器:3% NaClO溶液、吐温-80、ddH2O、细菌通用引物、2×TaqPCR Master Mix、绿色荧光核酸染料。Mastercycler®Nexus型PCR仪和5430R型多用途高效离心机(德国Eppendorf公司)、DYY-2C型电泳仪(北京六一生物科技有限公司)、RXZ-260B-30恒温光照培养箱(宁波赛福实验仪器有限公司)。
1.2 样品采集与处理
2个马铃薯品种种薯为根据GB 18133—2012《马铃薯种薯》划分为一级种的种薯,重量50~60 g,带有2~3个芽眼,芽长1.0 cm左右。将2个品种的种薯栽种于花盆(直径40 cm,高40 cm),用防虫网罩住以防止害虫危害。以含沙量高、通气性好的沙质土为栽培土壤,2个品种采用相同的种植管理方法,每个品种种植10盆,试验在大棚内进行。马铃薯栽种50 d其植株生长至块茎形成期时,2个品种均选择长势相同的植株中部展开复叶上的完整、健康、无病虫害症状的叶片为试验材料,5次重复,取样后即装入密封袋带回实验室分离内生细菌。
先将2个品种采集的新鲜叶片用流水冲洗1~2 min后用无菌滤纸擦干表面,用无菌水浸泡漂洗3次,放入75%酒精浸泡1 min,再用无菌水将酒精漂洗干净,然后把叶片放入3% NaClO溶液充分浸泡1 min后用无菌水将其漂洗,最后75%酒精浸泡30 s后用无菌水充分漂洗(杨瑞先等,2005)。取少量最后一次漂洗叶片的无菌水涂布在LB固体培养基作为对照,若对照培养基未长出菌落则证明叶片表面消毒彻底。
1.3 内生细菌的分离鉴定
1.3.1 内生细菌的分离 采用研磨法分离内生细菌。取1 g已表面消毒好的新鲜叶片用无菌剪刀剪碎,加入适量无菌水在无菌研钵中研磨成匀浆状溶液,取100 μL在LB固体培养基涂布培养,设5次重复(张海龙,2014)。
1.3.2 细菌种类鉴定
1.3.2.1 细菌形态学鉴定 将叶片中分离到的细菌菌株在LB固体培养基上纯化培养,对纯化好的菌株参照《伯杰细菌鉴定手册》(Buchanan and Gibbons,1999)对其菌落颜色、形态等培养性状进行初步鉴定。采用革兰氏染色法鉴定细菌显色反应确定革兰氏阴阳性,并观察其细胞形态。
1.3.2.2 细菌分子鉴定 以冻溶法提取细菌菌株DNA(郑亚强等,2017)。将LB固体培养基上纯化培养好的细菌用无菌接种环挑取少许单菌落放入装有500 μL无菌水的无菌离心管中,震荡仪振荡使离心管中的菌体和无菌水充分混匀,然后将装有菌液的离心管在液氮中冰冻10 min后取出,立即放置在漂浮盘上于沸水中水浴5 min,取出后放入离心机在12000 r/min条件下离心2 min,离心管中的上清液即为DNA模板。以细菌通用引物27F(5'-AGAGTTT GATCCTGGCTCAG-3')和1492R(5'-TACGACTTA ACCCCAATCGC-3')扩增细菌16S rDNA基因,PCR反应体系25.0 μL:上、下游引物各1.0 μL,PCR Master Mix 12.5 µL,DNA模板1.0 µL,ddH2O 9.5 μL。扩增程序:94 ℃预变性5 min;94 ℃1 min,53 ℃1 min,72 ℃2 min,进行32个循环;72 ℃延伸10 min。取3 μL PCR产物采用凝胶电泳检测,合格的PCR产物送往生工生物工程(上海)股份有限公司测序。
1.4 内生细菌对马铃薯块茎蛾的杀虫活性测定
1.4.1 内生细菌发酵液杀虫活性测定 以分离获得的内生细菌为供试菌株,将已在LB固体培养基上纯化好培养48 h的供试菌株分别接种于装有100 mL无菌LB液体培养基的锥形瓶中,置于摇床28 ℃下180 r/min培养24 h。参考杨建云等(2014)的方法测定细菌菌悬液浓度,使用分光光度计时首先以LB液体培养基调零,然后测定培养24 h时各原始细菌发酵液的OD600,稀释1×106~1×108倍后取100 μL涂布于LB液体培养基上培养,待细菌菌落长出后根据菌落数测定原始细菌发酵液浓度,以无菌LB液体培养基为空白对照,每处理3次重复。依据上述方法,测定培养24 h原始发酵液中细菌菌体浓度后再稀释到1.5×105~1.5×109CFU/mL用于试验。
采用浸叶饲喂法(苏造堂等,2020)饲喂马铃薯块茎蛾3龄幼虫。先将马铃薯植株复叶叶片在配置成不同浓度的细菌发酵液中分别浸渍30 s,将叶片取出自然风干后放置在灭菌培养皿中(培养皿直径为15 cm,垫1层吸水纸),并在叶片茎秆尾部包裹湿润脱脂棉以保持叶片水分,挑取20头供试马铃薯块茎蛾3龄幼虫置于叶片表面,保鲜膜封口,并扎一定数量的小孔透气,在恒温光照培养箱中28 ℃、湿度75%条件下培养。每株菌株发酵液不同浓度处理均设3个重复,以无菌LB液体培养基浸泡叶片为对照(CK)。
1.4.2 马铃薯叶片内生细菌发酵上清液杀虫活性测定 细菌发酵液的制备同1.4.1,将培养好的供试菌株发酵液配置为菌体浓度1.5×109CFU/mL,将发酵液在10000 r/min条件下离心10 min,最后用无菌注射器将上清液吸出使其通过孔径大小为0.22 μm无菌滤膜,以获得无菌发酵上清液(许守涛等,2018),采用浸叶饲喂法测定其对马铃薯块茎蛾3龄幼虫的毒力。
1.4.3 马铃薯叶片内生细菌菌悬液的杀虫活性测定将供试细菌菌株接种在LB固体培养基上活化培养2 d,用含有0.05%吐温-80的无菌水把培养好的菌落冲洗于无菌三角瓶中,在摇床180 r/min条件下充分振荡均匀(徐超等,2019),采用1.4.1方法测定菌悬液浓度,再将其配置为菌体浓度1.5×109CFU/mL,采用浸叶饲喂法测定其对马铃薯块茎蛾3龄幼虫的毒力。
1.5 数据处理与统计分析
对供试菌株测序结果用Contigexpress软件进行拼接、校对处理,把拼接好的16S rDNA序列提交至NCBI,查找并下载数据库中同源性最高的部分菌株序列。采用MEGA 7.0,运用邻接法(Neighbour-joining)及Kimura’s 2-parameter矫正模型构建菌株系统发育进化树,进行1000次Bootstrap验证。
用Excel 2021进行数据整理,采用DPS version 7.05的Duncan’s新复极差法对数据进行差异显著性分析,采用生物测定模块分析不同处理毒力测定数据,计算毒力回归方程、致死中时间(LT50)和致死中浓度(LC50)。
2 结果与分析
2.1 马铃薯叶片内生细菌分离和鉴定结果
从马铃薯品种丽薯6号叶片中分离到1株内生细菌,编号QL1;在28 ℃光照培养箱条件下经LB固体培养基培养,菌株QL1菌落呈近似圆形,淡白色,不透明,微隆起,边缘规则,表面干燥粗糙,革兰氏染色阳性,菌体为杆状(图1-A和图1-B),与已报道的苏云金芽孢杆菌(B.thuringiensis)相同(胡虓等,2020)。从马铃薯品种会-2叶片中分离到1株内生细菌,编号YH2;在28 ℃光照培养箱条件下经LB固体培养基培养,菌株YH2菌落呈圆形,橙色,不透明,隆起,边缘规则,表面湿润,革兰氏染色阳性,菌体为短杆状(图1-C和图1-D),与已报道的带化红球菌(R.fascians)相同(Radhika et al.,2015)。
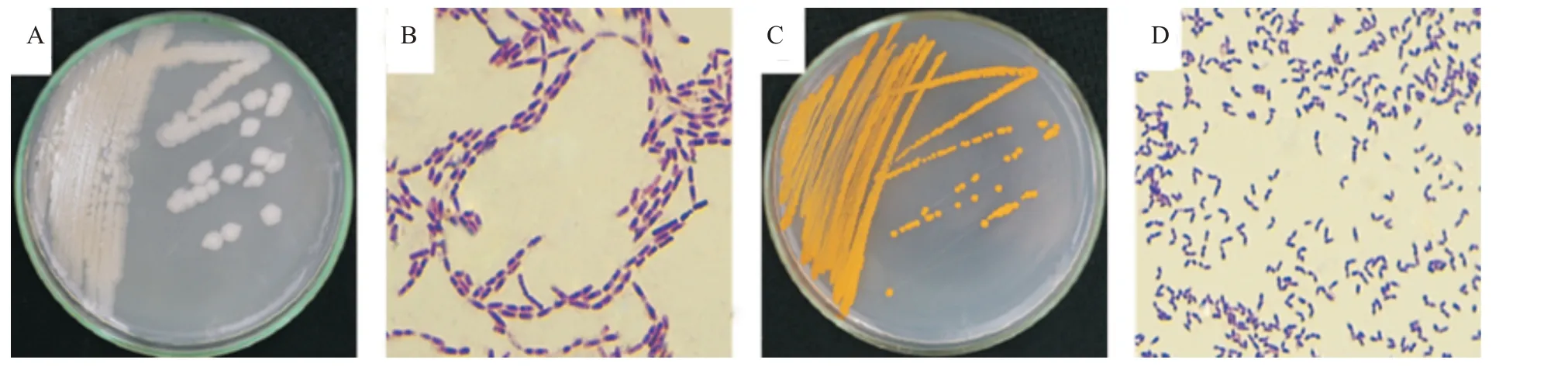
图1 内生细菌菌株QL1和YH2菌落形态及革兰氏染色Fig.1 Colony morphology and Gram staining of endophytic bacteria strains QL1 and YH2
将菌株QL1和YH2测序结果校对拼接后分别提交GenBank数据库进行比对,结果发现,菌株QL1与3株苏云金芽孢杆菌的相似性达99.79%以上,菌株YH2与2株带化红球菌的相似性达99.54%以上。从系统发育进化树(图2)可看出,菌株QL1先与3株苏云金芽孢杆菌聚为一支,且4株菌处在同一个分支长度上,然后再与其他芽孢杆菌属的菌株聚合,表明菌株QL1与苏云金芽孢杆菌的亲缘关系很近,结合形态特征等确定菌株QL1为厚壁菌门芽孢杆菌属的苏云金芽孢杆菌。从系统发育进化树(图3)可看出,菌株YH2先与2株带化红球菌聚为一支,且3株菌处在同一个分支长度上,然后再与其他红球菌属的菌株聚合,表明菌株YH2与带化红球菌的亲缘关系很近,结合形态特征等确定菌株YH2为放线菌门红球菌属的带化红球菌。
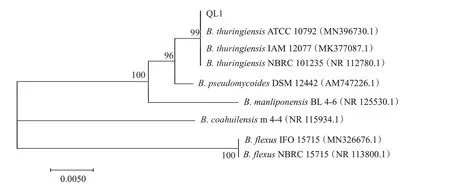
图2 基于16S rDNA序列构建的菌株QL1系统发育进化树Fig.2 Phylogenetic tree of strain QL1 based on 16S rDNA sequence
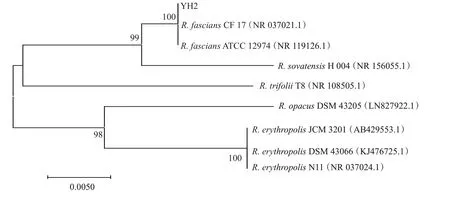
图3 基于16S rDNA序列构建的菌株YH2系统发育进化树Fig.3 Phylogenetic tree of strain YH2 based on 16S rDNA sequences
2.2 马铃薯叶片内生细菌对马铃薯块茎蛾3龄幼虫的杀虫活性
2.2.1 细菌发酵液浓度 2种细菌发酵液在摇床28 ℃,180 r/mim条件下培养24 h后用紫外分光光度计测得菌株QL1的OD600均值为0.97,对应涂布法浓度为1.5×109CFU/mL;菌株YH2的OD600均值为0.98,对应涂布法浓度为1.8×109CFU/mL。
2.2.2 菌株QL1发酵液对马铃薯块茎蛾3龄幼虫的毒力 菌株QL1各浓度发酵液处理后马铃薯块茎蛾3龄幼虫的累积死亡率均随着处理时间延长而逐渐升高;第3 d开始所有浓度处理幼虫的累积死亡率均显著高于对照(P<0.05,下同);处理后第7 d,菌株QL1发酵液浓度1.5×105~1.5×109CFU/mL处理下幼虫的累积死亡率分别为60.00%、66.67%、73.33%、75.00%和81.67%(表1)。

表1 菌株QL1不同浓度发酵液处理后马铃薯块茎蛾3龄幼虫的累积死亡率Table 1 Cumulative mortality rate of P.operculella 3rd instar larvae treated with strain QL1 fermentation broth of different concentrations
运用DPS进行生物测定,结果显示,以浓度为1.5×105~1.5×109CFU/mL的菌株QL1发酵液处理马铃薯叶片,马铃薯块茎蛾取食7 d后的LT50在3.03~5.73 d,浓度最高时LT50最短,表明浓度越高对3龄幼虫的杀虫活性越强,毒力方程相关系数在0.982~0.992,毒力方程拟合程度较好(表2)。各浓度菌株QL1发酵液处理叶片后,马铃薯块茎蛾取食第3、5和7 d的LC50分别为7.99×108、3.21×106和2.73×103CFU/mL,表明菌株QL1对马铃薯块茎蛾3龄幼虫的致死作用表现出明显的剂量与效应关系,随时间延长LC50逐渐降低(表3)。
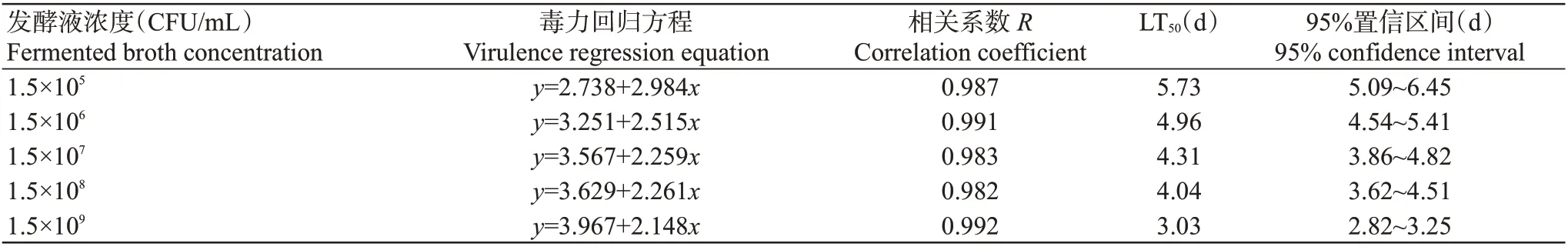
表2 菌株QL1不同浓度发酵液对马铃薯块茎蛾3龄幼虫的毒力(7 d)Table 2 Virulence of different concentrations of strain QL1 fermentation broth to the P.operculella 3rd instar larvae(7 d)
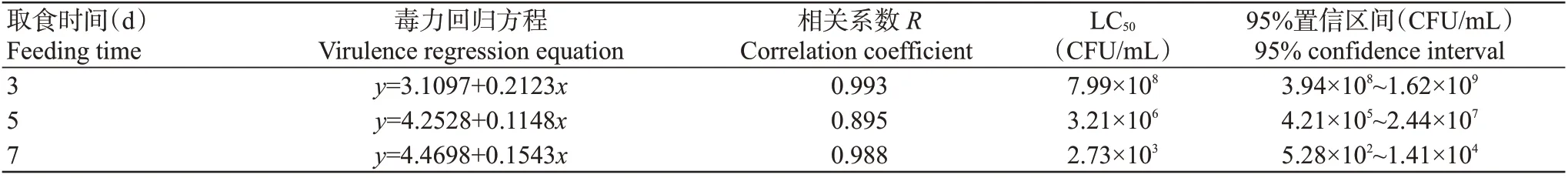
表3 不同处理时间菌株QL1发酵液对马铃薯块茎蛾3龄幼虫的毒力Table 3 Virulence of different treatment times of strain QL1 fermentation broth to the P.operculella 3rd instar larvae
从图4可看出,处理1 d后,菌株QL1发酵液处理马铃薯块茎蛾3龄幼虫的累积死亡率均显著高于发酵上清液和菌悬液处理,而发酵上清液和菌悬液处理间无显著差异(P>0.05,下同);从处理第4 d起,3种处理对马铃薯块茎蛾3龄幼虫的累积死亡率均显著高于对照;在第7 d时,菌株发酵液、发酵上清液和菌悬液处理对马铃薯块茎蛾3龄幼虫的累积死亡率分别达81.67%、50.00%和60.00%。
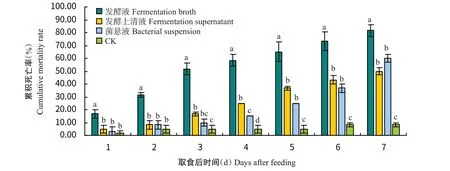
图4 取食菌株QL1不同溶液后马铃薯块茎蛾3龄幼虫的累积死亡率Fig.4 Daily cumulative mortality rate of P.operculella 3rd instar larvae after feeding different solutions of strain QL1
2.2.3 菌株YH2发酵液对马铃薯块茎蛾3龄幼虫的毒力 以浓度为1.5×105~1.5×109CFU/mL的菌株YH2发酵液处理马铃薯叶片,马铃薯块茎蛾3龄幼虫取食后的累积死亡率均随着处理时间延长而逐渐升高,第3 d开始所有浓度处理马铃薯块茎蛾3龄幼虫的累积死亡率均显著高于对照;处理后第7 d,菌株YH2发酵液浓度1.5×105~1.5×109CFU/mL处理造成该幼虫的累积死亡率分别为48.33%、53.33%、55.00%、61.67%和65.00%,除最低浓度发酵液处理外,其他浓度处理后马铃薯块茎蛾3龄幼虫的累积死亡率间无显著差异(表4)。
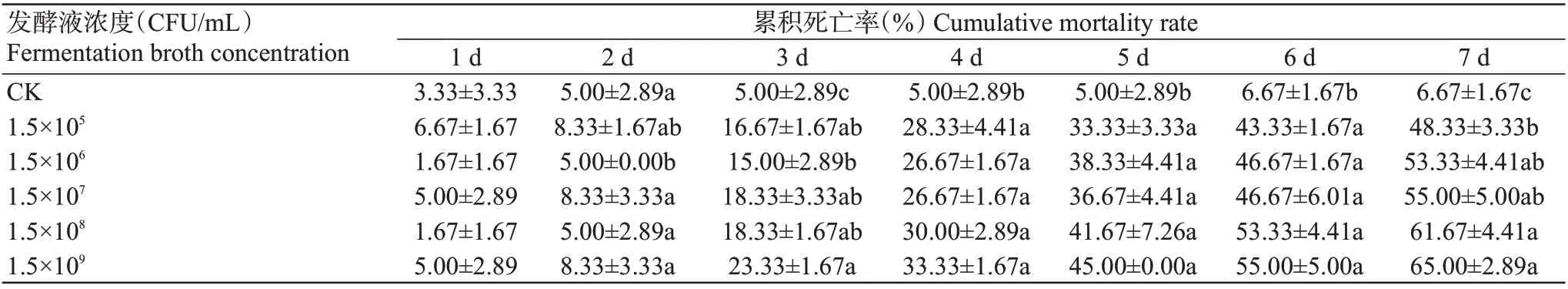
表4 菌株YH2不同浓度发酵液处理后马铃薯块茎蛾3龄幼虫的累积死亡率Table 4 Cumulative mortality rate of P. operculella 3rd instars larvae treated by different concentrations of strain YH2 fermentation broth
运用DPS进行生物测定,由数量型数据机制分析可知,以浓度为1.5×105~1.5×109CFU/mL的菌株YH2发酵液处理马铃叶片,马铃薯块茎蛾取食7 d后LT50在5.58~7.07 d,浓度最高时LT50最短,但不同浓度处理间LT50差距不明显,毒力方程相关系数在0.965~0.991,毒力方程拟合程度较好;1.5×105CFU/mL的发酵液处理因致死率低于50.00%,故无法估算LT50(表5)。此外,菌株YH2菌体无显著致死效应,因此无法以菌体浓度来估算其对马铃薯块茎蛾幼虫的LC50。
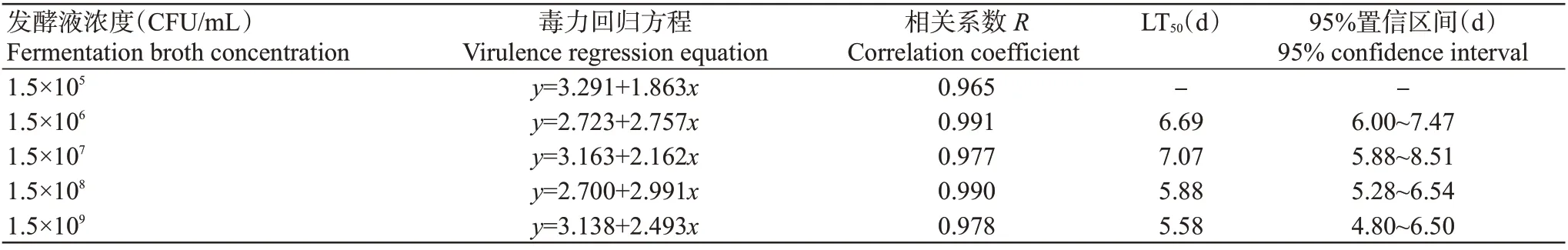
表5 菌株YH2不同浓度发酵液对马铃薯块茎蛾3龄幼虫的毒力(7 d)Table 5 The toxicity of different concentrations of strain YH2 fermentation broth to the P.operculella 3rd instar larvae(7 d)
从图5可看出,在处理第7 d时,菌株YH2发酵液和发酵上清液处理对马铃薯块茎蛾3龄幼虫的累积死亡率间无显著差异,且均显著高于菌悬液和对照处理;从第3 d后,发酵液处理的累积死亡率一直为3种处理中最高,且3种处理对马铃薯块茎蛾3龄幼虫的累积死亡率均显著高于对照处理;在第7 d时,菌株YH2发酵液、发酵上清液和菌悬液处理对马铃薯块茎蛾3龄幼虫的累积死亡率分别达65.00%、55.00%和38.33%。
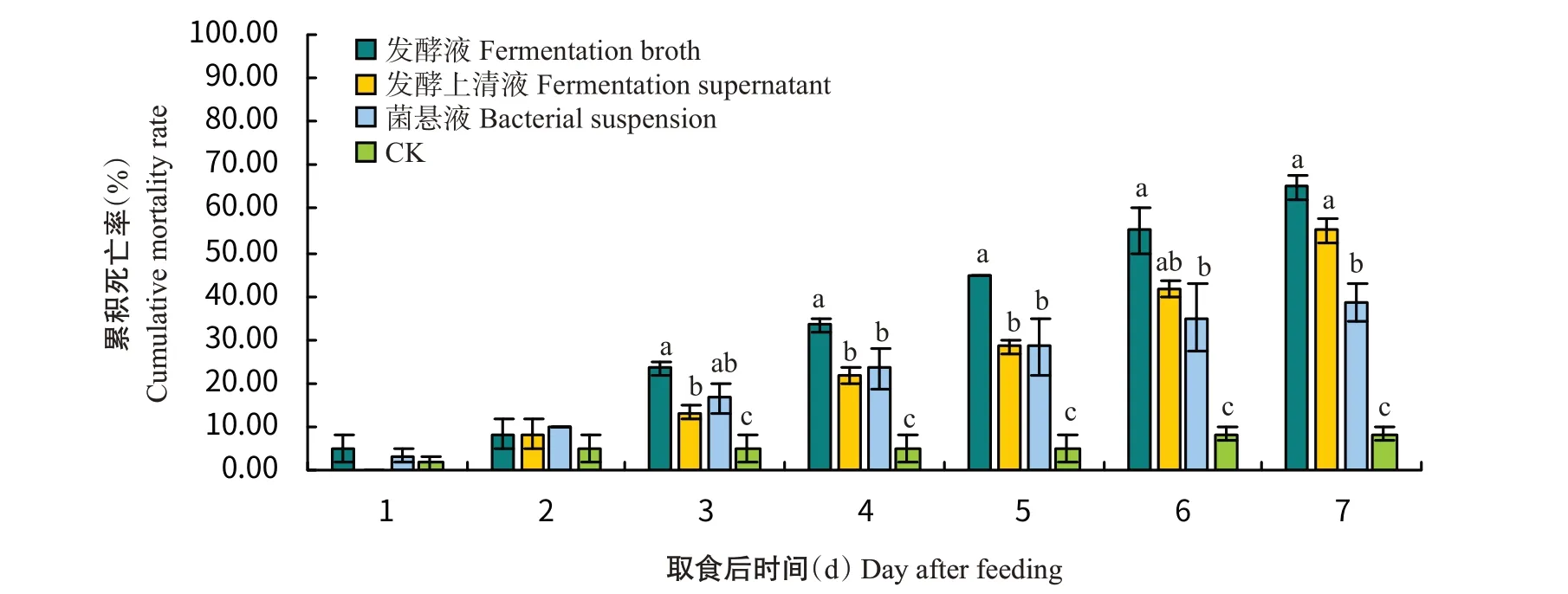
图5 取食菌株YH2不同溶液后马铃薯块茎蛾3龄幼虫的累积死亡率Fig.5 Daily cumulative mortality rate of P.operculella 3rd instar larvae after being fed with different solutions of strain YH2
2.2.4 菌株QL1和YH2不同溶液对马铃薯块茎蛾3龄幼虫的毒力差异 比较供试菌株QL1和YH2不同溶液处理马铃薯块茎蛾3龄幼虫7 d后的毒力差异,结果(表6)显示,菌株YH2发酵液处理后幼虫的累积死亡率显著低于菌株QL1发酵液处理;菌株QL1和YH2发酵上清液处理后其幼虫的累积死亡率间无显著差异;菌株YH2菌悬液处理后幼虫的累积死亡率显著低于菌株QL1菌悬液处理,且累积死亡率低于50.00%,无法估算其LT50。
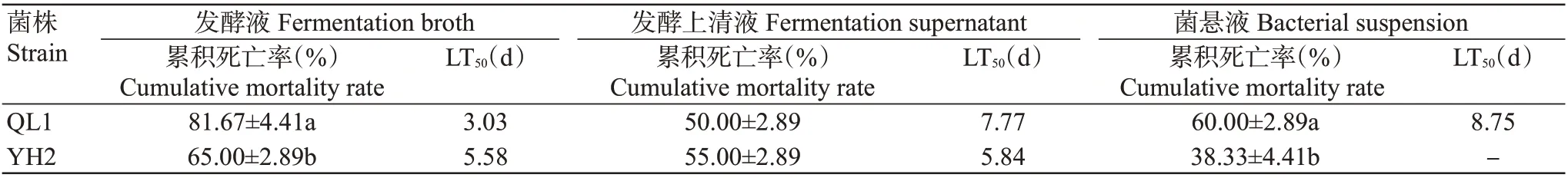
表6 菌株QL1与YH2不同溶液对马铃薯块茎蛾3龄幼虫累积死亡率的影响Table 6 Effects of different solutions of strains QL1 and YH2 on cumulative mortality rate of P.operculella 3rd instar larvae
3 讨论
本研究从云南主栽马铃薯品种丽薯6号和会-2叶片中分别分离到对马铃薯块茎蛾有杀虫活性的内生细菌芽孢杆菌属的苏云金芽孢杆菌QL1和红球菌属的带化红球菌YH2。当前对马铃薯叶片内生细菌的研究较少,在文献报道的马铃薯块茎内生细菌研究中发现了与本研究供试菌株相同菌属的细菌(戴超,2017;Liu et al.,2020),但未发现同一个种的内生细菌,表明在不同马铃薯植株部位和品种,不同马铃薯种植地区和生长环境等条件下可能存在同菌属的内生细菌,但因这些条件不同也使各马铃薯品种存在其独有的内生细菌种类。此外,在研究报道分离的马铃薯内生细菌中,其功能主要有促进和抑制马铃薯生长、使植株致病或抑制病原菌的作用(Pageni et al.,2014;Istifadah et al.,2018;陈星伊等,2020),尚未发现对马铃薯块茎蛾有杀虫活性的内生细菌。
本研究通过室内生物测定发现,在1.5×109CFU/mL浓度下,菌株QL1和YH2发酵液处理7 d后马铃薯块茎蛾3龄幼虫的累积死亡率可达81.67%和65.00%,表明2种供试菌株对该幼虫有较强的致死率。研究表明,菌株QL1和YH2对马铃薯块茎蛾3龄幼虫的毒杀作用有明显差异的同时也存在相似之处:2种菌株的不同溶液对马铃薯块茎蛾3龄幼虫均有杀虫活性,且均表现为发酵液杀虫活性更强,菌悬液及发酵上清液单独使用对幼虫的毒杀效果较发酵液低,该结果与粘质沙雷氏菌对马铃薯块茎蛾的杀虫活性特点相似(苏造堂等,2020),这可能是菌体在液体培养基中代谢产生了具有杀虫活性的次生代谢产物被充分溶解,幼虫取食后导致死亡率升高,但其具体原因还有待深入研究验证。此外,在处理第7 d时,菌株QL1菌悬液对马铃薯块茎蛾3龄幼虫的毒杀效果高于发酵上清液处理,而菌株YH2则相反,其原因可能与幼虫取食2种菌株后对其肠道微生物产生的影响有关,因为昆虫肠道菌群的变化可对宿主的生长代谢造成直接影响,甚至可导致疾病等(魏博帆等,2021)
苏云金芽孢杆菌是一种传统的杀虫细菌,已开发为害虫生防产品用于多种农林害虫和卫生害虫的生物防治,此类细菌也被应用于防治马铃薯块茎蛾(杜霞等,2021),可在马铃薯收获前施用以抵御马铃薯块茎蛾幼虫在田间对块茎的侵害,已成为印度、秘鲁和埃及等国家防治马铃薯块茎蛾的重要药剂(Aryal and Jung,2015)。本研究从马铃薯叶片中分离获得的苏云金芽孢杆菌QL1,其不同溶液对马铃薯块茎蛾3龄幼虫均具有较强的杀虫活性。红球菌属的细菌种类丰富、分布广泛、代谢功能多样,具有的酶种类丰富,可降解许多合成或天然的有机化合物(吴金芳等,2022),但鲜有对防治虫害方面的研究报道。带化红球菌作为植物内生菌时可释放多种细胞分裂素对寄主植物形态造成显著影响,还可引起寄主植物一系列的疾病症状(Jameson,2019)。而本研究分离到的带化红球菌YH2发酵液和发酵上清液饲喂马铃薯块茎蛾3龄幼虫后,其致死率均可达50.00%以上。
4 结论
从云南2种马铃薯主栽品种丽薯6号和会-2叶片中分别分离到内生细菌菌株QL1和YH2,2种菌株配制的3种溶液(发酵液、发酵上清液和菌悬液)对马铃薯块茎蛾3龄幼虫均有较强的杀虫活性,可作为马铃薯块茎蛾生防制剂开发的潜力菌株。
