红鳍笛鲷视杆蛋白和长波敏感视蛋白的基因克隆及表达规律分析
2022-03-18陈子钊刘雪媚许振民梁秋璐王中铎郭昱嵩
陈子钊,刘雪媚,许振民,梁秋璐,王中铎*,郭昱嵩,2*
(1广东海洋大学水产学院/南海水产经济动物增养殖广东普通高校重点实验室,广东 湛江 524088;2广东省水产动物病害防控与健康养殖重点实验室,广东 湛江 524088)
0 引言
【研究意义】视蛋白(Opsin)是一类约含350个氨基酸残基,含有7个跨膜结构域,隶属于G蛋白偶联受体超家族(G protein-coupled receptors,GPCR)的成员(Bowmaker and Hunt,2006)。根据是否参与视觉成像,可将视蛋白分为视觉视蛋白和非视觉视蛋白两大类。视杆蛋白(Rhodopsin,RH1)和长波敏感视蛋白(Long-wave sensitive opsin,LWS)作为直接参与视觉成像的2种重要视觉视蛋白,不仅对外界不断变化的光环境视觉和信号系统适应方面发挥重要作用,而且在物种的进化上也具有重要影响(Cortesi et al.,2015)。相关研究表明,鱼类在不同发育阶段的视蛋白表达种类及生物表达量是适应光谱环境变化的自身调节机制,如黄盖鲽(Pseudopleuronectes yokohamae)在生长发育中,RH1基因和LWS基因表达量逐渐上升,SWS1和SWS2b等短波敏感视蛋白基因则逐渐下降(Sato et al.,2021)。此外,鳜鱼(Siniperca chuatsi)、斑马鱼(Danio rerio)等不同鱼类也具有视蛋白基因,且随生长发育其表达量呈动态变化的规律(Takechi,2005;Tang et al.,2022)。红鳍笛鲷(Lutjanus erythropterus)是一种广泛分布于热带和亚热带海域及岩礁地区的暖水性近底层鱼类,是我国南部沿海地区的重要养殖对象,具有很高的经济价值。红鳍笛鲷早期发育过程中历经几次重要的变态发育,从早期的浮游生活到后期的底栖生活,其不同栖息水层的光照环境变化大,视蛋白基因在驱动红鳍笛鲷变态发育过程中起作用。因此,克隆红鳍笛鲷RH1基因和LWS基因,并分析其在早期发育阶段及成鱼不同组织的表达规律,对探究笛鲷属鱼类视觉形成在早期发育及环境适应中的变化机制具有重要意义。【前人研究进展】视觉是动物获取外界信息的重要途径,在觅食、求偶及躲避敌害等方面具有重要作用(Liang et al.,2022)。动物的视觉形成与视网膜上的视觉色素形成有关。Wald(1968)发现了视觉色素的分子结构,组成单位为一个生色基团和一个视蛋白,同时提出视觉色素的光敏感性变化规律和对不同波长的最大吸收峰(λmax)是由这两者的相互作用而决定。根据序列之间的相似性和λmax不同,可将视蛋白基因划分为5种亚型,分别为视锥蛋白中的长波敏感视蛋白(Long-wave or middle-wave sensitive opsin,LWS/MWS)、紫外敏感视蛋白和蓝色敏感视蛋白(Shortwave sensitive opsin/SWS1-like opsin,SWS1/SWS2)、绿色敏感视蛋白(Rhodopsin-like opsin)及视杆蛋白中的RH1(Rhodopsin)(刘楚吾等,2015)。国外学者对视蛋白的研究早于国内,Nathans 和Hogness(1983)于1983年在牛眼睛上克隆获得第一个视杆蛋白基因;Kim等(2019)则通过实时荧光定量PCR和石蜡切片探究赤点石斑鱼(Epinephelus akaara)幼体发育过程中视蛋白基因的表达和定位情况,结果发现MWS、LWS和RH1基因在视网膜中高度表达。此外,Kasagi等(2015)研究了日本比目鱼(Veraspermoseri)在发育过程及栖息水域的变化过程中相关视蛋白基因的表达情况,结果发现随着日本比目鱼生长发育,RH1基因的表达量逐渐增高,LWS、SWS2b和SWS1等视蛋白基因则逐渐下降。国内学者对视蛋白基因的研究,随着高通量测序技术的发展,已由模式生物逐渐转移到非模式生物上。Liu等(2020)通过基因组和转录组测序技术鉴定了牙鲆(Paralichthys olivaceus)32个视蛋白基因,包括10个视觉视蛋白和22个非视觉视蛋白,并对非视觉视蛋白进行表达谱分析。【本研究切入点】目前对红鳍笛鲷早期发育阶段的划分及形态数据已有较多报道(刘皓等,2015;陈子钊等,2022),但与此过程密切相关的视蛋白基因表达变化尚未见报道,在红鳍笛鲷生长发育中的变化机制也尚未明确。【拟解决的关键问题】利用RACE克隆红鳍笛鲷2个视蛋白基因RH1和LWS的cDNA全长,利用实时荧光定量PCR检测其在不同发育时期和不同组织的表达规律,明确红鳍笛鲷早期视蛋白基因随光谱变化的表达特征,为揭示笛鲷属鱼类视觉形成在早期发育及环境适应中的变化机制打下理论基础。
1 材料与方法
1.1 试验材料
红鳍笛鲷早期发育阶段样品取自三亚市陵水县蓝海洋水产有限公司,包括胚胎发育阶段的原肠下包1/2期(1/2)、胚孔封闭期(bc)、视囊期(oc)、晶体出现期(eIf)、心脏跳动期(hb)和孵化出膜期(hat),每管取样30~40枚受精卵,以及孵化出膜后1、3、10、15和20 d共5个时期的仔鱼,样品保存于液氮中。试验成鱼3尾均购自广东省湛江市霞山水产品批发市场,体重497±23 g,体长29±2 cm,于自然光下暂养2 h,暂养水温(26±2)℃,采用适量MS-222(25 mg/L)将试验成鱼麻醉后,用高温灭菌过无RNA酶的解剖剪快速解剖,取脑、心脏、肝脏、肌肉、黑色皮肤、红色皮肤、胃和视网膜等8个组织样品放于液氮速冻后,置于-80 ℃冰箱保存备用。
1.2 主要试剂
MS-222购自Sigma公司;TRIzol Reagent RNA购自Invitrogen公司;TransScript Uni All-in-One First-Strand cDNA Synthesis购自北京全式金生物技术有限公司;SYBR®Green qPCR Mix 购自广州东盛生物科技有限公司;RACE试剂盒购自Clontech公司;pMD18-T载体和JM109感受态细胞购自TaKaRa公司。
1.3 总RNA提取及cDNA第一链合成
采用TRIzol法提取总RNA,全程冰上操作,经NanoDrop 2000核酸定量仪检测RNA浓度及A260/A280比值,并用1.0%琼脂糖凝胶电泳检测RNA质量后,采用反转录试剂盒合成cDNA第一链。
1.4 RH1和LWS基因cDNA全长序列克隆
从南海水产经济动物增养殖重点实验室构建的转录组数据库中获取RH1和LWS2个基因的EST序列,采用Primer Premier 5.0设计特异性扩增引物(表1)。以视网膜cDNA为模板,对红鳍笛鲷RH1和LWS基因中间片段进行扩增,并按照SMARTerTMRACE cDNA Amplification Kit说明进行5'-RACE和3'-RACE。采用1%琼脂糖凝胶电泳对扩增产物进行检测,并切胶回收和纯化目的条带,然后将其与pMD18-T载体连接,然后转化至大肠杆菌JM109感受态细胞,于37 ℃恒温箱中倒置振荡培养过夜,菌液PCR鉴定后挑取阳性克隆送至生工生物工程(上海)股份有限公司测序。
1.5 生物信息学分析
通过DNAMAN 9.0对测序获得的序列进行拼接,分别得到RH1基因和LWS基因cDNA序列全长。使用ORFfinder预测2个基因的开放阅读框(ORF)及其编码的蛋白序列。利用ExPaSy中的ProtParam对蛋白的氨基酸序列进行理化性质分析;分别用ProtScale和SignalP 4.1预测蛋白的亲水性和信号肽;用SMART预测蛋白的结构域;分别用SOPMA和SWISS-MODEL 预测蛋白的二级结构和三级结构;利用DNAMAN 9.0进行氨基酸多序列比对,以MEGA X中的邻接法(Neighbor-joining,NJ)构建系统发育进化树,各分支的置信度以Bootstrap为1000次进行检验。
1.6 实时荧光定量PCR检测
根据RH1基因和LWS基因序列,分别设计不同外显子的实时荧光定量PCR扩增引物(表1),以RPL13为内参基因(Liang et al.,2022)。反应体系:50 ng/μL cDNA模板1.5 μL,正、反向引物(10 μmol/L)各0.3 μL,2×SYBR Green qPCR Mix 7.5 μL,超纯水补足至15.0 μL。利用Light Cycler®96 PCR System定量PCR仪进行荧光定量PCR扩增,扩增程序:94 ℃预变性3 min;94 ℃15 s,58 ℃15 s,72 ℃20 s,进行40个循环。采用2-ΔΔCt法计算RH1基因和LWS基因在不同发育时期及不同组织的相对表达量,用Graphpad Prism 8.0和SPSS 21.0进行统计分析。
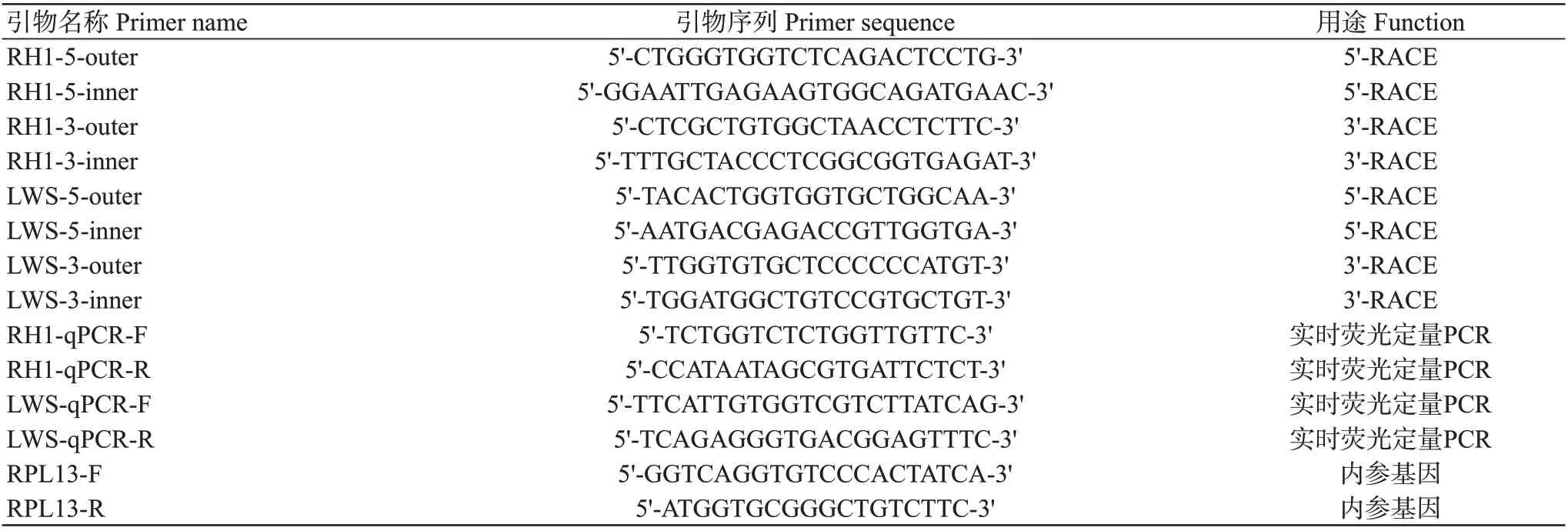
表1 引物序列信息Table 1 The primer sequences used in the experiment
2 结果与分析
2.1 红鳍笛鲷RH1和LWS基因cDNA全长序列分析结果
利用DNAMAN 9.0拼接出2个基因的cDNA全长序列,其中RH1基因(GenBank登录号为ON623871)cDNA序列全长1723 bp,ORF长度为1062 bp,编码353个氨基酸残基,3'-UTR长度为465 bp,5'-UTR长度为196 bp;LWS基因(GenBank登录号ON623870)cDNA序列全长为1302 bp,其中ORF长度为1074 bp,编码357个氨基酸残基,3'-UTR长度为213 bp,5'-UTR长度为15 bp,3'末端均具有明显的ploy(A)尾巴(图1和图2)。
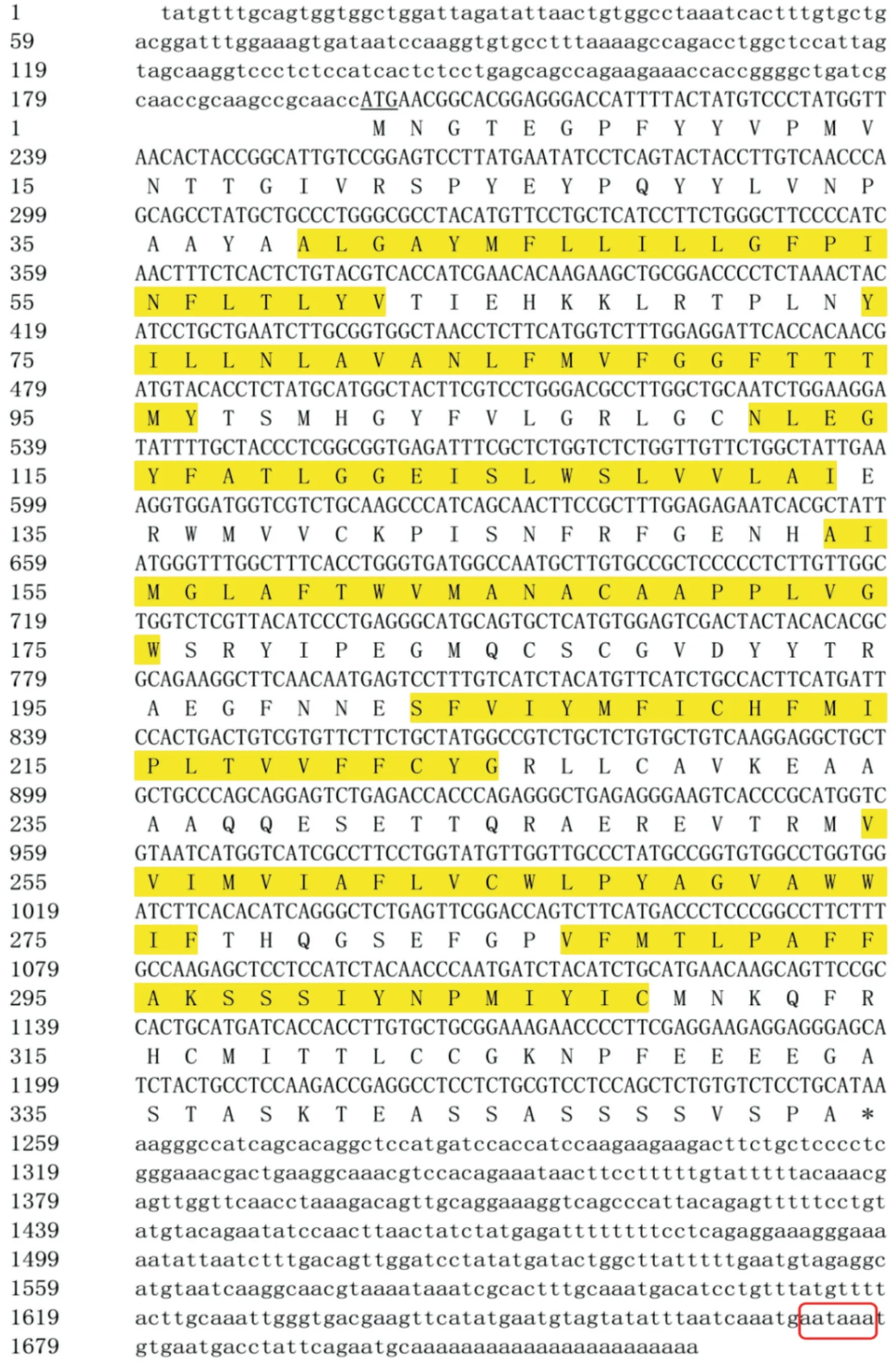
图1 红鳍笛鲷RH1基因全长cDNA序列Fig.1 Full-length cDNA sequence information of L.erythropterus RH1 gene
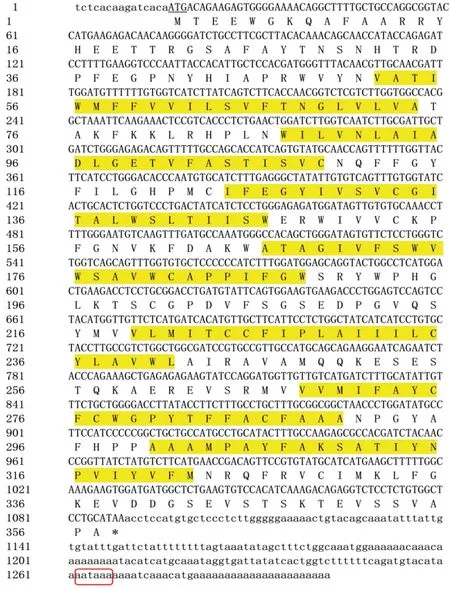
图2 红鳍笛鲷LWS基因全长cDNA序列Fig.2 Full-length cDNA sequence of L.erythropterus LWS gene
2.2 RH1和LWS氨基酸多序列比对及系统发育分析结果
从GenBank搜索其他物种的RH1氨基酸序列(表2)和LWS氨基酸序列(表3),再利用DNAMAN 9.0将获得的红鳍笛鲷RH1和LWS氨基酸序列与其他脊椎动物对应的氨基酸序列进行多序列比对,结果如图3所示。红鳍笛鲷RH1氨基酸序列与鱼类的RH1氨基酸序列相似性最高,其次是陆生脊椎动物,其中与鲈形目鱼类的相似性最高,最高达92%;LWS氨基酸序列的多序列比对结果与RH1氨基酸序列相似,与陆生哺乳动物LWS氨基酸序列的相似性最低,只有72%~78%,而与鱼类LWS氨基酸序列的相似性达80%~90%,其中与鲈形目的相似性较高,与鳉形目的相似性稍低,达85%~89%,与鲑形目的相似性最低,为74%~80%。系统发育分析结果表明,所有物种的RH1氨基酸序列聚为两大分支,其中鱼类聚为一支,陆生脊椎动物聚为一支,在鱼类分支中,红鳍笛鲷与金头鲷(Sparus aurata)、快乐东方鲷(Astatotilapia calliptera)等鲈形目的遗传距离最近,其次与鲽形目、鲑形目和鲤形目等鱼类均具有较近的亲缘关系(图3-A);所有物种的LWS氨基酸序列也聚为两大分支,陆生脊椎动物和鱼类同样各为一大分支,在鱼类分支中,红鳍笛鲷与奈里朴丽鱼(Pundamilia nyererei)的遗传距离最近,与孔雀花鳉(Poecilia reticulata)、海水青鳉(Oryzias melastigma)等鳉形目有较近的亲缘关系,与水牛(Bubalus bubalis)、灰短尾负鼠(Monodelphis domestica)等陆生脊椎动物的遗传距离最远,与传统的物种分类相一致(图3-B)。
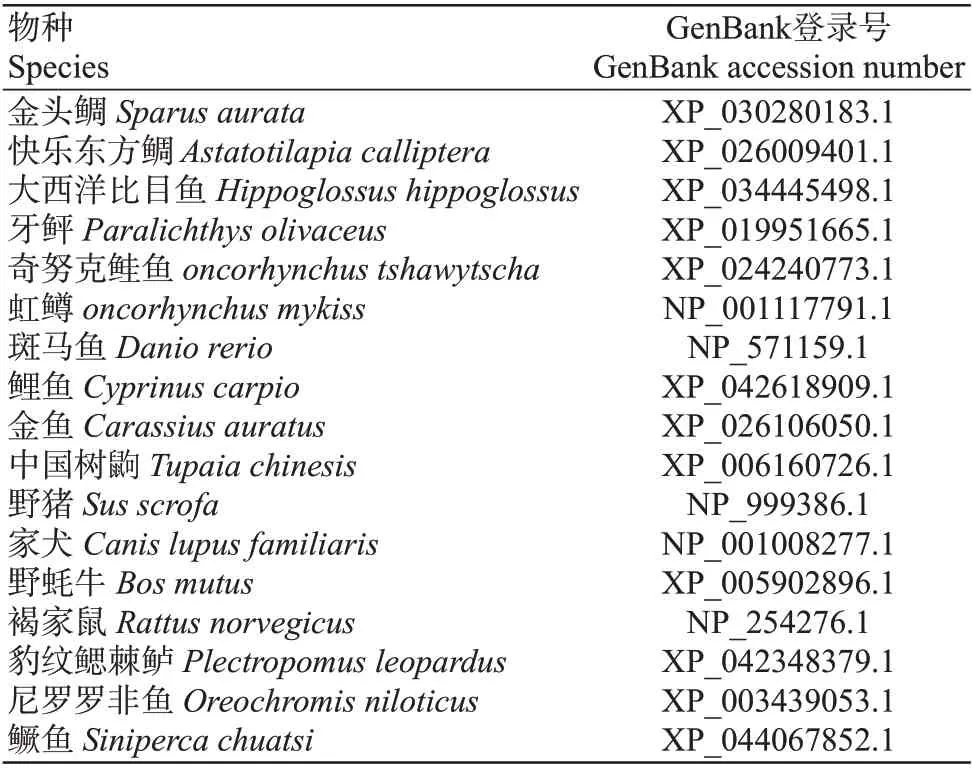
表2 不同物种RH1氨基酸序列的GenBank登录号Table 2 GenBank accession numbers of amino acid sequences of RH1 in different species
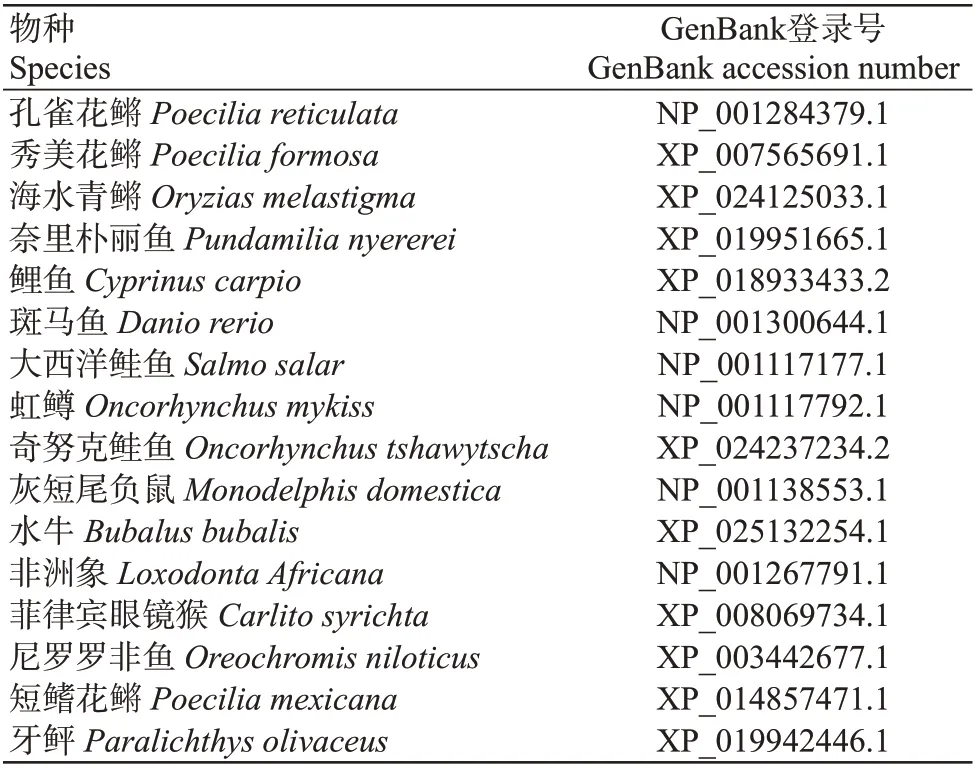
表3 不同物种LWS氨基酸序列的GenBank登录号Table 3 GenBank accession numbers of amino acid sequences of LWS in different species
2.3 红鳍笛鲷RH1和LWS蛋白的理化性质分析结果
ProtParam预测结果显示,RH1和LWS蛋白的分子量大小分别为39.57和40.22 kD,理论等电点(pI)分别为6.31和8.36,其中RH1蛋白带有22个负电荷残基(Asp+Glu)和20个正电荷残基(Arg+Lys),总平均疏水指数值(GRAVY)为0.415,属于疏水性蛋白,LWS蛋白带有23个负电荷残基(Asp+Glu)和27个正电荷残基(Arg+Lys),总平均疏水指数值为0.395,同样属于疏水性蛋白。亲水性预测结果显示,RH1和LWS蛋白的最小亲水性指数分别为-2.689和-2.933,最大亲水性指数分别为3.544和3.178(图5)。信号肽预测结果(图6)显示,2个视蛋白均无信号肽,证明RH1和LWS蛋白均属于非分泌型蛋白。据SMART在线网站预测结果(图7)显示,2个视蛋白均含有典型的7个跨膜结构域,其中RH1还含有一个Rhodopsin_N结构域。
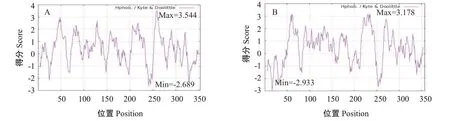
图5 红鳍笛鲷视蛋白RH1(A)和LWS(B)的亲/疏水性预测结果Fig.5 Hydrophilicity/hydrophobicity prediction of L.erythropterus opsins RH1(A)and LWS(B)
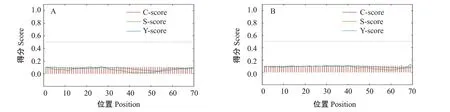
图6 红鳍笛鲷视蛋白RH1(A)和LWS(B)的信号肽预测结果Fig.6 Signal peptide prediction of L.erythropterus opsins RH1(A)and LWS(B)
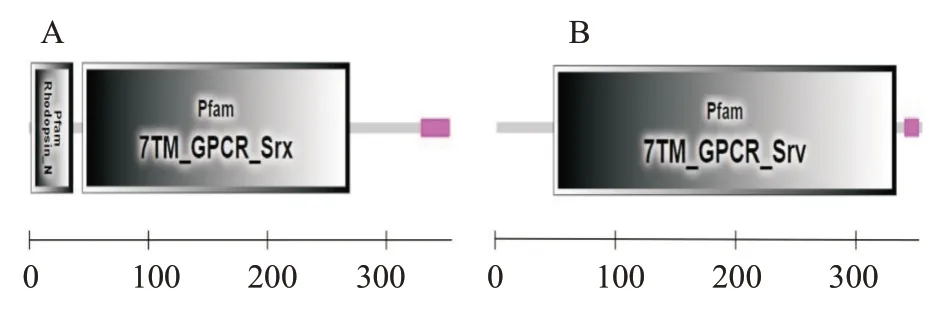
图7 红鳍笛鲷视蛋白RH1(A)和LWS(B)的蛋白结构域预测结果Fig.7 Protein domains prediction of L.erythropterus opsins RH1(A)and LWS(B)
2.4 红鳍笛鲷RH1和LWS的蛋白结构预测结果
利用SOPMA在线分析2个视蛋白的二级结构,结果显示,RH1的二级结构包括α-螺旋(42.21%)、延伸连(16.43%)、β-转角(3.97%)和无规则卷曲(37.39%);LWS的二级结构包括α-螺旋(44.26%)、延伸连(16.53%)、β-转角(2.52%)和无规则卷曲(36.69%)(图8)。
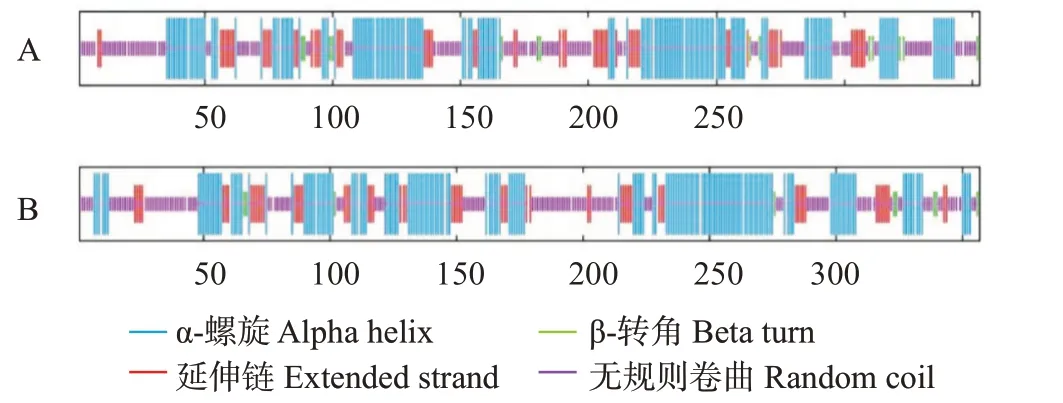
图8 红鳍笛鲷RH1(A)和LWS(B)蛋白二级结构预测结果Fig.8 Protein secondary structure prediction of L. erythropterus RH1(A)and LWS(B)
利用SWISS-MODEL预测RH1和LWS蛋白的三级结构,结果显示,这两个蛋白的三级结构相似,均具有典型的7个跨膜结构域(图9)。
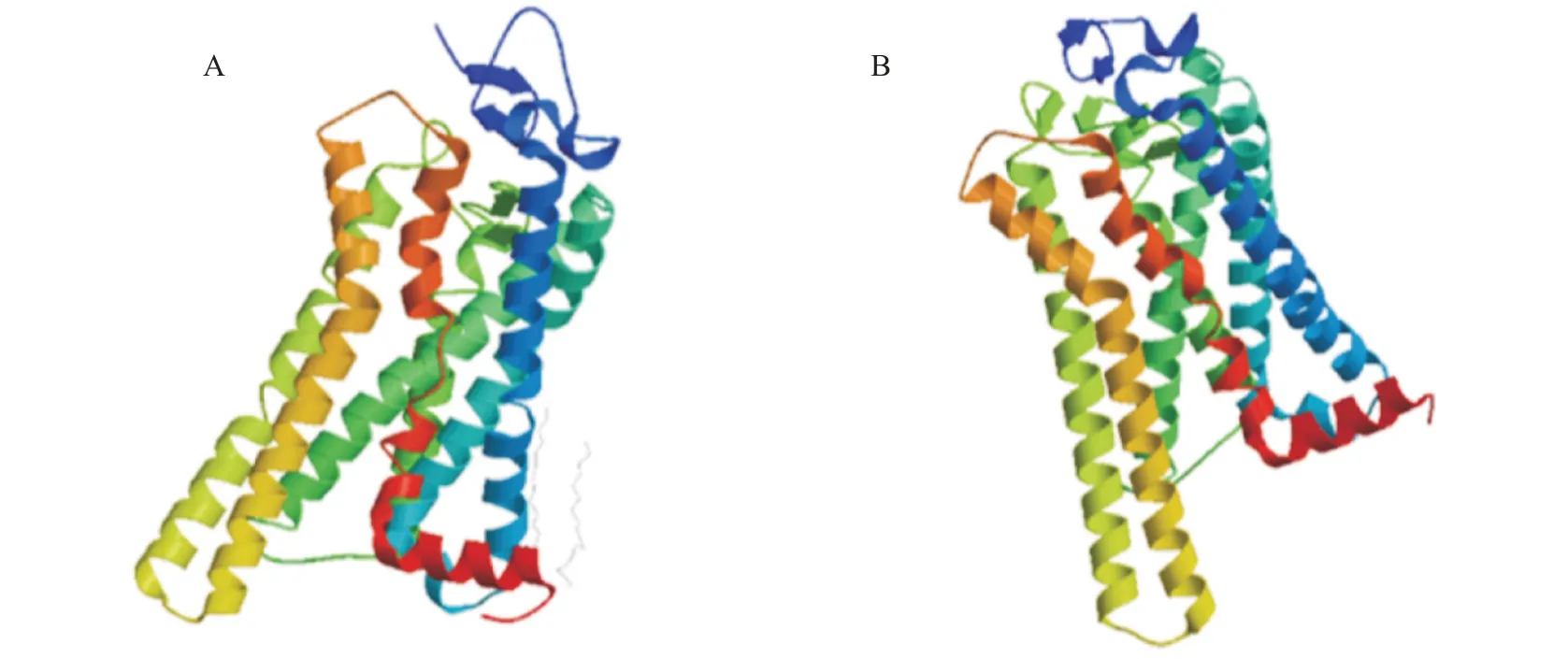
图9 红鳍笛鲷视蛋白RH1(A)和LWS(B)的蛋白三级结构预测结果Fig.9 Protein tertiary structure prediction of L.erythropterus opsin RH1(A)and LWS(B)
2.5 红鳍笛鲷RH1基因和LWS基因时空表达分析结果
采用实时荧光定量PCR检测红鳍笛鲷RH1基因和LWS基因在早期发育11个阶段和成鱼8个不同组织的表达情况,结果如图10和图11所示。在早期发育过程中,RH1基因的相对表达量呈现先降低后升高的趋势,在孵化出膜前的下包1/2期和胚孔封闭期均有微弱表达,然后相对表达量降低,趋近于0,孵化出膜后相对表达量逐渐升高,在15 d和20 d达最高值,显著高于其他时期的相对表达量(P<0.05,下同);在成鱼各组织中,RH1基因在视网膜中的相对表达量显著高于在其他7个组织中的相对表达量。LWS基因在孵化出膜后的20 d相对表达量最高,显著高于其他时期,其早期发育的表达水平变化模式与RH1基因相似,LWS基因同样具有组织表达特异性,在视网膜上的相对表达量显著高于其他组织,而其他组织间的相对表达量无显著差异(P>0.05)。
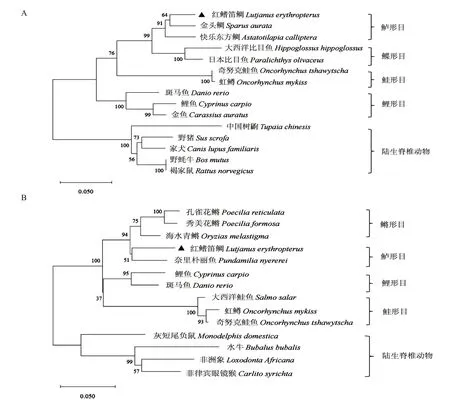
图4 基于红鳍笛鲷RH1(A)和LWS(B)氨基酸序列构建的系统发育进化树Fig.4 Phylogenetic tree based on the amino acid sequences of L.erythropterus RH1(A)and LWS(B)
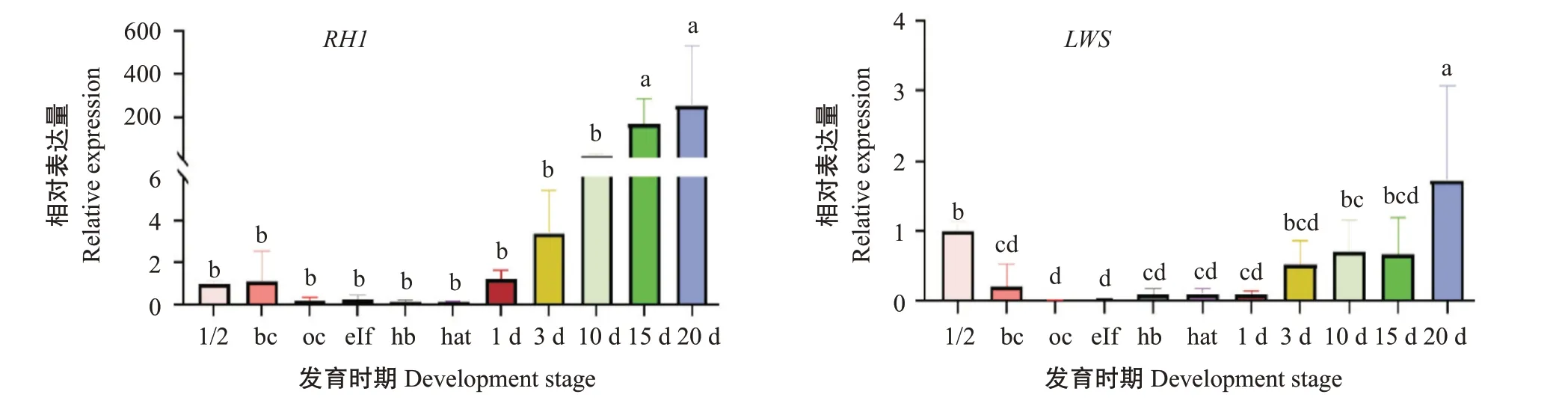
图10 RH1和LWS基因在红鳍笛鲷早期发育的表达情况Fig.10 Expression of RH1 and LWS genes in early development of L.erythropterus
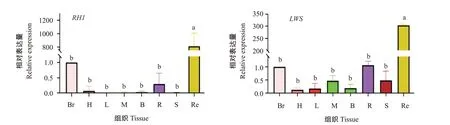
图11 RH1和LWS基因在红鳍笛鲷不同组织的表达情况Fig.11 Expression of RH1 and LWS genes in various tissues of L.erythropterus
3 讨论
3.1 RH1和LWS基因序列特征和系统发育分析
本研究通过RACE克隆红鳍笛鲷RH1基因和LWS基因cDNA全长,分别为1723和1302 bp,二者分别是视杆细胞和视锥细胞中的视蛋白(Yokoyama et al.,2000),是具有典型的7个跨膜结构域的膜蛋白。通过氨基酸多序列比对和系统发育分析发现,红鳍笛鲷和其他物种的视蛋白基因序列相似性很高,其中与鱼类RH1氨基酸序列相似性达84%~92%,而与陆生脊椎动物的相似性稍低,只有68%~74%,LWS氨基酸多序列比对结果与RH1氨基酸序列类似。系统发育分析结果显示,基于RH1和LWS氨基酸序列的相似性构建的系统发育进化树大致相同,所有物种可分为五大分支,红鳍笛鲷先与同属于鲈形目的鱼类聚为一支,其次与亲属关系较远的鲽形目、鲑形目和鲤形目等聚为一支,最后再与亲属关系最远的陆生脊椎动物聚为一支。2个视蛋白基因构建的系统发育进化树所体现的高级阶元亲缘关系与传统分类学和分子分类学基本一致(Allen et al.,1985;Guo et al.,2016),表明视蛋白基因同样适用于高级阶元的系统发育分析(刘楚吾等,2015)。
3.2 RH1和LWS基因的进化分析
视蛋白基因对鱼类的进化和物种的形成具有重要作用(刘楚吾等,2015),自Archer等(1992)研究大堡礁笛鲷属鱼类视蛋白λmax与水色的关系以来,珊瑚礁鱼类的视蛋白基因研究逐渐引起学者的关注,从而获得较多成果,从单一的形态学数据和视蛋白λmax数据分析逐渐转为以分子生物学手段对视蛋白基因介导环境适应性的研究。研究表明,RH1等视觉视蛋白基因主要起感受暗视觉的调控功能,对于海洋物种在海洋底栖环境下摄食、逃避敌害等方面发挥重要的作用,推测这些视觉视蛋白基因在昏暗且光谱受限制的海洋环境下承受更强的选择压力(Larmuseau et al.,2011)。多数脊椎动物RH1的λmax约500 nm(陈新页等,2015;Hart,2020;Sato et al.,2021),且根据栖息水域深度不同,RH1的λmax也会发生相应的红移或蓝移,表明大多数海水鱼类通过改变RH1氨基酸序列中的某些氨基酸残基位点来适应海洋光环境的变化(Larmuseau et al.,2011;Shozo,2012),在不同栖息水域环境下吸收不同波长的光线反映出鱼类的适应性进化。相对于淡水鱼类,海洋鱼类要面对深海更复杂的环境及光线变化,导致其视蛋白基因分化程度远高于淡水鱼类,发生基因复制的概率也会更高。LWS是一类负责参与动物视觉中红绿色成像的视蛋白,又被称作红—绿视蛋白(谢子强等,2015)。在视蛋白基因中,LWS基因分化程度最高,受各方面的选择压力也最大(刘旦,2012)。因此,LWS基因是5种视蛋白基因中探究分子进化和遗传学的理想基因。在笛鲷属鱼类中,LWS基因的选择性进化在笛鲷鱼类生态重叠区的物种分化过程中可能起到重要的作用(王中铎,2009)。
3.3 RH1和LWS基因的表达规律
本研究中RH1基因和LWS基因在早期发育的不同阶段和成鱼不同组织均呈现出非常相似的表达模式,视蛋白基因在鱼类不同发育阶段的表达变化模式,体现了鱼类自身眼睛发育及适应不同光境下的强大调节机制(Carleton et al.,2008)。在孵化出膜前,红鳍笛鲷2个视蛋白基因在原肠下包1/2期和胚孔封闭期均有较高表达,其余阶段表达量较低,可能与鱼类眼睛器官的形成与胚层分化和神经分化有关(谢子强等,2015),其原因是鱼类神经系统的形成与原肠胚的下包有关,原肠胚的下包影响了眼睛的发育,且与神经胚的形成密切相关。作为在视网膜上介导明暗视觉的视蛋白,红鳍笛鲷早期阶段RH1基因和LWS基因的表达可能与自身眼睛的开始发育有关。在孵化出膜后,红鳍笛鲷2个视蛋白基因的表达量均开始上升,其表达模式与大多数鱼类一致(Chang et al.,2020;Wang et al.,2021;Tang et al.,2022)。红鳍笛鲷在早期仔鱼阶段以浮游生活,且具有趋光性,此时波长较长的光能到达海水水层上方,而LWS主要感受长波的红色光(λmax约560 nm)(陈新页等,2015),表明对光环境的适应及摄食迫切需要相应视蛋白的表达回升(Luehrmann et al.,2018)。红鳍笛鲷由仔稚幼鱼转变为成鱼之后,开始以底栖生活为主(30~100 m),视杆蛋白RH1在对鱼类适应黑暗环境、摄食及躲避敌害等方面发挥重要作用,此时RH1基因在视网膜中占据绝对优势,而视锥蛋白的作用相对被弱化(Lupše et al.,2021)。RH1基因和LWS基因在红鳍笛鲷不同组织中的表达情况与其他鱼类相似,均在视网膜中高表达,尽管某些鱼类如蓝刻齿雀鲷(Chrysiptera cyanea)(Takeuchi et al.,2011)存在视网膜以外的光感受器,但视网膜依然是视蛋白基因的表达的主要部位。
4 结论
红鳍笛鲷2个视蛋白基因RH1和LWS基因的表达具有组织特异性,且在早期发育阶段的表达水平与红鳍笛鲷的生活习性变化相关,表明红鳍笛鲷RH1和LWS基因表达与其生活光环境的变化密切呼应,在早期发育变态阶段的光线调节,以及适应黑暗环境等方面发挥重要作用。
