烟草根系杂种优势表现及相关基因差异表达分析
2022-03-18段丽丽莫泽君柯渔洲王平松曾帅波刘仁祥
皮 凯,黄 莺,段丽丽,莫泽君,罗 雯,柯渔洲,王平松,曾帅波,刘仁祥
(贵州大学烟草学院/贵州省烟草品质研究重点实验室,贵州 贵阳 550025)
0 引言
【研究意义】烟草(Nicotiana tabacumL.)是我国重要的经济作物,大田生育期120~130 d左右,植株高大,需要大量的水分和营养元素供生长发育,而这些物质均通过根系向地上部运输(刘国顺,2012)。根系是植物植物生长必不可少的源器官,具有吸收并运输土壤水分和无机盐、固定及支撑植株、合成氨基酸及蛋白质等功能(郭毓勉,2020),对烟株的生长发育和烟叶产量、品质有很大的影响(晁逢春等,2003)。此外,烟叶中的烟碱是通过根系合成后运输至叶片中,对烟草的吸食品质极其重要(莫泽君,2020)。利用杂种优势可获得烟碱含量和钾含量优于亲本的烟草杂交种(王国琴,2015;罗雯等,2022),而烟碱杂种优势的形成是因为杂交种具有更高的烟碱合成效率(Tian et al.,2018;莫泽君,2020),表明烟草根系可能是烟碱含量和钾含量杂种优势形成的基础,直接影响烟草植株发育及烟叶品质。因此,研究烟草根系杂种优势表现及相关基因的差异表达有利于解析烟草地上部分杂种优势的形成机制,对选育高产优质杂交种具有重要的指导意义。【前人研究进展】烟草根系由主根、侧根和不定根组成,其中侧根和不定根最发达,不定根约占烟草总根重的1/3,成为根系的主要部分(黄泽春等,2008)。晁逢春等(2003)研究表明烟草不定根中的烟碱含量显著高于主根和侧根中的烟碱含量,调控不定根的发育将对叶片烟碱含量发挥积极作用。目前对烟草根系发育的研究主要集中在营养物质(蔡赛男,2015;李玥等,2015)、遗传因素(许杰等,2017a;钟思荣,2018;Singh et al.,2018)、生长调节剂(王守刚等,2015;张兆扬等,2017;Song et al.,2019;张琳等,2019;冯长春等,2020)、栽培方式(王德俊等,2019)等方面。蔡赛男(2015)研究发现根系构型参数最大的烤烟品种K326烟叶中的氮含量、钾含量及干物质量的积累均达国际标准,烟叶品质较好,而根系构型参数最小的烤烟品种G80烟叶氮含量和钾含量相对较低,同时其烟叶干物质量的积累相对较少。李玥等(2015)研究表明缺氮处理促进K326和湘烟四号烤烟根系生长,然而地上部生物量明显下降。许杰等(2017b)研究表明烤烟F1代的根系具有较强的杂种优势,其中根体积和根系阳离子交换量(CEC)的遗传以基因的加性效应为主,烤烟根系活力和腺嘌呤核苷三磷酸(ATP)酶活性的遗传以基因的显性效应为主。王宇辰等(2018)研究表明低浓度La3+能促进烟草根系发育,显著提高了根系生物量和根活力,高浓度La3+则表现出一定抑制作用。王德俊等(2019)研究表明不同覆盖模式对烤烟根系活力具有显著影响,其中“旺长期揭膜+培土+覆盖稻草”覆盖模式的根系活力最高。冯长春等(2020)研究表明外源吲哚乙酸(IAA)处理显著提高了烤烟幼苗根系生物量,10 nmol/L处理较对照增幅分别为89.43%,不同浓度IAA处理的烤烟幼苗根系总长度、总表面积和体积均不同程度的增加。李秀春等(2020)对烟草施用火土灰处理能显著增加植株根系总根长、根系表面积、根系体积和烟株苗期生物量。近年来,在其他作物中与根系发育相关的基因已有大量研究,拟南芥AtFLA18、At-PIN3、AtYUC6和AtNAC2基因通过调控根系的发育和形成来改变根系结构(He et al.,2005;Omelyanchuk et al.,2016;Allelign et al.,2021);AtAUX1具有调节根的向地性和根毛发育的功能(Villaecija-Aguilar et al.,2022)。水稻OsCRL1和OsDGL1基因及生长素信号响应因子基因OsARF16在水稻侧根的发育形成和冠根起始中发挥关键作用(Liu et al.,2005;Qin et al.,2013;Lavarenne et al.,2019)。尚未见烟草根系发育相关基因的研究报道。【本研究切入点】本课题组前期研究表明,杂交种中更强的烟碱合成效率和钾离子吸收能力是地上部分烟叶烟碱含量和钾含量表现出杂种优势的重要原因(Tian et al.,2018;罗雯等,2022)。因此,推测杂交种烟叶中烟碱含量和钾含量杂种优势的形成可能与杂交种的根系发育有关,但关于烟草根系的杂种优势表现及相关基因差异表达分析尚未见报道。【拟解决的关键问题】本研究基于课题组前期研究基础(莫泽君,2020;罗雯等,2022),选用烟碱和钾含量性状具有杂交优势的杂交组合及其亲本为材料,通过测定根系长度、数量和体积,分析了杂交种根系优势的表现,采用实时荧光定量PCR(qRT-PCR)检测烟草杂交种根系优势相关基因的差异表达,为深入解析烟叶烟碱含量和钾含量杂种优势的形成机制提供参考。
1 材料与方法
1.1 试验材料
供试的3个母本品种(系)为K326、Va116和G70,4个父本品种(系)为南江三号、GDH88、毕纳一号和韭菜坪2号,按照NCⅡ设计将其配制成12个杂交组合,所有材料均由贵州省烟草品质研究重点实验室制种和保存。RNAprep Pure Plant Kit试剂盒、Fast-King gDNA Dispelling RT SuperMix试剂盒和Talent qPCR PreMix(SYBR Green)试剂盒购自天根生化科技(北京)有限公司。主要仪器设备:WinRHIZO根系扫描仪LA2400(Regent,加拿大)、CFX96 Real-Time System(Bio-Rad,美国)、凝胶成像系统(Bio-Rad,美国)、冷冻离心机(Thermo,美国)和PowerPac电泳仪(Bio-Rad,美国)。
1.2 试验设计
试验于2021年在贵州省安顺市杨武乡贵州大学烟草科研基地进行,采用漂浮育苗,待烟株长至6片真叶时,每个材料选取3株大小一致、生长良好的烟草幼苗。将根系洗净后测定根系表型,从每株中选取根系鲜活末端,立即用液氮处理后存入-80 ℃冰箱,供后续RNA的提取和基因表达量的测定。
1.3 根系性状测定方法
对洗净的完整根系使用WinRHIZO根系扫描仪LA2400进行扫描,获得清晰的根系结构图像。使用WinRhizo Pro 2013a对烟草根系图像进行分析,得到根长、根体积和根数量等特征参数。
1.4 基因表达量检测
1.4.1 总RNA提取及cDNA合成 采用RNAprep Pure Plant Kit试剂盒提取总RNA,按FastKing gDNA Dispelling RT SuperMix试剂盒说明反转录合成cDNA第一链。
1.4.2 实时荧光定量PCR检测 在NCBI数据库中搜索8个在拟南芥和水稻中与根系发育相关的基因(表1),获得其cDNA全长序列,通过烟草K326基因组(https://solgenomics.net/organism/Nicotiana_tabacum/genome)比对到高度同源的序列后,根据此序列利用Primer 5.0设计引物,并委托北京擎科生物科技有限公司合成。按照Talent qPCR PreMix(SYBR Green)试剂盒说明对这些基因进行实时荧光定量PCR检测。反应体系20.0 μL:2×Talent qPCR Pre-Mix 10.0 μL,上、下引物(10 μmol/L)各0.6 μL,cDNA模板2.0 μL,ddH2O 补足至20.0 μL。扩增程序:95 ℃3 min;95 ℃5 s;退火温度(表1)10 s,75 ℃15 s,进行40个循环。每个样品设3次技术重复,以K326为对照,采用比较阈值法(2-ΔΔCt)法计算各基因相对表达量(Livak and Schmitten,2001)。
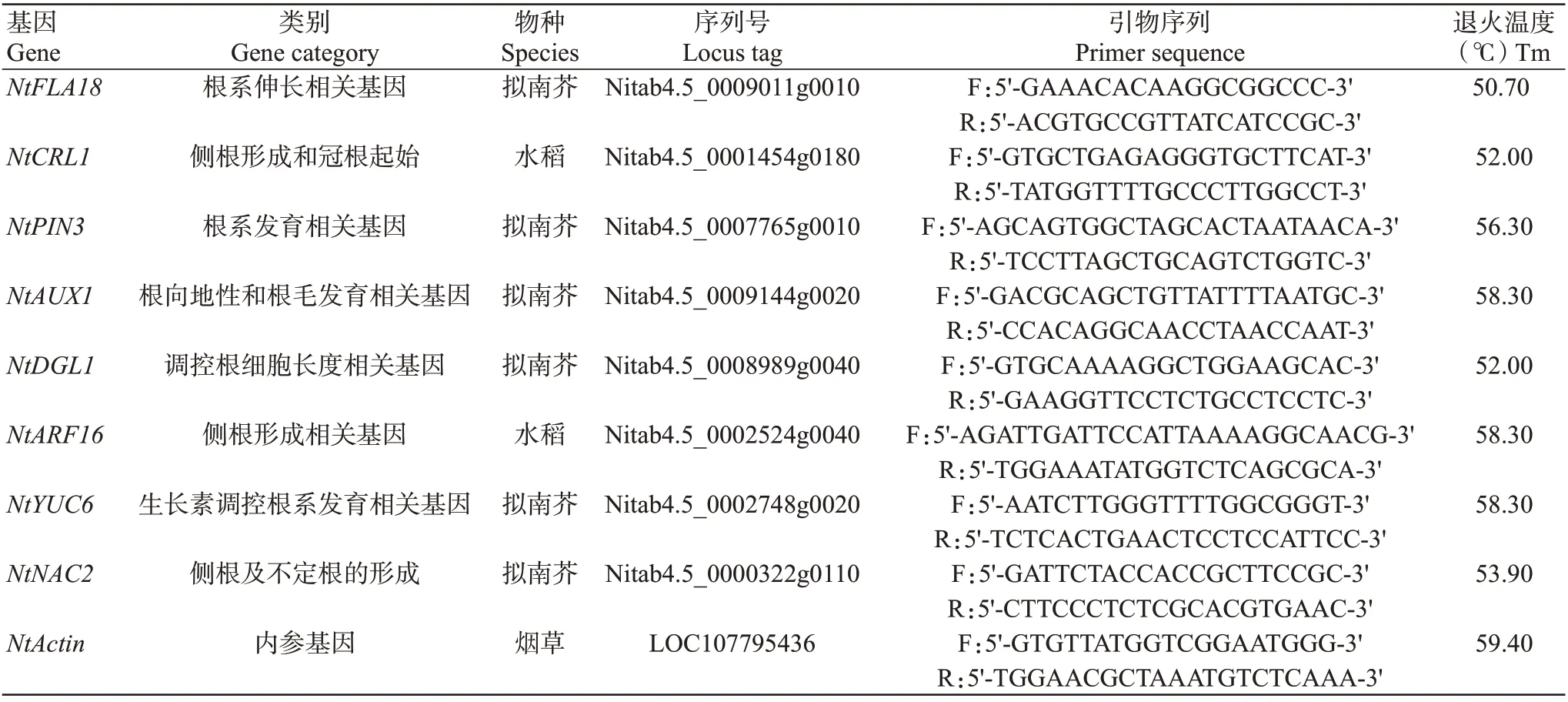
表1 根系发育相关基因及其引物信息Table 1 Root development related genes and their primers
1.5 杂种优势计算方法
以中亲优势进行根系性状的杂种优势分析。中亲优势是指杂种F1代某一数量平均值与双亲同一性状平均值的差值占双亲均值的百分比。参照陈泽辉(2009)方法计算根系长度、数量和体积的杂种优势,计算公式如下:
式中,F1表示杂交种。
1.6 统计分析
采用Excel 2013和DPS 7.05对相关数据进行统计分析。
2 结果与分析
2.1 亲本及杂交组合间根系的差异分析
由表2可知,7个亲本材料的根系长度为94.80~476.77 cm,根系数量为343~820条,根系体积为0.18~0.61 cm3,其中,Va116的根系数量和长度显著高于其余6个亲本(P<0.05,下同),根系体积(除显著小于南江三号外)均显著高于其他5个亲本材料。此外,韭菜坪2号和毕纳1号的根系长度、数量和体积均显著低于其他5个亲本,结果表明亲本间根系的各性状存在遗传差异,适于用来开展烟草根系的遗传利用研究。
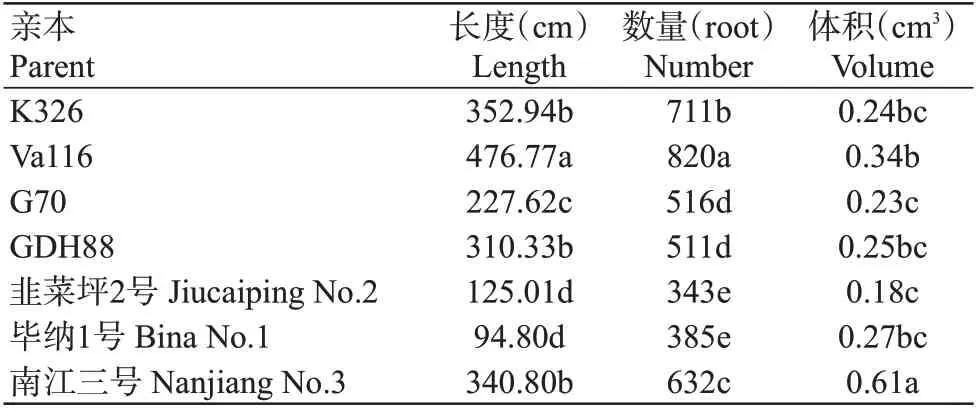
表2 亲本间根系性状差异分析Table 2 Analysis of differences in root traits among parents
由表3可知,12个杂交组合的根系长度为285.56~734.75 cm,根系数量为576~1184条,根系体积为0.41~1.01 cm3;K326×GDH88的根系长度和根系数量显著高于其他杂交组合,G70×南江三号的根系体积显著高于其他杂交组合;各性状的变异系数为19.97%~29.68%。由此可知,12个杂交组合的根系性状存在明显差异和遗传变异,表明利用杂种优势可选育出不同根系长度、数量和体积的杂交种。
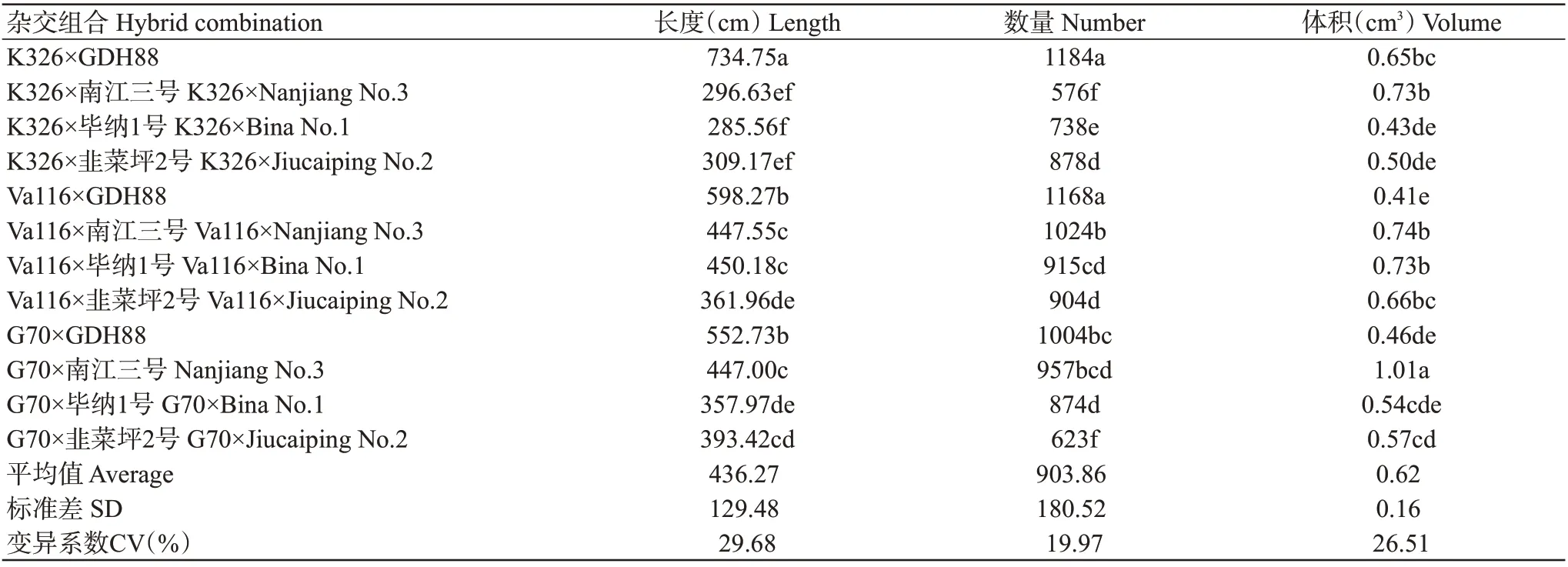
表3 12个杂交组合间根系性状差异分析Table 3 Analysis of differences in root traits among hybrid combinations
2.2 烟草根系的遗传特点分析结果
2.2.1 遗传力及配合力方差 由烟草根系各性状的遗传力及配合力方差结果(表4)可知,烟草根系长度、数量和体积的广义遗传力分别为95.49%、92.12%和85.76%,表明根系长度、数量和体积的遗传力高,受环境影响较小。根系长度的一般配合力大于特殊配合力,表明根系长度主要受到基因加性效应的影响;根系数量和体积的特殊配合力方差高于一般配合力方差,表明这两个性状主要受到基因非加性效应的作用,适于采用杂种育种方法进行遗传改良。
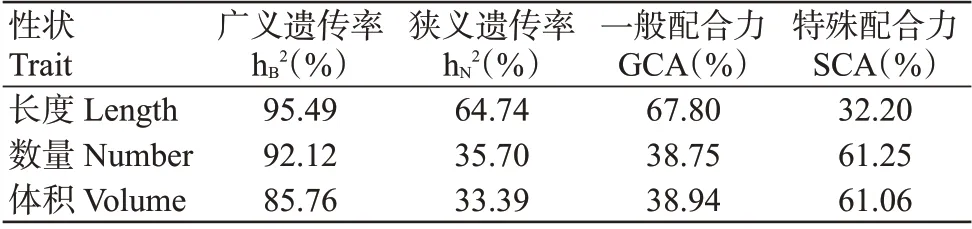
表4 烟草根系各性状的遗传力和配合力方差Table 4 Variance of heritability and combining ability of tobacco root traits
2.2.2 一般配合力和特殊配合力分析 一般配合力是指一个自交系与其他多个自交系杂交后遗传给F1代性状的平均表现,由基因的加性效应决定,可稳定遗传。选择一般配合力高的材料作亲本,更易获得优良的杂交组合。由7个亲本根系性状的一般配合力(表5)可知,在根系长度和体积2个性状上,分别有3个亲本的一般配合力表现为正向效应,4个亲本的一般配合力表现为负向效应;在根系数量上,有2个亲本的一般配合力表现为正向效应,5个亲本的一般配合力表现为负向效应。通过进一步分析发现,GDH88和Va116可作为改良根系长度和数量的优良亲本,南江三号、Va116和G70可作为改良根系体积的优良亲本。
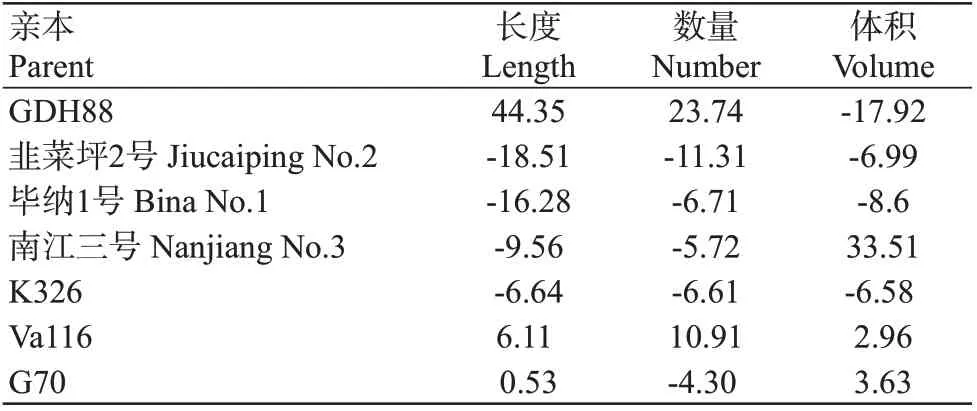
表5 7个亲本根系各性状的一般配合力相对效应值Table 5 Relative effect values of GCA of root traits of 7 parents
特殊配合力是指两个特定自交系所组配性状的表现水平,由非加性效应基因决定,即受基因的显性和上位性互作效应影响,只能在特定的组合中由双亲基因互作反映出来,不可遗传。特殊配合力的高低可为优良杂交组合的选育提供依据。由12个杂交组合根系性状的一般配合力(表6)可知,5个杂交组合根系长度的特殊配合力表现为正向效应,6个杂交组合的根系数量特殊配合力表现为正向效应,4个杂交组合的根系体积特殊配合力表现为正向效应。通过进一步分析发现,K326×GDH88、Va116×毕纳1号和G70×韭菜坪2号是根系长度较为理想的烤烟杂交组合;G70×南江三号、K326×韭菜坪2号和K326×GDH88是根系数量较为理想的烤烟杂交组合;K326×GDH88、G70×南江三号和Va116×毕纳1号是根系体积较理想的烤烟杂交组合。可见,采用杂种优势改良烟草根系性状是行之有效的。
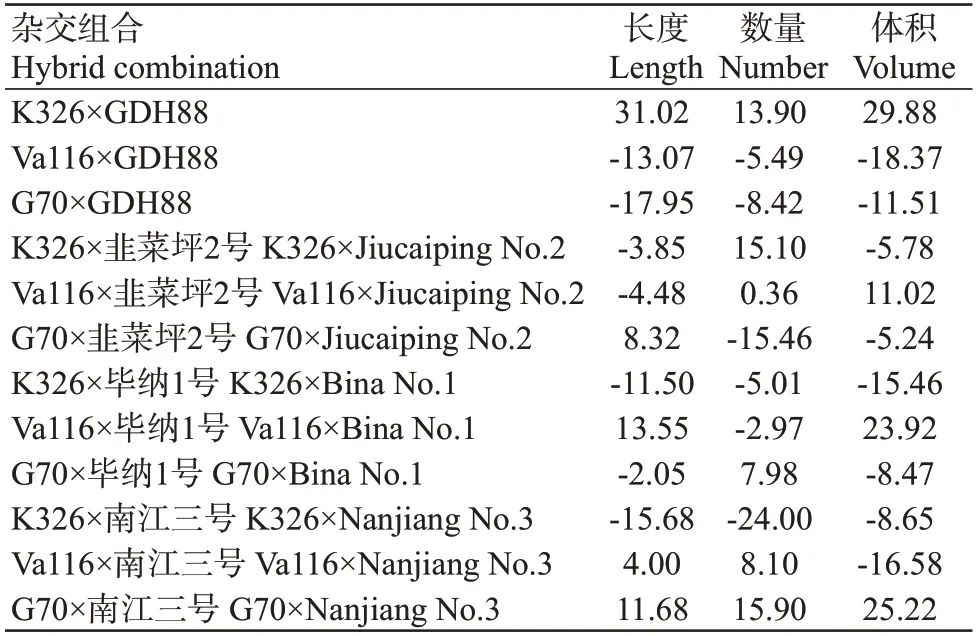
表6 12个杂交组合根系各性状的特殊配合力相对效应值Table 6 Relative effect values of SCA of root traits of 12 hybrid combinations
2.3 烟草根系的杂种优势表现
通过对不同杂交组合烟草根系3个性状的中亲优势进行差异性分析,结果如表7所示。12个杂交组合的3个性状均表现为强优势,正向中亲优势组合率分别为91.67%、91.67和100.00%,结果表明烟草根系具有普遍的杂种优势。后续从12个杂交组合中随机选择一个杂交组合(K326×韭菜坪2号),通过分析不同基因在该杂交组合及其亲本中的表达情况,进一步探讨烟草根系杂种优势的形成原因。
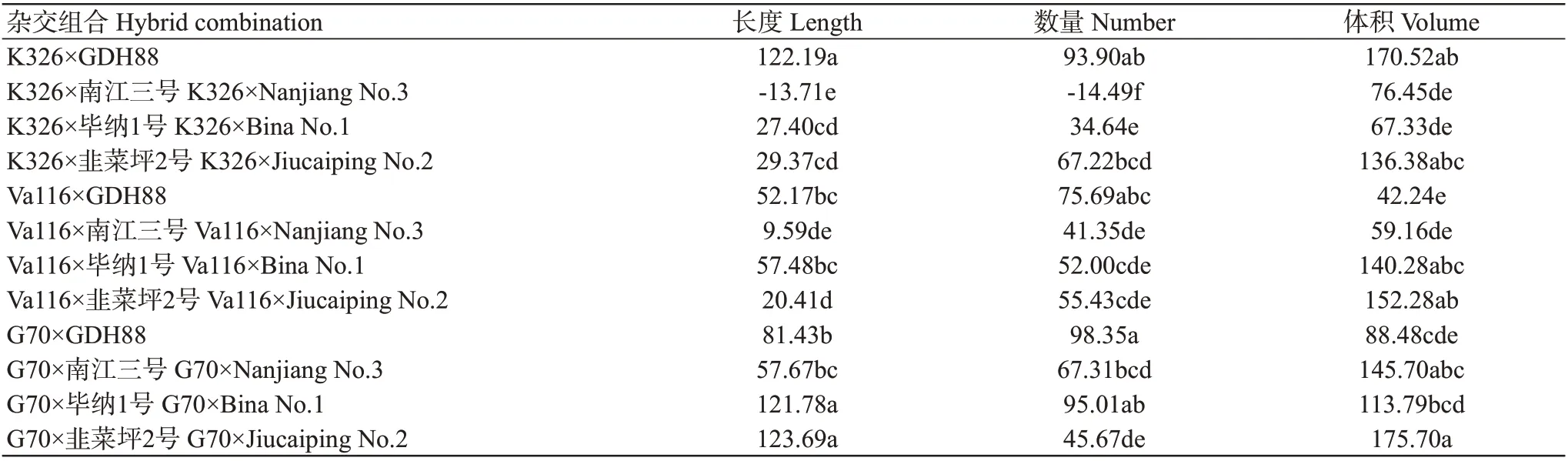
表7 不同杂交组合根系各性状的杂种优势差异分析(%)Table 7 Heterosis difference analysis of root traits of different hybrid combinations(%)
2.4 烟草根系及其杂种优势相关基因的筛选
2.4.1 烟草根系杂种优势及相关基因的中亲表达优势 利用实时荧光定量PCR检测8个根系发育相关基因在K326×韭菜坪2号及其亲本中的相对表达量(图1),并计算各基因的中亲表达优势(表8),结果显示NtFLA18、NtAUX1和NtDGL1基因表现为负向优势;NtCRL1、NtPIN3、NtARF16、NtYUC6和NtNAC2基因表现为正向优势。通过分析根系发育相关基因的中亲表达优势与根系各性状杂种优势的关系,结果(表9)表明不同根系发育相关基因的中亲表达优势与根系性状杂种优势之间存在一定的相关性,其中,NtAUX1和NtARF16基因的中亲表达优势与根系长度杂种优势呈显著(P<0.05,下同)或极显著(P<0.01,下同)相关,相关系数为-0.98和1.00,说明NtAUX1和NtARF16基因对根长度杂种优势的形成起关键调控作用;NtFLA18基因的中亲表达优势与根系数量和体积的杂种优势呈显著或极显著负相关,相关系数为-0.98和-1.00,说明NtFLA18基因对根系数量和体积杂种优势的形成起关键调控作用。

表8 强优势杂交组合的基因中亲相对表达量(%)Table 8 Relative expression of genes in strong heterosis hybrid combination(%)

表9 根系发育相关基因中亲表达优势与根系性状杂种优势的相关系数Table 9 Correlation coefficient between parental expression heterosis in genes related to root development and heterosis of root traits
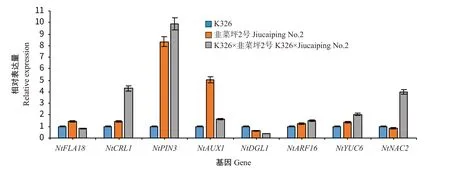
图1 根系发育相关基因在亲本及杂交种中的相对表达量Fig.1 Relative expression of genes related to root development in parents and hybrids
2.4.2 根系杂种优势相关基因在杂交组合中的差异
表达分析 由烟草根系杂种优势形成的关键基因在K326×韭菜坪2号及其亲本中的表达情况(图2)可知,NtFLA18基因在K326×韭菜坪2号中的相对表达量较双亲下调表达,NtAUX1基因表现为显性表达模式,NtARF16基因的相对表达量较双亲上调表达,推测NtFLA18基因的下调表达、NtFLA18基因的显性表达及NtARF16基因的上调表达均与烟草根系杂种优势的形成有关。
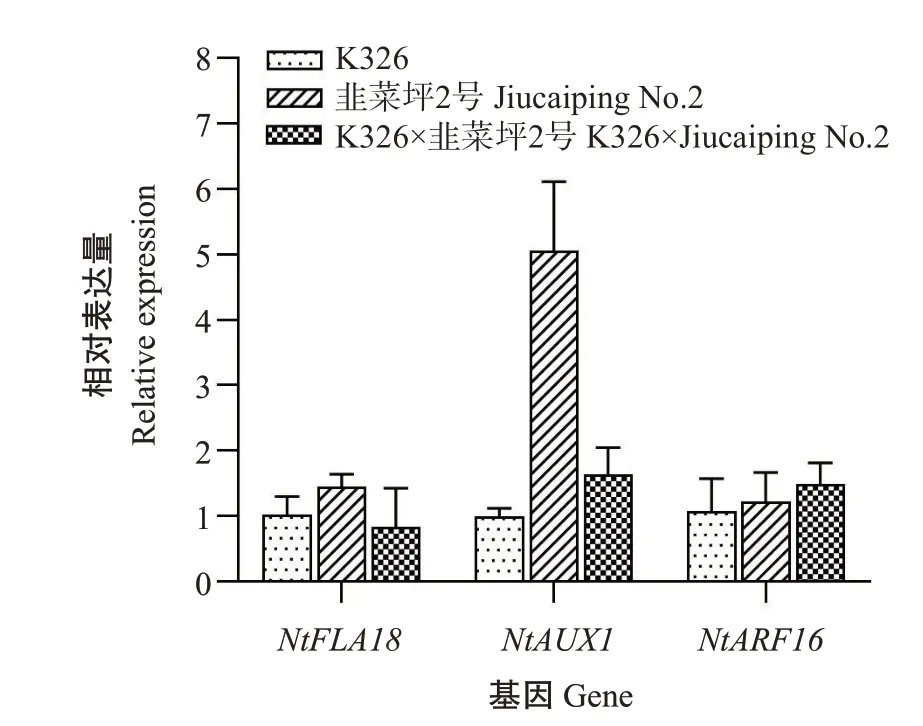
图2 根系杂种优势相关基因在杂交组合及其亲本中的相对表达量Fig.2 Relative expression of root heterosis related genes in hybrid combinations and their parents
3 讨论
根系是植物吸收水分和矿物质的重要器官,对植物地上部的发育有重要影响,根系的生物量(干重、数量或根长)与株高、最大叶长宽呈正相关,并且与烟叶中的烟碱和钾含量呈显著正相关(何礼军,2006)。目前,对于烟草根系的研究主要集中在连作障碍(于会泳等,2014)、营养调控(罗勇等,2019;张玉宁,2020)、土传病害(娄晓平等,2019;张成省,2020)、逆境胁迫(梁栋,2021)及作为砧木提高烟叶钾含量(Hu et al.,2021)等方面,而有关烟草根系植物学性状及其遗传的研究鲜见报道。许杰等(2017b)对烤烟主要根系生理特性的遗传效应进行分析,结果发现F1根系阳离子交换量、根系活力和ATP酶活性的杂种优势较强。本研究对烟草根系的植物学性状进行遗传分析,结果表明烟草根系长度、数量和体积均具有普遍的杂种优势,与许杰等(2017b)对根系体积的研究结果一致。
王秀全等(2002)研究表明玉米根体积、鲜干重、数量和总长的遗传均以加性效应为主。本研究发现根系长度主要受基因的加性效应控制,根系数量和根系体积主要受基因的非加性效应控制。可见,根系长度的研究结果与王秀全等(2002)的研究结果一致,但根系数量和体积的研究结果与该文献不一致,可能是由于试验材料的不同而导致。南文华等(2011)研究表明,根长、根鲜质量、根体积、活根数和根干质量的变异系数分别为14.43%~36.00%。本研究发现烟草根系长度、数量和体积的变异系数为19.97~29.68%,与南文华等(2011)的研究结果相近。赵丽萍等(2014)对甘蔗根系性状的遗传效应进行研究,结果发现根长、根表面积、根尖数、根干重和根体积的遗传力均大于60%,并指出根长、根表面积和根直径可作为甘蔗根系育种的选择指标。本研究结果表明根系长度、数量和体积的广义遗传率为85.76~95.49%,基因效应在遗传中起重要作用,与赵丽萍等(2014)的研究结果基本一致,在今后的育种工作中可利用杂种优势选育出根系发达的杂交种。
杂种优势的应用促进了烟草育种的发展,但其机理尚不清楚。继基因组组成差异及基因效应研究之后,基因表达差异成为探寻杂种优势分子机理新的切入点(许晨璐等,2013;滕建辉等,2021)。滕建辉等(2021)研究表明FPF1和PHYC基因的下调表达有利于调控烟草叶片数杂种优势的形成。罗雯等(2022)研究表明AKT1、NtKC1、KAT1、NKT1和NtHAK基因的上调表达与杂交组合烟叶钾含量优势有关。本研究发现,基因差异表达与根系杂种优势表现呈显著相关性,其中,NtAUX1和NtARF16基因表达与根系长度杂种优势呈显著或极显著相关性,NtAUX1基因在K326×韭菜坪2 号中呈显性表达模式,NtARF16基因呈上调表达模式。NtFLA18基因表达量与根系数量和体积杂种优势呈显著或极显著相关,在K326×韭菜坪2号中呈下调表达模式,但NtFLA18、NtARF16和NtAUX1基因如何调控根系杂种优势还需进一步探讨。随着高通量生物学技术的不断发展,今后可采用转录组、蛋白质组等技术挖掘与烟草根系杂种优势形成的相关基因,分析其表达模式,进一步探讨根系杂种优势形成的分子机理。
4 结论
烟草根系长度、数量和体积均具有较强的遗传率和普遍的杂种优势,可利用杂种优势选育出根系发达的烟草新种质。GDH88、Va116、南江三号和G70可作为改良根系的优良亲本;K326×GDH88、Va116×毕纳1号、G70×韭菜坪2号、G70×南江三号和K326×韭菜坪2号可作为培育优良根系构型的重要储备材料。
