低温胁迫对葛藤幼苗生长发育及生理特性的影响
2022-03-18赵文武黄莉娟赵丽丽王雷挺谢文辉
赵文武,黄莉娟,赵丽丽,王雷挺,谢文辉
(贵州大学动物科学学院,贵州 贵阳 550025)
0 引言
【研究意义】低温是植物生长周期中最常见的非生物胁迫之一。当环境温度长时间低于植物生长所需的适宜温度时,就会对植物形成低温胁迫,根据严重程度可分为冷害(一般为0~10 ℃)和冻害(一般在0 ℃以下)(宋静爽等,2019),植株遭受严重冻害时甚至会发生死亡。冷害多发生在我国高纬度的东北和高海拔的云贵高原等地区(蔡志欢和张桂莲,2018)。葛藤(Pueraria lobata)为豆科(Leguminosae)多年生藤本植物,在我国分布广泛,尤其在我国南方各地数量较多,其根、茎、藤均可入药,作为牧草和饲料加工原材料时可用于防治动物的某些疾病,具有良好的饲药合用特质。葛藤具有耐贫瘠、耐干旱、抗病虫害能力强等特点,还具有喜钙性及岩生性特征,能在土壤和水分严重亏缺的石漠化地区生长,可作为石漠化治理的先锋植物(陈俊等,2014)。作为高蛋白的饲草资源,葛藤藤茎生长旺盛,可常年提供青绿饲料(郑霞等,2018),能有效缓解我国南方高蛋白牧草季节性不均衡的问题。葛藤喜欢温暖的气候,适宜生长温度为22~26 ℃(陈俊等,2014),在冷害发生区,低温成为限制葛藤生长的首要因素。因此,研究葛藤在低温下的生长状况,对葛藤种质的耐低温鉴定及在西南地区的推广应用具有重要意义。【前人研究进展】生理生化指标可作为评价植物抗寒性的重要指标。正常环境下,细胞内活性氧(ROS)的产生和清除处于动态平衡(Mittler et al.,2011),低温会引起ROS大量积累,进而诱导膜脂过氧化并产生丙二醛(MDA)(Velikova et al.,2000)。植物会形成一系列复杂的酶和非酶防御系统来维持这一动态平衡(Uemura et al.,2003)。田丰等(2018)对低温处理后的小麦幼苗进行研究,发现超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性及脯氨酸(Pro)等渗透调节物质含量均随温度的降低而升高,以抵抗低温带给小麦的不良影响。李倩等(2021)指出当植物遇到低温冷害时可能会造成生理代谢活动失调,表皮细胞死亡。盖志佳等(2021)指出低温会导致种子萌发降低、产量与质量严重下降,抗病性减弱等不利影响。低温还会诱导植物体内激素活性和含量发生改变,进而对植物的生长代谢过程产生影响。逆境胁迫能诱导植物内源脱落酸(ABA)积累,提高植物的抗逆性(刘海卿等,2016),赤霉素(GA)可提高植物体内生长素的含量,调节自由水和束缚水比例,从而对植物的抗寒能力产生影响(Qu et al.,2016)。马学才等(2021)研究发现油菜可通过增加ABA合成,同时适当减少生长素(IAA)和GA合成来抵御低温。【本研究切入点】葛藤在我国西南地区可作为优质的牧草饲料来源和环境保护植物物种。目前,与葛藤相关的研究主要集中在其水土保持(丁艳芳,2003)、饲用价值(张倩,2011)、药用价值(尹乐斌等,2016;房伟,2017)、种子萌发(谭景晨等,2021)等方面,而针对葛藤对低温胁迫响应的研究鲜见报道。葛藤喜温,低温对其生长不利,对葛藤的耐低温性研究还有待加强。【拟解决的关键问题】以葛藤种质为试验材料,采用盆栽土培试验,设轻度和重度低温胁迫处理,对比分析不同低温处理下葛藤幼苗的生长速度、叶形态、内源激素和生理生化指标变化情况,探明葛藤幼苗耐冷性的生理生化机制,揭示葛藤响应低温的关键机制,为葛藤种质耐低温鉴定及及合理选育抗冷害性强的种质提供理论依据。
1 材料与方法
1.1 试验材料
供试葛藤种子由江苏园林绿化种子公司提供。采取盆栽试验,花盆长50 cm、宽20 cm、高15 cm,栽培基质为自然土∶蛭石∶腐殖质=1∶1∶1。
1.2 试验方法
挑选籽粒饱满、种胚完整、无霉变、大小基本一致的葛藤种子,先用次氯酸钠溶液浸泡20 min,再用蒸馏水冲洗4~5次后于常温下萌发,出苗后,每盆选取生长良好且长势基本一致的葛藤幼苗12株,移入盆栽,置于光照培养箱(昼/夜=25 ℃/20 ℃,光周期12 h/12 h,光照强度30000 LX,相对湿度75%)进行预培养一周。温度设2个水平梯度,分别为轻度低温胁迫(记为LD,昼/夜15 ℃/10 ℃)和重度低温胁迫(记为SD,昼/夜10 ℃/5 ℃);以昼/夜25 ℃/20 ℃为对照(CK)(条件同预培养)。每处理3个重复。在胁迫第10 d采用随机法每盆挑选3株葛藤,分别测量各项生长指标,并取新鲜的葛藤叶片用锡箔纸包住迅速置于液氮中速冻,保存于-80 ℃冰箱中用于测量各项生理指标。
1.3 测定项目及方法
1.3.1 生长指标测定 生长速度:每盆挂牌3株分别在胁迫前和胁迫最后1 d测量对应株高,计算生长速度,生长速度(cm/d)=(胁迫后高度-胁迫前高度)/生长天数。
叶形态:用Epson Perfection V800 Photo(印度尼西亚)扫描仪扫描第3片完全展开功能叶的叶形态,每盆3株。
干鲜比:每盆取3株完整的葛藤植株,洗净后吸水纸吸干称重,然后烘干至恒重再次称重,计算干鲜比。
1.3.2 生理生化指标测定 称重法测定水含量(来强等,2008);浸泡法测定相对电导率(高俊凤,2006);可溶性糖、Pro和MAD含量及SOD、POD、CAT和抗坏血酸过氧化物酶(APX)活性采用北京索莱宝科技有限公司试剂盒测定,测定方法均按照说明书进行操作。
采用北京索莱宝科技有限公司试剂盒通过酶联免疫吸附测定法(ELISA)进行植物组织内ABA、GA3、玉米素(ZT)、IAA等激素含量测定(Zhao et al.,2006)。
1.4 耐低温能力综合评价
采用主成分分析法,将各处理葛藤叶片的18个生长及生理生化指标进行降维,并筛选出抗冷性综合指标,最后采用隶属函数法综合评价葛藤的抗冷能力。
综合指标隶属函数值(U):
式中,Xj表示第j个综合指标;Xmin表示第j个综合指标的最小值;Xmax表示第j个综合指标的最大值。
综合指标权重(W):
式中,Wj表示第j个综合指标在所有综合指标中的重要程度,即权重;Pj为第j个综合指标的贡献率。
抗冷能力综合评价:
式中,D值为各低温胁迫下由综合指标评价所得的葛藤抗冷能力值。
1.5 统计分析
利用Excel 2019 进行数据处理,采用Origin 2019作图,以SPSS 22.0进行差异显著性、相关及主成分分析。
2 结果与分析
2.1 低温胁迫对葛藤幼苗生长的影响
2.1.1 生长速度、干鲜比、水含量 由图1可知,与CK相比,LD和SD处理下葛藤幼苗的生长速度(图1-A)分别显著下降72.19%和80.15%(P<0.05,下同);幼苗干鲜比(图1-B)分别上升7.59%和30.90%,SD处理上升显著;葛藤叶片水含量(图1-C)则随着低温胁迫程度的加强而降低,SD处理下较CK显著降低3.74%。
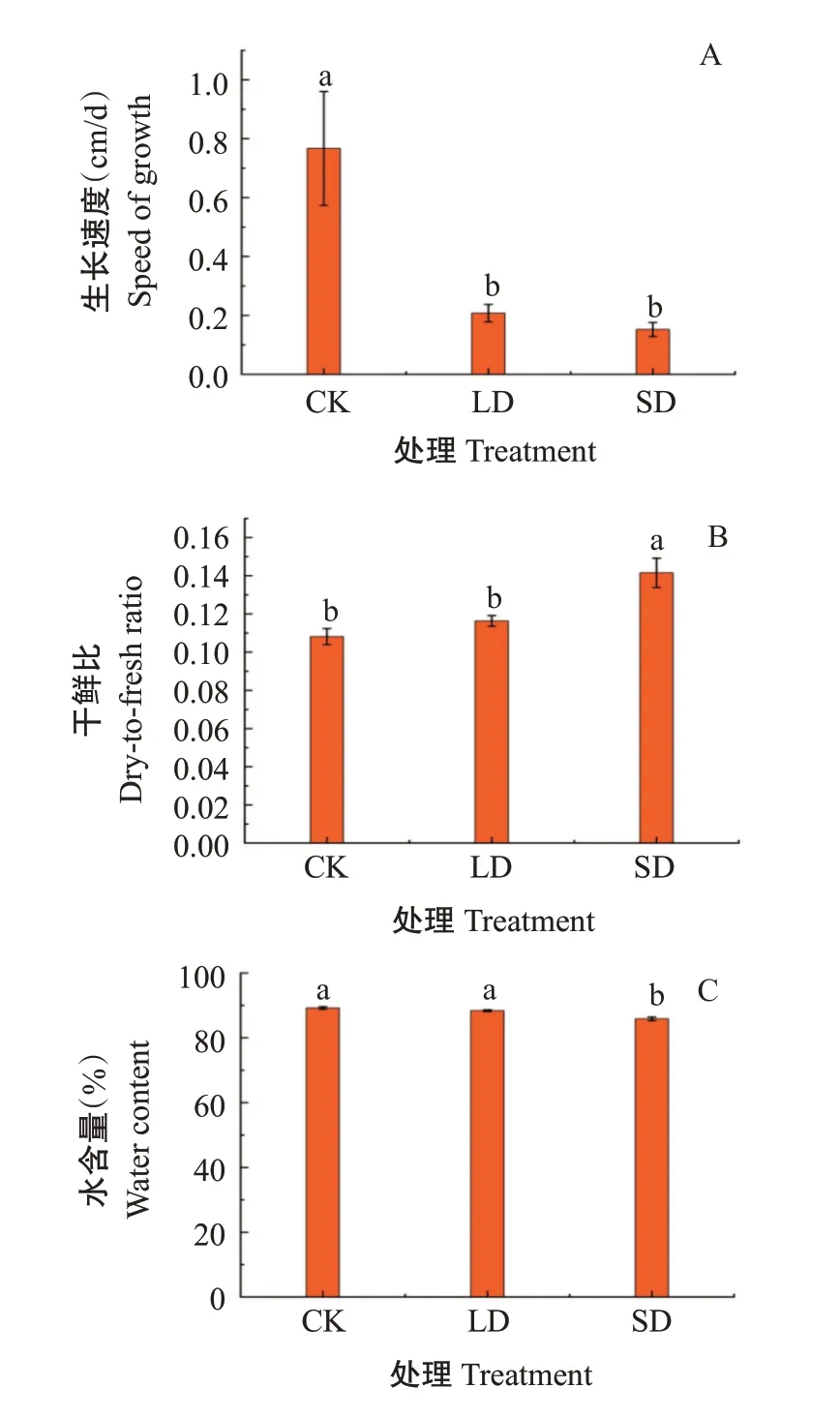
图1 低温胁迫对葛藤幼苗生长速度、干鲜比、水含量的影响Fig.1 Effects of low temperature stress on the growth rate,dryto-fresh ratio and water content of P.lobata seedlings
2.1.2 叶形态 由表1可知,与CK相比,葛藤幼苗在低温胁迫处理下的叶面积和叶周长均出现下降,LD处理下差异不显著(P>0.05,下同),SD处理下差异显著;低温下叶宽均显著低于CK,而叶长差异不显著。表明低温会影响葛藤的叶片形态,表现为叶片生长受抑制,叶面积、叶宽、叶周长均明显小于自然生长的叶片。

表1 低温胁迫对葛藤幼苗叶形态的影响Table 1 Effects of low temperature stress on the leaf morphology of P.lobata seedlings
2.2 低温胁迫对葛藤幼苗生理生化指标的影响
2.2.1 Pro和可溶性糖含量及相对电导率 随着低温胁迫程度的加强,葛藤幼苗叶片中Pro含量呈上升趋势(图2-A),LD 和SD 处理下分别较CK 上升14.02%和37.21%,但差异不显著。葛藤幼苗叶片可溶性糖含量(图2-B)和相对电导率(图2-C)均随低温胁迫程度的加强呈上升趋势,SD处理下分别较CK提高144.65%和279.21%,差异达显著水平。
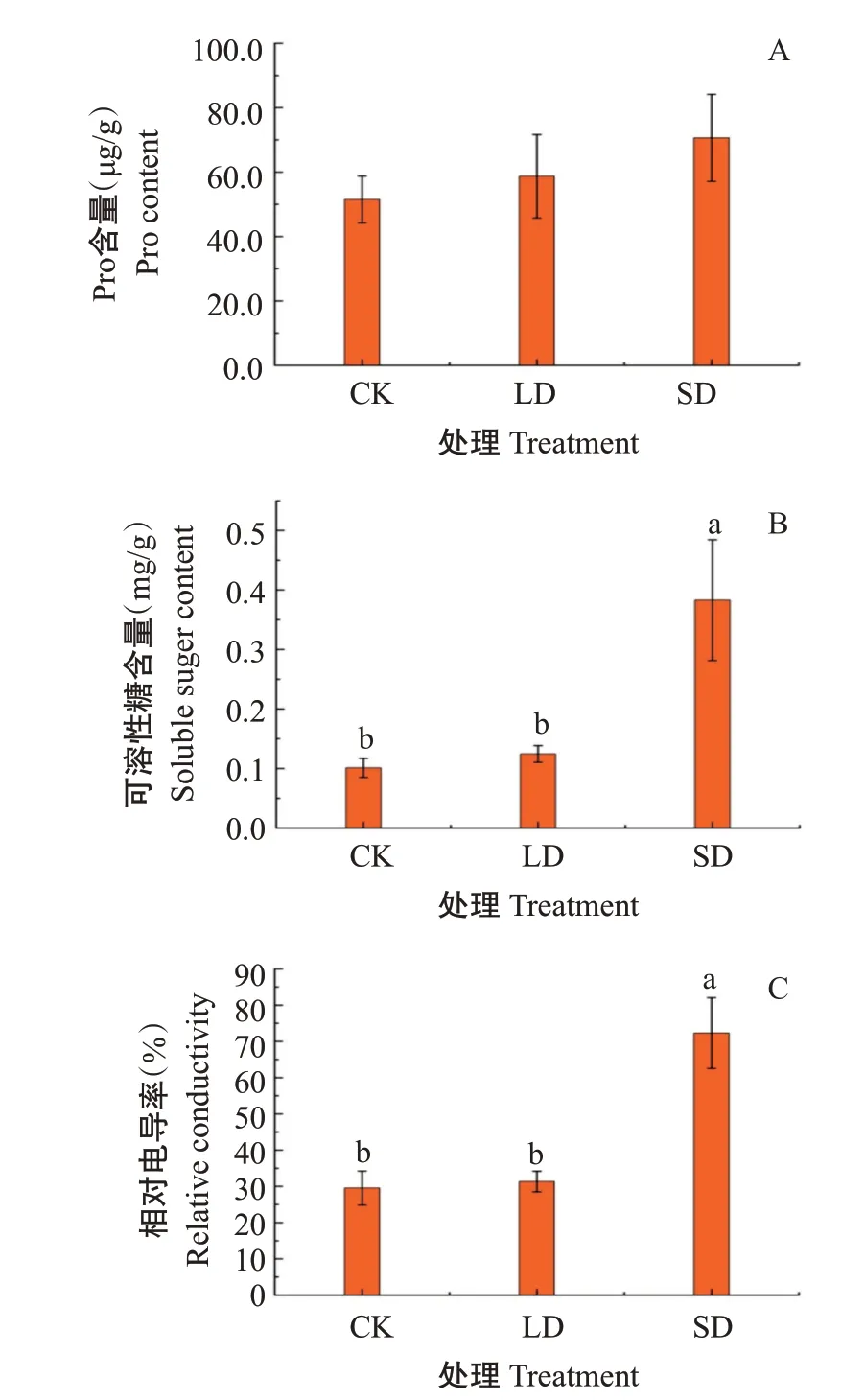
图2 低温胁迫对葛藤叶片Pro和可溶性糖含量及相对电导率的影响Fig.2 Effects of low temperature stress on the content of Pro,soluble sugar and relative conductivity of P. lobata leaves
2.2.3 MDA含量及保护酶活性 由图3-A可知,与CK相比,LD和SD处理下,葛藤幼苗叶片的MDA含量有所上升,但差异不显著。低温胁迫下各保护酶活性变化趋势不同,相对于CK,葛藤叶片中SOD(图3-B)、POD(图3-C)和APX(图3-E)活性均呈上升趋势,SD处理下分别显著提高41.32%、178.22%和74.67%;LD和SD处理下叶片CAT(图3-D)活性较CK有所下降,分别降低14.29%和24.33%,但差异不显著。
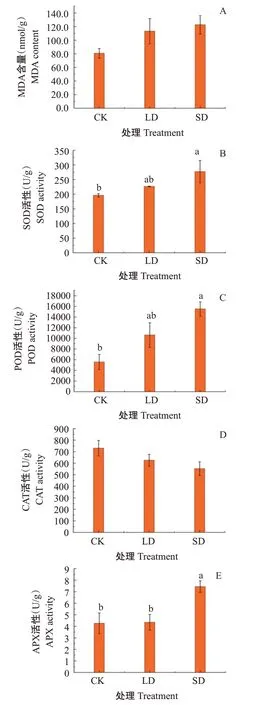
图3 低温胁迫对葛藤叶片MDA含量和保护酶活性的影响Fig.3 Effects of low temperature stress on MDA content and protective enzyme activity in P.lobata leaves
2.3 低温胁迫对葛藤幼苗内源激素的影响
随着低温胁迫程度的加强,葛藤幼苗叶片ABA含量(图4-A)、GA3含量(图4-C)、ZT含量(图4-D)呈先上升后下降的变化趋势,均在LD处理时达最大值,ABA含量显著增加17.46%,GA3和ZT含量分别增加3.31%和4.09%,但差异不显著;在SD处理下,ABA含量较CK显著降低25.40%,GA3和ZT含量分别降低5.67%和4.82%,差异不显著。葛藤幼苗叶片的IAA含量(图4-B)为CK最高,LD和SD处理下,IAA含量分别下降7.04%和21.38%,但与CK无显著差异。
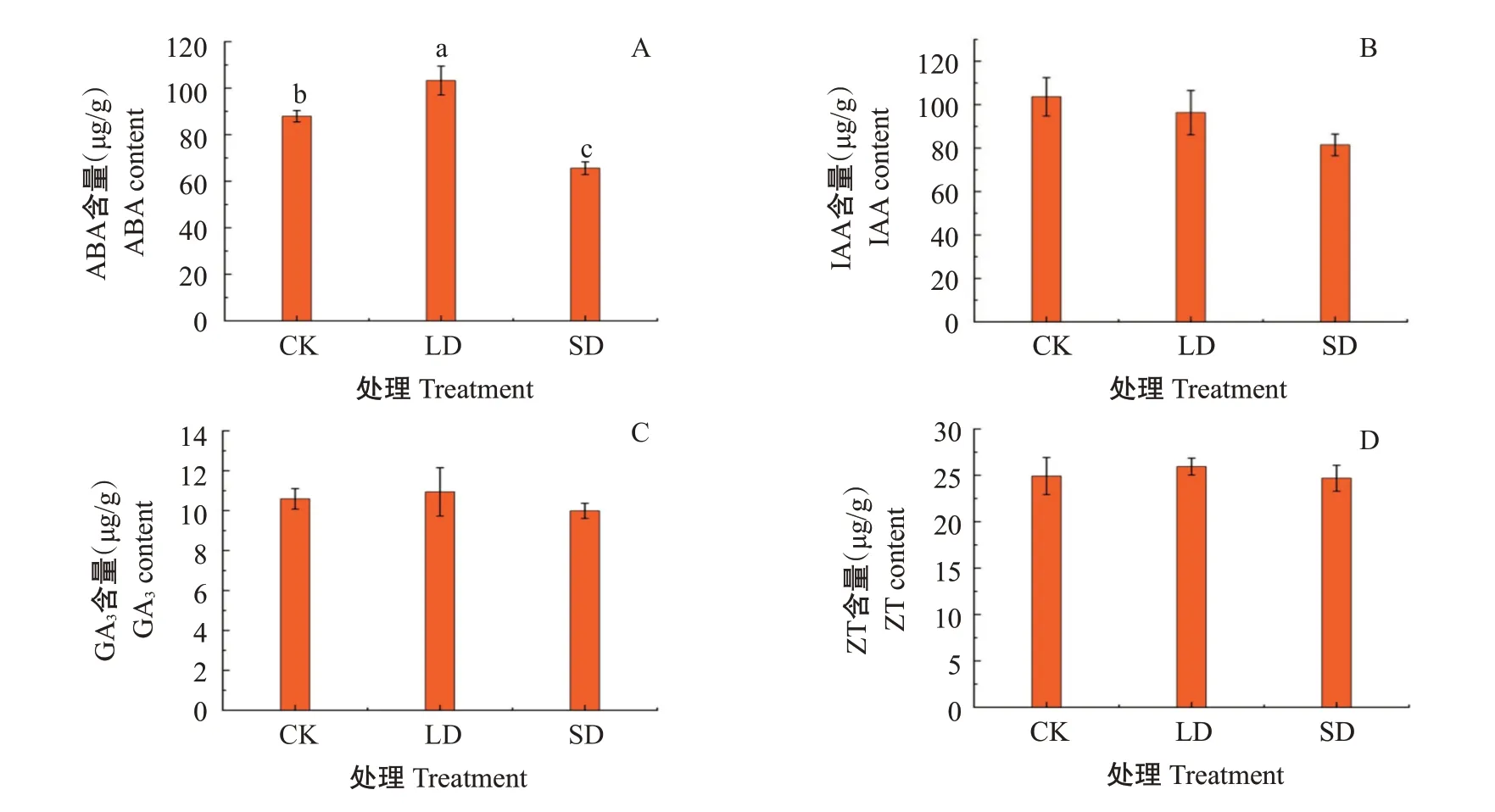
图4 低温胁迫对葛藤叶片内源激素含量的影响Fig.4 Effects of low temperature stress on the content of endogenous hormones in P.lobata leaves
2.4 低温胁迫下葛藤幼苗生长发育及生理生化指标的相关分析
对葛藤幼苗的形态指标和生理指标进行相关分析,结果(图5)表明,各性状间存在不同程度的相关性。其中生长速度与MDA含量呈显著负相关;干鲜比与相对电导率和可溶性糖含量呈显著正相关,与叶面积、叶长、叶宽及叶周长等指标呈显著负相关;ABA含量与可溶性糖含量和相对电导率分别呈显著和极显著(P<0.01,下同)负相关;Pro含量与APX活性呈极显著正相关;POD活性与IAA含量呈显著正相关。
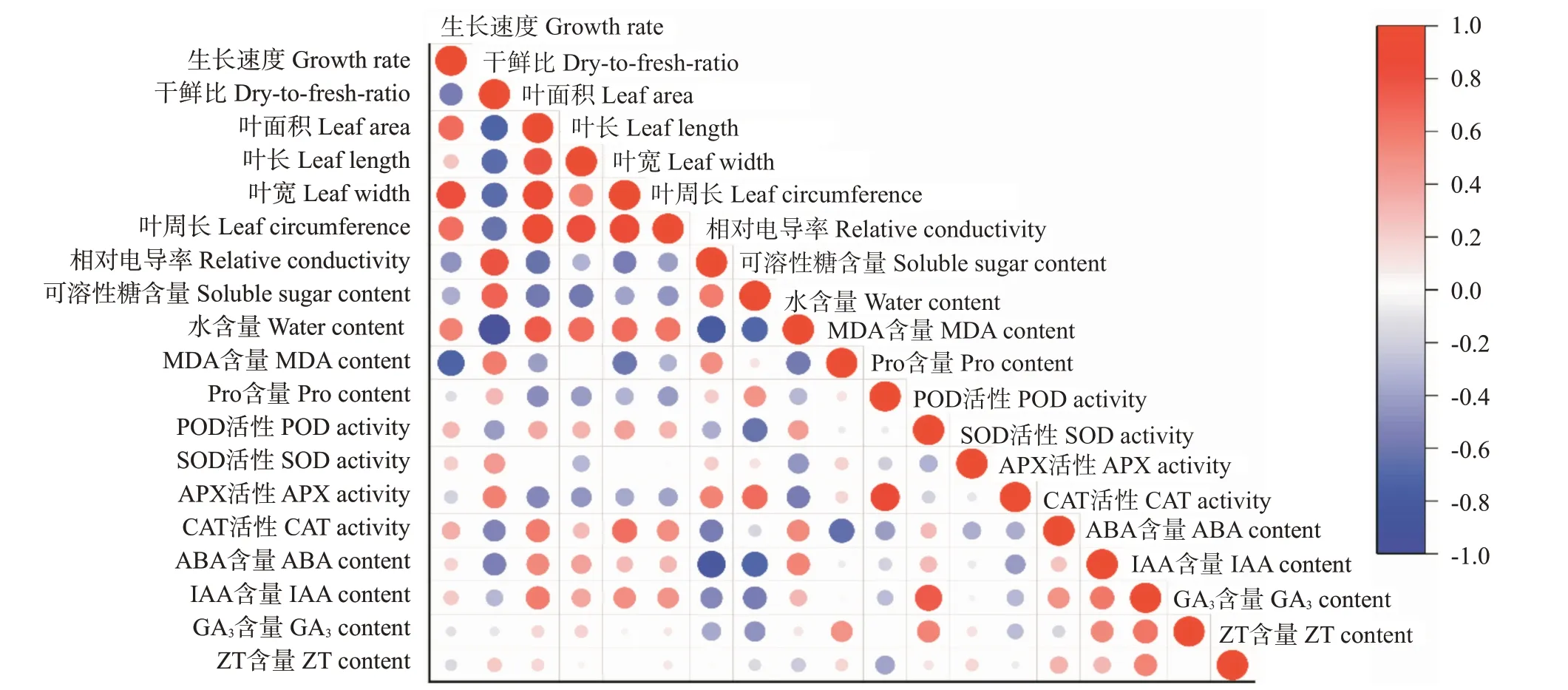
图5 低温胁迫下葛藤幼苗19个性状的相关分析Fig.5 Correlation analysis of 19 traits of P.lobata seedlings under low temperature stress
2.5 低温胁迫下葛藤幼苗生长发育及生理生化指标的主成分分析
综合低温下葛藤幼苗生长发育及生理生化2组指标,根据主成分分析(表2)中特征值大于1的标准选取6个主成分,第一~六个主成分贡献率分别为45.076%、15.050%、10.463%、8.066%、7.494%、6.714%,累积贡献率达92.863%,具有较好的代表性,其余可忽略不计。
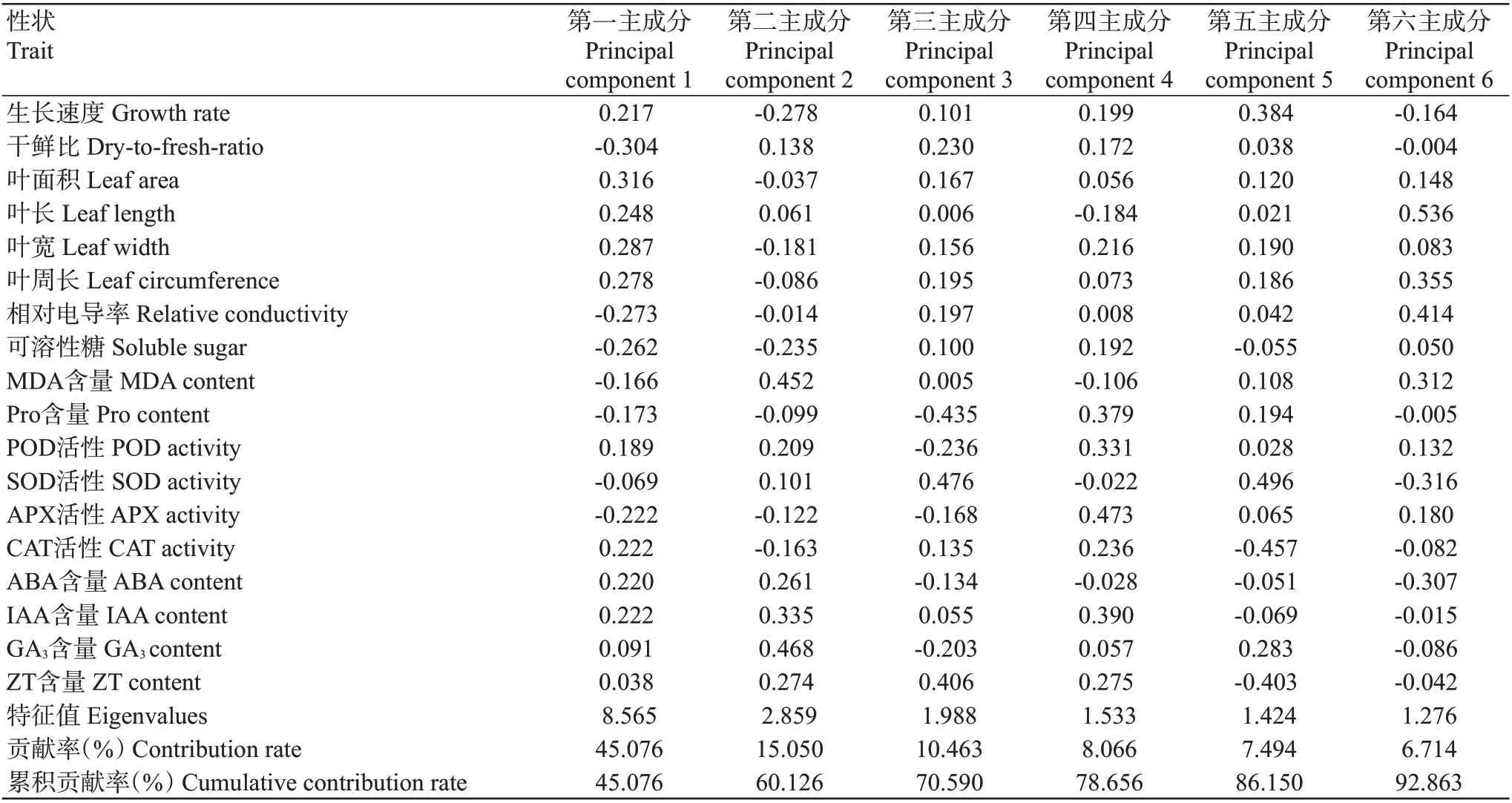
表2 低温胁迫下葛藤幼苗18个性状的主成分分析Table 2 Principal component analysis of 18 traits of P.lobata seedlings under low temperature stress
第一主成分中贡献率较大的是干鲜比、叶面积、叶宽,低温胁迫下葛藤生长发育减缓,叶片生长受到一定影响;第二主成分中贡献率较大的是GA3和MDA含量,低温下植物膜脂过氧化会造成植物体内MDA积累;第三主成分中SOD活性、Pro和ZT含量贡献率较大,细胞内抗氧化酶活性及膜保护物质含量越高,幼苗受到的伤害程度就越小;第四主成分中贡献率较大的是APX活性和IAA含量,其中IAA是生长促进类激素,植物通过降低体内IAA含量,抑制植物生长,增强植物的抗冷性;第五主成分中SOD和CAT活性贡献率较大,寒冷条件下细胞内保护酶活性的变化反映了葛藤幼苗的抗寒能力;第六主成分中贡献率较大的是叶长和相对电导率。
2.6 低温胁迫下葛藤幼苗形态及生理生化指标的综合性评价
对葛藤的性状经标准化处理后,将不同温度下葛藤各主成分得分分别与所对应的贡献率相乘,最终求出综合得分,以此来对抗冷性进行综合评价。前6个主成分可有效表现不同温度下葛藤的生长情况,其综合得分情况如表3所示。葛藤抗冷能力的综合评价值D值随着胁迫强度的上升而减少,重度低温胁迫(SD)下葛藤幼苗叶片的D值与轻度低温胁迫(LD)下D值的平均值相差不大,LD2处理下D值最小,隶属函数分析表明低温会对葛藤的生长带来一定的负面影响。
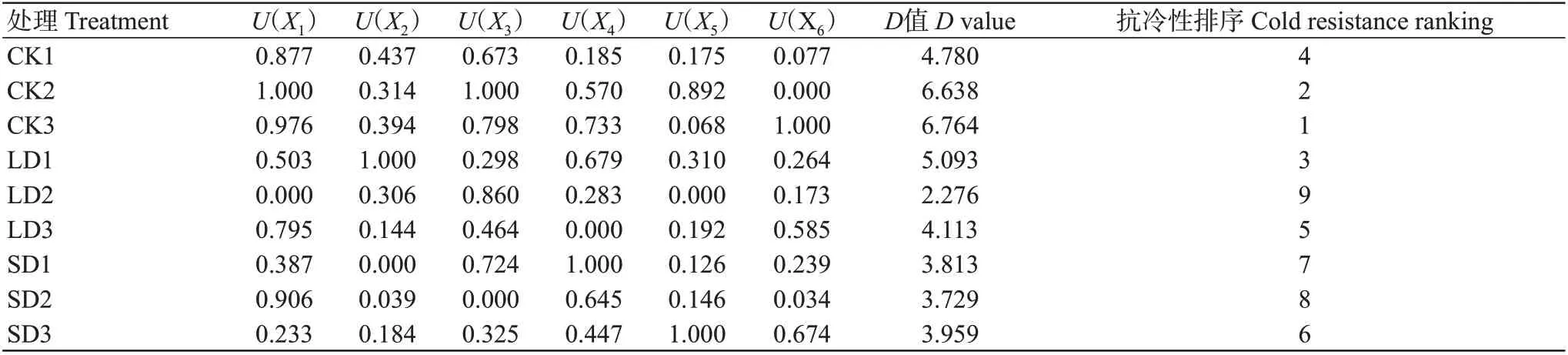
表3 葛藤幼苗各综合指标的隶属函数值、D值及综合评价Table 3 Membership function value,D value and comprehensive evaluation of each comprehensive index of P.lobata seedlings
3 讨论
3.1 低温胁迫对葛藤幼苗生长的影响
温度是影响植物生长发育的重要环境因子之一,在低温下植物的生长会受到严重抑制。本研究中,随着温度的降低,葛藤幼苗生长速度不断下降,且这种受抑制的情况在叶片上也存在,叶面积、叶宽、叶周长及水含量与CK相比均出现显著下降,且温度越低,各生长指标的值越低,与欧阳孔胜等(2011)对低温处理甜瓜砧木的研究结果相同。植物细胞中含有大量水分,充足的水分使得植物在生长中枝叶挺立,便于充分接受光照和交换气体;同时,水分还是植物对物质吸收及转运的溶剂。水分的减少会限制各种物质在植物体内的运输,从而导致植株的光合和代谢作用大大减弱,叶片内合成光合产物的原料减少则会使植株营养物质的合成减少,使得植物生长发育减缓。本研究中,葛藤叶片干鲜比随温度下降呈上升趋势,通过观察叶片,发现有萎焉、收缩现象。表明低温会引起葛藤叶片水分含量下降,其水分的利用受到一定限制,叶片细胞膨压不足,不能保持叶片固有姿态,从而发生改变,植株的生长发育也因低温受到一定程度的抑制。
3.2 低温胁迫对葛藤幼苗生理生化的影响
低温环境下植物细胞的渗透势可通过可溶性物质的积累来调节,从而缓解逆境对膜系统的伤害(侯祯丹等,2019)。可溶性糖和游离脯氨酸是细胞质重要的渗透调节物质。植物体内的可溶性糖积累可提高细胞液浓度,从而降低水势及冰点(王卓敏等,2016)。本研究中,葛藤幼苗叶片中Pro和可溶性糖含量均随温度降低呈上升趋势,与CK相比在SD处理下叶片中的可溶性糖含量显著上升279.21%。与前人对番茄(王孝宣等,1998)、西番莲(董万鹏等,2015)及棉花(王晓曼等,2020)的研究结果相同,表明葛藤会通过增加体内可溶性糖和Pro的积累,增强自身的保水能力,维持自身渗透势和膜的正常功能,避免细胞遇冷凝固,防止细胞损伤,从而抵抗低温胁迫。
基于电解质外渗量测定开发的电导法是在植物上应用较广泛的抗寒性鉴定方法(王晓辉等,2012),在拟南芥低温抗性鉴定中普遍采用存活率和相对电导率作为衡量指标,低温下存活率高、相对电导率低则表明低温抗性能力强。本研究中,葛藤叶片相对电导率呈上升趋势,但LD处理下与CK差异不明显,SD处理时显著上升。在不同的处理温度下,抗寒性强的品种相对电导率较低,抗寒性弱的品种相对电导率较高(王召元等,2022)。本研究中葛藤在LD处理下电导率与对照组差异不显著的结果与前人对谷子(张彬等,2020)、卫矛属植物(杨航宇和刘长仲,2020)的研究结果相同,表明葛藤在LD处理下具有较好的抗寒性,但具体适宜温度还有待于进一步验证。
植物遭受低温时,细胞内的自由基产生、清除都会受到影响,严重时过氧化自由基代谢平衡会被破坏,自由基过多的积累则会导致膜系统受到损伤,造成膜脂过氧化(洪舟等,2020)。膜脂过氧化会使细胞膜渗透性变大,细胞内的MDA大量积累,进而影响植物体内多种正常反应。本研究中,LD处理下葛藤叶片的MDA含量有所上升但差异不显著,与前人对番茄(高茜等,2020)、紫花苜蓿(陈胜萍等,2017)进行低温处理的研究结果相似,表明在低温胁迫下葛藤幼苗膜脂过氧化程度低,受氧化损伤程度较弱,低温未对葛藤造成明显的膜脂过氧化损伤,说明葛藤具有较强的抗寒能力。
本研究中,葛藤幼苗叶片的SOD、POD、APX活性均随温度的降低而升高,低温胁迫下葛藤叶片中CAT活性虽然表现为下降趋势,但与CK差异并不显著,CAT活性始终保持在较高水平。SOD是植物重要的保护酶系统,同时也是保护酶系统的第一道防线,能将毒性较强的转化为毒性较弱的H2O2(高茜等,2020);但H2O2过量累积仍会对细胞造成伤害,所以转化的H2O2需通过CAT和POD等其他抗氧化酶清除。本研究中葛藤幼苗叶片的CAT活性呈下降趋势,会使H2O2清除能力降低,可能造成H2O2的积累,使得膜系统遭受活性氧物质积累的伤害,但葛藤在低温下POD活性与CK相比有明显上升,说明POD清除过氧化氢的调控机制已形成,能在一定程度上帮助葛藤幼苗调节细胞膜透性,防止膜受损伤,从而提高葛藤幼苗的低温适应能力,与周福平等(2020)对高粱的研究结果相同。根据研究结果及葛藤接受低温处理后的生长发育情况来看,葛藤幼苗并未出现严重损伤及死亡,表明可能是低温下葛藤幼苗叶片的SOD、POD、CAT和APX等保护酶的综合耐低温机制已形成,活性氧等各种自由基积累减少,细胞内活性氧代谢处于一定的平衡状态,膜脂过氧化作用得到缓解,降低了细胞膜的破坏程度,即葛藤可通过调整自身的保护酶系统来增强其耐低温性。
3.3 低温胁迫对葛藤幼苗内源激素的影响
植物激素是由植物体自身代谢产生的一类有机活性化合物,数量很少,自产生部位运输到作用部位,在很低浓度时就能发挥显著生理效应(丁广文等,2016)。本研究中,随着温度下降,葛藤幼苗叶片的ABA、GA3和ZT含量先上升后下降,IAA含量不断下降。与周锦业等(2020)的研究结果相似,可能是由于低温胁迫引发植物体内渗透调节的改变,葛藤为抵抗低温带来的伤害从而增加抑制类内源激素如ABA的含量,以增强自身耐低温性。而GA3和IAA有加速细胞分裂、促进成熟细胞伸长的作用(袁梦佳等,2021),被认为是生长促进类激素。GA3还可通过影响植物体内自由水和束缚水的含量来改变其抗冷性(许耀照等,2020)。葛藤在低温下通过降低体内IAA和GA3含量,抑制植物生长,增强了自身的抗冷性。本研究结果表明葛藤幼苗为抵御低温胁迫带来的不利影响,通过增加体内生长抑制类激素,降低生长促进类激素的含量,以此来增强自身的抗冷性。
3.4 低温胁迫对葛藤幼苗生长发育及生理生化变化的综合评价
对葛藤幼苗的形态指标和生理指标进行相关分析,发现生长速度与MDA含量呈显著负相关。植物在遭遇低温胁迫时,膜脂过氧化产物MDA会大量积累(蒋安等,2010),MDA积累会对细胞,特别是细胞膜造成一定伤害。本研究结果表明,当葛藤幼苗遭受低温胁迫时,MDA含量不断上升,对植株生长造成了不良影响。干鲜比与相对电导率和可溶性糖含量呈显著正相关,说明植物水分的减少和干物质的积累会对相对电导率上升起重要作用,可溶性糖含量增加与细胞渗透势调节密切相关。通过对各个材料主成分值进行比较,可了解不同样本间的差异(于凤瑶等,2010)。本研究对葛藤生长性状的主成分分析发现,前6个主成分对表型变异的累计贡献率达92.863 %。根据各主成分的特性,可将6个主成分分为三大类,第一类是产量性状,比如第1主成分中的生长速度、叶面积和周长等,主要反映低温胁迫下葛藤生长发育变化,在低温胁迫下葛藤产量性状呈下降趋势;第二类是保护酶活性,如主成分中贡献率较大的MDA、SOD、APX、CAT活性,细胞内抗氧化酶活性及膜保护物质含量越高,植物的抗冷性越强,说明幼苗受到的伤害程度越小,与冯昌军等(2005)对苜蓿进行低温胁迫的研究结果相似;第三类是内源激素,如IAA、GA3等,内源激素在主成分累计贡献率占比较高,说明内源激素对植物抗冷性调节具有重要意义。
结合形态、生理指标及隶属函数进行分析,低温处理下隶属函数值呈下降趋势,叶片也出现轻微萎焉、收缩,说明低温对葛藤幼苗的生长产生了一定的胁迫作用;最低的函数值出现在LD处理下,而并非在SD处理,对比2个处理下隶属函数平均值,分别为3.82和3.83,并无明显差异。可溶性糖等渗透调节物质含量上升,SOD、POD等保护酶活性上升,MDA等膜脂过氧化物质积累较少,说明葛藤具有一定的抗冷性。综合上述分析,虽然低温会对葛藤生长造成影响,但遭遇低温时,葛藤会通过减缓自身生长、降低水含量、提高保护酶活性等一系列措施来抵御低温带来的不利影响,抗冷性较强,具有在低温下存活的能力。但本研究只是基于植物生理和激素水平对葛藤的耐低温性进行探讨,今后还应结合不同产地、不同生长阶段、不同低温胁迫时间对植株生长性状的差异和当地地理环境进行综合探讨与研究,以期选育出抗冷性强、适合在西南地区栽培的葛藤品种。
4 结论
葛藤幼苗在低温下MDA增加不明显,可溶性糖含量和各种保护酶活性升高,生长促进类激素含量降低,生长抑制类激素含量增加。低温下葛藤生长减缓,但并未出现明显损伤,表明葛藤幼苗可通过增强渗透调节能力、提高保护酶活性、调节激素含量来提高自身抗冷性,进而抵御低温胁迫带来的不利影响。
