福建省鼠感染钩端螺旋体调查和分离株基因种的鉴定
2022-03-18徐国英潘敏楠刘维俊林代华韩腾伟周淑姮王加熊肖方震
徐国英,潘敏楠,刘维俊,林代华,韩腾伟,周淑姮,王加熊,刘 菁,肖方震
钩端螺旋体病是由问号钩端螺旋体(简称钩体)引起的一种常见的人兽共患传染病。鼠类是钩体重要宿主之一,钩体栖息在宿主的肾小管中,随尿液排出到环境中。人通过参加农业劳动、接触疫水或参加抗洪抢险及涉水,间接接触污染水源,环境中的钩体通过皮肤、黏膜进入机体而感染。福建省曾是我国钩体病重点流行地区,疫区分布广泛,全省各县市均有病例报告。20世纪曾发生过较大流行,自1992年起,虽疫情较为平稳,但仍有局部暴发流行。近年福建省疫报病例数居全国前列,关于福建省钩体病的基因组学及分子流行病学特征尚未见详细报道。本研究根据2015-2019年福建省钩体病疫情选取14个调查点进行鼠类钩端螺旋体的调查和监测,了解福建省不同地区鼠中钩体的感染状况,同时对从宿主动物鼠类监测分离的23株钩体分离株,扩增菌株的16SrRNA基因并对其进行基因种鉴定,初步探讨福建省钩体基因种的特征,为福建省钩体病预防控制提供科学依据。
1 材料与方法
1.1 标本采集 2015-2019年在闽东(长乐、福清、闽侯、罗源、连江和霞浦)、闽西(武平、永安、大田和泰宁)、闽南(长泰和德化)和闽北(建阳和邵武)共14个县市,采用笼夜法捕鼠,对捕获鼠进行种类、性别、成幼等鉴定,计算鼠捕获率和鼠种组成,采集鼠肾、鼠心血等进行分离培养,采集鼠血清检测钩体抗体。
1.2 菌株来源 本研究选取福建省2005年以来从啮齿类动物监测中分离到的23株钩体分离株,分别来自漳州(长泰9株)、龙岩(武平7株)、福州(长乐1株、福清1株、闽侯1株和罗源1株)、和南平(建阳2株)、三明(将乐1株)共5个地区,菌株具体信息见表1。
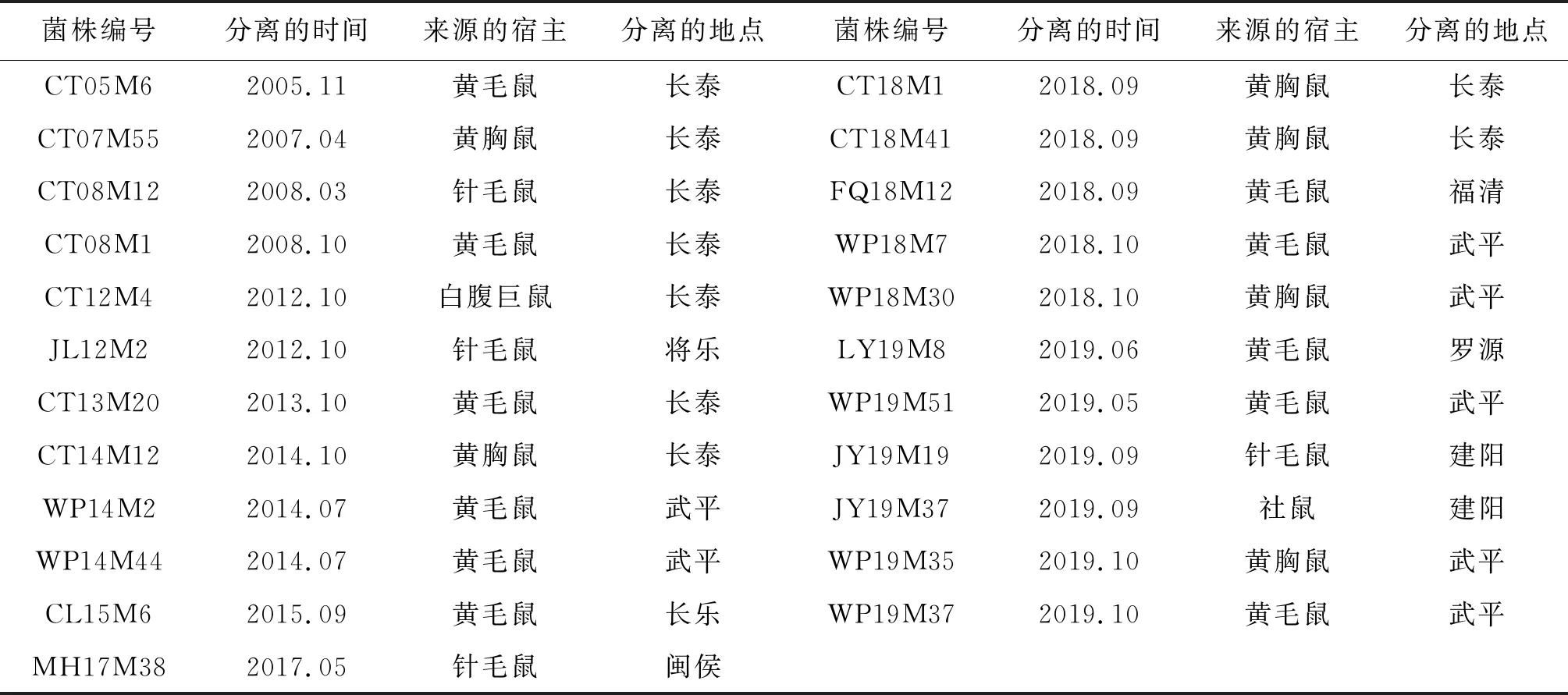
表1 福建省钩端螺旋体分离株基本信息
1.3 主要试剂 EMJH钩体培养基由美国BD公司生产,PCR Premix 和2 000bp DNA Marker(购自 TaKaRa 公司),基因组提取采用QIAamp DNA Mini Kit(购自QIAGEN公司)。
1.4 血清群鉴定 采用暗视野显微镜凝集试验(Microscopic agglutination test, MAT),用15群15型钩体参考菌株诊断血清(来源于中国食品药品检定研究所)进行血清凝集试验,在暗视野显微镜下观察,以出现50%菌体凝集菌群为最终鉴定血清群。
1.5 分离菌株基因组提取 分离菌株在EMJH钩体液态培养基中28 ℃培养7~10 d,达到对数生长期提取基因组DNA,操作方法按试剂盒说明书进行,提取的DNA保存于-20 ℃冰箱中。
1.6 16SrRNA基因PCR扩增 参照参考文献[1]报道的引物序列进行PCR扩增,引物序列如下: fD1: 5′-CCG AAT TCG TCG ACA ACA GAG TTT GAT CCT GGC TCAG -3′rP2: 5′-CCC GGG ATC CAA GCT TAC GGC TAC CTT GTT ACG ACTT-3′引物由上海生工生物工程技术服务有限公司合成。采用50 μL PCR反应体系扩增, Premix Taq 25 μL,DNA 模板 2 μL,上下游引物各2 μL(10 μmol/L),用双蒸水补足至50 μL。PCR 反应条件:94 ℃ 5 min,94 ℃ 15 s,50 ℃ 5 s,72 ℃ 90 s,共设35个循环,72 ℃延伸5 min。经1.2% 琼脂糖凝胶在电压100 V条件下电泳40 min后,在凝胶成像仪中观察目标条带。
1.7 核酸序列测定及分析 扩增产物送上海生工生物工程股份有限公司进行双向测序,测序得到的DNA片段序列通过美国国家生物技术信息中心(NCBI)BLAST进行同源性分析,同时应用MEGA6.0软件,并以GenBank中已知的17种钩体菌株基因序列作为参照,以邻接法(Neighbor -Joining)构建系统进化树,采用Bootstrap验证(n=1000)。
2 结 果
2.1 鼠捕获率和鼠种构成 共布放鼠笼18 040笼次,捕获鼠类1 221只,鼠平均捕获率为6.77只/100笼;鼠种构成以黄毛鼠、黄胸鼠、针毛鼠及褐家鼠为主,分别占40.87%(499/1 221)、19.57%(239/1 221)、16.79%(205/1 221)及8.85%(108/1 221)。闽东鼠捕获率最高,为8.95只/100笼,其次是闽西,为6.46只/100笼,闽北最低为5.97只/100笼(表2)。
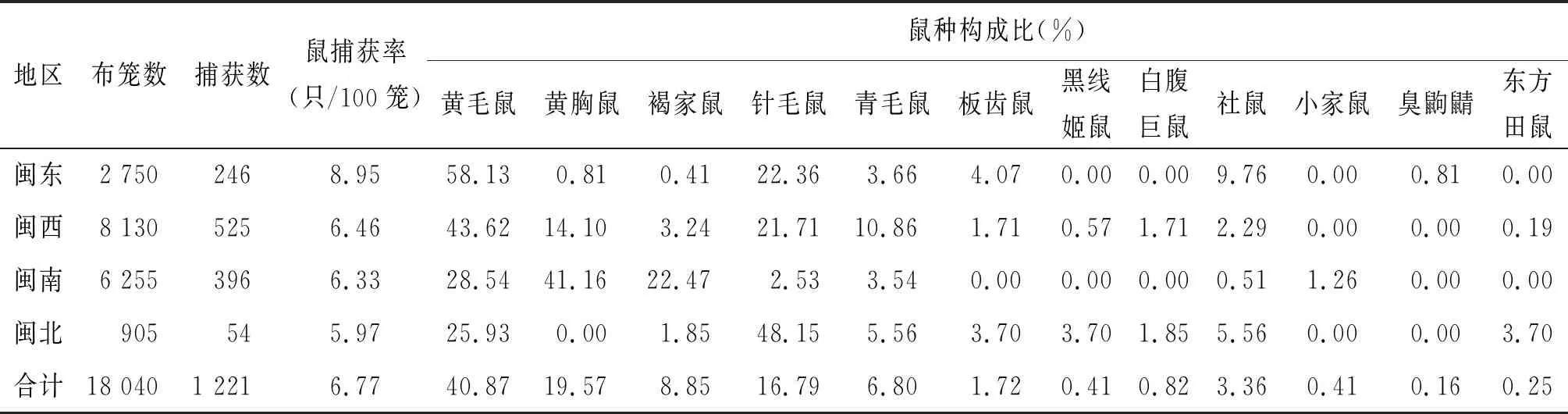
表2 福建省鼠捕获率及鼠种构成
2.2 不同鼠种钩体感染状况 捕获181 221只鼠中有706只鼠采集心血,其中207只鼠血清感染钩体,阳性率为29.32%。其中黄毛鼠、褐家鼠和青毛鼠感染率较高,依次为42.16%(121/287)、26.00%(13/50)、22.22%(14/63)。不同鼠种的血清钩体感染率差异有统计学意义(Fisher检验,P<0.05)(表3)。
2.3 不同地区钩体感染状况 207只感染钩体的鼠中血主要感染菌群为爪哇群和秋季群,分别占53.62%(111/207)和20.77%(41/207)。表3显示,不同地区鼠感染率的差异无统计学意义(χ2=0.55,P=0.91>0.05)。
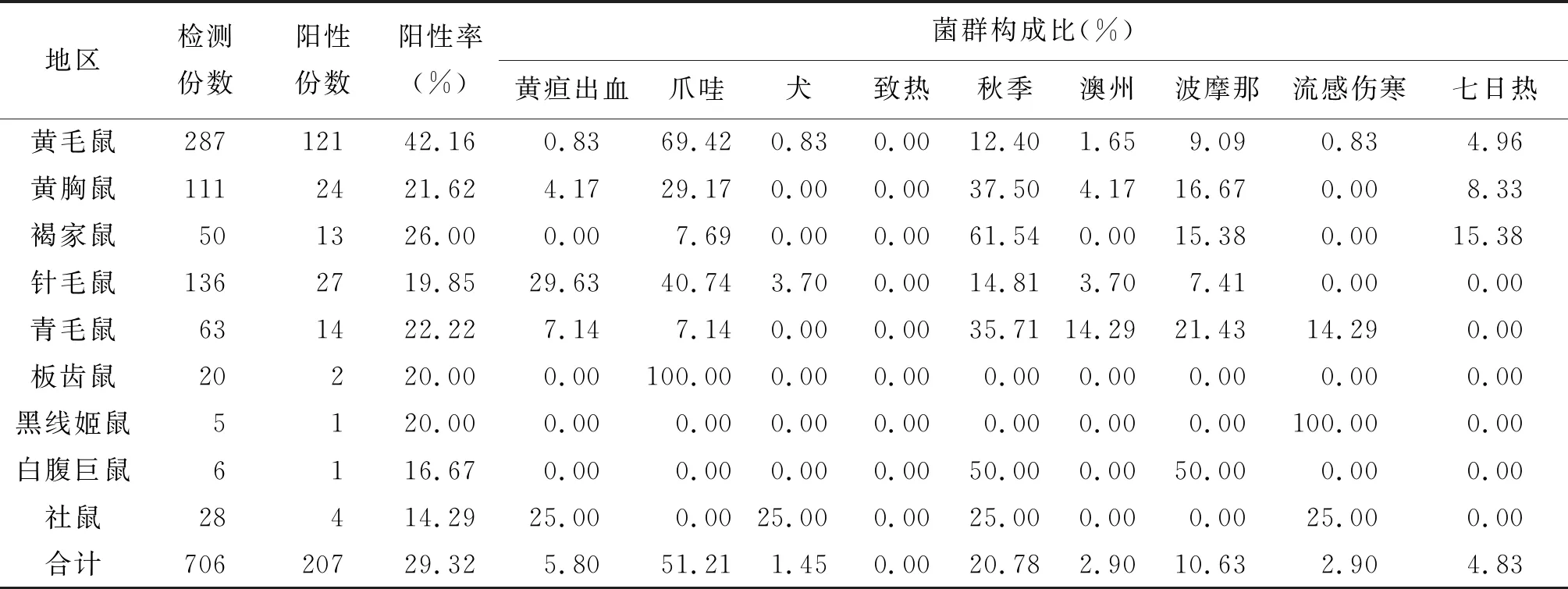
表3 福建省不同鼠种血清钩体抗体检测及菌群分布
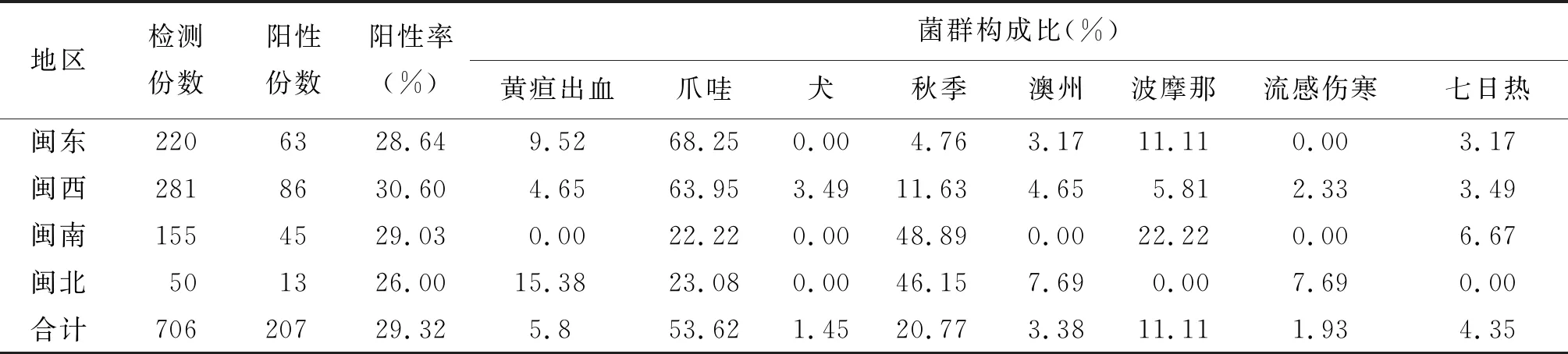
表4 福建省不同地区鼠血清钩体抗体检测及菌群分布
2.4 PCR扩增的结果 23株钩体分离株扩增16SrRNA基因,凝胶电泳显示,条带处于约1 484 bp位置,与预期目标条带一致(见图1)。

M为DL2000 DNA Marker;1~12、14~24为分离菌株;25为阳性对照(黄疸出血群);13、26为阴性对照。
2.5 基因种的鉴定 23株钩体分离株的16SrRNA基因测序结果与GenBank数据库收录的钩体16SrRNA基因序列比对,同源性>99.7%认为是同一个基因种,结果显示:23株菌株中属于致病性的有13株,其中隶属于L.borgpetersenii基因种有8株,分别是编号CL15M6、MH17M38、WP18M7、WP18M30、WP19M35、WP19M51、WP19M37、LY19M8,血清群均为爪哇群。属于L.interrogans基因种有5株,分别是CT18M1、CT18M41、FQ18M12、JY19M37、JY19M19,其中CT18M41、FQ18M12两株血清群鉴定为巴达维亚群,CT18M1为致热群,其余2株为黄疸出血群。剩余10株属于非致病的L.meyeri基因种,编号分别是CT05M6、CT07M55、CT08M1、CT08M12、CT12M4、CT13M20、CT14M12、JL12M2、WP14M2、WP14M44,血清群鉴定为不凝集。
2.6 序列分析结果 采用MEGA6.0软件,选择邻接法建树(Bootstrap=1000),结果显示:福建省鼠中分离保存的23株钩端螺旋体,16SrRNA基因与L.borgpetersenii(AY887899)基因种序列处于同一分支的有8株,分别是来自闽东的3株即长乐区、闽侯区、罗源县(各1株)、闽西的武平县(5株),与L.interrogans(AY996791)基因种序列在同一分支的有5株,分别是闽南2株即长泰县(2株)、闽东福清县(1株)以及闽北建阳区(2株),与非致病的L.meyeri(AY631889)基因种处于同一分支的有10株钩体菌株,分别来自闽南长泰县(7株)、闽西武平县(2株)及将乐县(1株)。见图2。

图2 钩端螺旋体16S rRNA基因系统发生树
3 讨 论
鼠类作为钩体的重要储存宿主,在钩体病的传播、流行中起着重要作用。福建省是钩体病的老疫区,地理环境复杂,气候湿润,啮齿类动物丰富,感染钩体菌群众多[2]。目前,国际上对钩端螺旋体的分类方法主要有血清学分类和基因分类2种[3]。钩体菌群菌型复杂,在全球范围内,至少包括24个血清群300多个血清型,基因种至少也有18个。传统血清学分型方法需要保存大量的标准菌株及多种抗血清,存在操作繁琐,费时费力且难以实现标准化等不足之处,不能反映菌株之间基因上的联系,而基因分类是通过比较基因DNA片段核苷酸序列同源性,继而从遗传学和分子生物学水平上进一步阐明不同钩体菌株的差别。细菌的16SrRNA具有高度的保守性已被广泛用于钩体基因种鉴定、分子进化的分析指标[4]。
本研究根据2015-2019年福建省钩体病疫情选取14个调查点进行鼠类钩端螺旋体的调查和监测,发现福建省每个调查点的鼠类均有钩体的感染,不同鼠种血清均有钩体抗体检出,但优势鼠种黄毛鼠、针毛鼠、黄胸鼠、褐家鼠感染率位于前列,尤以黄毛鼠最高,与2011-2016年福建省[2]的调查结果基本一致,与赵明惠[5]调查江西口岸及国检监管区鼠类的状况相似,也是从褐家鼠、社鼠、黄毛鼠及黄胸鼠中检测出钩体的感染。不同地区鼠类钩体感染率差异无统计学意义,与2011-2016年福建省[2]的调查结果即闽北最高不吻合,可能与此次闽北采样数较少,不能较全面反映该地状况有关。
本研究还对自2005年以来在钩体病啮齿动物监测中分离到且在液体培养基传代保存的23株钩体菌株进行基因种鉴定和序列分析,发现福建省鼠类的钩体基因种以致病性为主,也存在非致病性基因种。同时发现基因种的分类与血清群的分类存在着交叉的现象,即致病性基因种含有爪哇群,非致病性基因种也有爪哇群,与严杰[3]描述的情况(不同基因种的钩体可表达同一血清型特异抗原)一致。与其它省份的钩体基因种有所不同,如贵州省李世军从黑线姬鼠分离的3株菌株均为L.interrogans基因种[6-7],张翠彩[8]对江西省的2013年以来啮齿类动物监测中分离到的27株钩体分离株进行基因种的鉴定,发现江西省菌株隶属于L.interrogans和L.borgpetersenii2个致病性基因种,其中L.interrogans为江西省主要优势基因种,占77. 78%(21/27)。Florence等[9]对法国里昂城市的鼠类进行钩体检测,发现L.interrogans为主要基因种,Benacer等[10]对马来西亚半岛的城市鼠类进行钩体检测,研究发现:L.interrogans(59%,23/39)和L.borgpetersenii(41%,16/39)2个致病性基因种为主要基因种,Voronina等[11]对俄罗斯29株钩体分离株进行分析,发现该地区存在L.interrogans、L.kirschneri和L.borgpetersenii3个基因种。综上,国内及国际鼠类携带的钩体常见致病基因种为L.interrogans和L.borgpetersenii基因种,以L.interrogans基因种为主,但不同地区和国家还存在一些特定的基因种,比如我省钩体菌株除致病性基因种外,还存在非致病性基因种10株,且这10株均是2014年及以前分离的菌株,不知是在钩体液体培养基传代保存时间较长,使其抗原性及致病性发生改变,或是某些基因片段发生变化所致,有待今后继续研究。
研究发现我省优势鼠种携带致病性基因种钩体。这13株致病性基因种钩体菌株分别是从黄毛鼠(6株)、黄胸鼠(4株)、针毛鼠(2株)、社鼠(1株)分离到,10株非致病性基因种从黄毛鼠分离到5株,黄胸鼠分离到2株,针毛鼠分离到2株,白腹巨鼠分离到1株,说明我省优势鼠种黄毛鼠、黄胸鼠、针毛鼠和白腹巨鼠均携带有钩体。国内贵州省刘英是从黑线姬鼠分离到56株菌株[12],张翠彩[8]从啮齿类动物监测中分离到的27株钩体分离株中有16株是从黑线姬鼠分离到,9株从黄毛鼠分离得到。Florence等[9]对法国里昂城市的鼠类调查以及Benacer等[10]对马来西亚半岛的城市鼠类的调查发现是以褐家鼠为主,不同国家和地区,鼠种不同,携带的钩体基因种也会有所差别,因此需要加强监测,做好钩体病的传染源控制即灭鼠工作。
从携带钩体菌株的地点分布来看,13株致病性钩体分离地点分布在闽西的武平县5株,闽南的长泰县2株和闽北的建阳区2株,闽东的闽侯区、罗源县、长乐区、福清市各分离出1株。非致病性菌株10株分布的地点是闽南的长泰县占7株,闽西的武平县占2株,将乐县占1株,地点分布比较广泛即在我省的东部、南部、西部和北部地区的鼠种中均分离出钩体菌株,与张翠彩[7]报道的江西省鼠种钩体基因种的分布存在一定的地域差异,说明我省各地的啮齿动物均有感染钩体,应加强各地鼠种感染钩体的监测工作。
本研究也存在一定不足,其一是选取的调查监测面还不够广泛,选用的钩体分离菌株来源于监测点的不同检测年份,有些菌株距离现在相对比较久远,在液体培养基传代过程中可能出现抗原性与致病性的改变;其二是菌株数量相对比较少,今后将继续扩大样本量和不同宿主来源,对钩体分离株的致病力和遗传进化特征等进行研究。
利益冲突:无
引用本文格式:徐国英,潘敏楠,刘维俊,等. 福建省鼠感染钩端螺旋体调查和分离株基因种的鉴定[J]. 中国人兽共患病学报,2022,38(2):175-181. DOI:10.3969/j.issn.1002-2694.2022.00.016
