株、行距配置对南疆冬小麦干物质积累、转运及产量的影响
2022-03-17梁雪齐吴全忠陈国栋翟云龙
梁雪齐,张 锋,陈 猛,李 玲,吴全忠,陈国栋,翟云龙
(塔里木大学 农学院,新疆阿拉尔 843300)
小麦作为新疆第二大作物,其生产对保证边疆粮食安全至关重要。小麦产量由穗数、穗粒数以及千粒质量三因素共同制约,干物质积累是衡量作物产量的一项重要指标,在一定范围内产量与花后干物质转运量呈极显著正相关,小麦籽粒的饱满程度1/3来源于花前营养器官转移,2/3来源于花后的同化产物[1-3],大穗型品种地上部分干物质量、花后不同营养器官干物质量向籽粒的转运量极显著高于多穗型、中穗型品种[4-5]。
外界环境和栽培管理措施对小麦干物质积累、转运及产量均有较大的影响[6],通过改变农艺措施来调节源库平衡以确定合理群体结构。前人研究大多集中在耕作方式、播期、密度以及土壤肥力等方面[7-8],对超窄行密植种植冬小麦研究较少。有研究表明,调整行距配置可改善植株空间分布,缓和群体压力,使个体协调发展,窄行距种植条件下不仅使花后干物质积累量增加,还增加单位面积成穗数,达到高产效果[9-10]。Chen等[11]研究发现行距从30cm降至7.5cm分蘖数逐渐增加,超窄行距显著高于宽行距,在此基础上继续缩小行距至匀播状态,对群体内单株性状变异程度影响的研究未见报道。本试验以此为切入点,选用两种不同穗型冬小麦品种为材料,确定同一密度,在常规播种模式基础上,通过不断缩小行距、扩大株距的方式,研究株、行距配置对冬小麦干物质积累、转运、产量及其构成因素的影响,以期为南疆冬小麦提供适宜的株、行距配置模式及不同栽培模式下的品种选育提供参考。
1 材料与方法
1.1 试验地概况
试验于2019-2020年在新疆阿拉尔市塔里木大学农学试验站(40°32′20″N,81°17′57″E)进行,试验区位于塔里木盆地北缘,≥10 ℃年有效积温为4 113 ℃,年均气温10.8 ℃,年平均降水量50 mm左右,海拔1 015 m,无霜期220 d,属暖温带极端大陆性干旱荒漠气候区。试验地前茬作物为大豆,土壤质地为壤土,土壤有机质含量为 7.81 g·kg-1、速效磷18.3 mg·kg-1、速效钾112 mg·kg-1、碱解氮33.5 mg·kg-1、pH 7.9。
1.2 试验材料
供试冬小麦为多穗型品种‘新冬22号’(A1)和大穗型品种‘新冬50号’(A2)。
1.3 试验设计
采用裂区设计,品种为主区(A):多穗型品种‘新冬22号’(A1)、大穗型品种‘新冬50号’(A2)。株、行距配置为副区(B):设行距、株距配置为15 cm×1.7 cm(B1)、12.5 cm×2 cm(B2)、10 cm×2.5 cm(B3)、7.5 cm×3.3 cm(B4)、 5 cm×5 cm(B5)5个处理,种植密度400万 株·hm-2,小区长4 m,宽1.9 m,重复3次(图1)。2019-10-03播种,2020-06-15收获。出苗后以间苗的方式确定各处理的基本苗数,后期管理措施同一般高产田。

图1 裂区试验设计Fig.1 Split-plot experiment design
1.4 测定项目与方法
1.4.1 干物质测定 于越冬前、返青期、拔节期、开花期和成熟期取样测定干物质,将各器官分开(茎、叶、鞘、穗,成熟期将穗分为籽粒和穗轴+颖壳)装入信封置于烘箱中,105 ℃下杀青0.5 h, 80 ℃下烘干至恒质量,使用天平称其干物质量。计算公式如下:
花前营养器官干物质向籽粒转运量=开花期营养器官干物质积累量-成熟期营养器官干物质积累量
花前营养器官干物质转运率=花前营养器官干物质向籽粒转运量/花前营养器官干物质积累量×100%
花前营养器官干物质转运量对籽粒产量贡献率=花前营养器官干物质向籽粒转运量/籽粒产量×100%
花后干物质生产量=籽粒产量-花前营养器官干物质向籽粒转运量
花后干物质生产对籽粒产量贡献率=花后干物质生产量/籽粒产量×100%
1.4.2 产量测定 于成熟期在各小区内选取长势均匀一致的区域(去掉边行),划定1 m2的样方,调查有效穗数,而后将所有麦穗剪下后脱粒、风干、称质量,折算成公顷产量;每样方测产籽粒中随机取4个1 000粒烘干后称质量,最终按照籽粒含水率13%折算成千粒质量;每小区选取有代表性的20穗室内考种,调查小穗数、穗粒数等。
1.4.3 数据处理与统计分析 采用Microsoft Excel 2010进行数据统计和处理,采用DPS v 7.05进行数据分析。
2 结果与分析
2.1 株、行距配置对冬小麦干物质积累的影响
冬小麦群体干物质积累量随生育期的推进而增加,越冬期至拔节期增长缓慢,拔节期至成熟期迅速增长(图2)。‘新冬22号’于越冬前以B5处理最高,达2 596.13 kg·hm-2,较B4、B3、B2、B1处理分别高16.95%、12.02%、16.95%和 24.04%;拔节期各处理间差异不显著,表现为随行距缩小干物质积累量呈上升趋势;抽穗期各处理间存在显著性差异,以B2处理最高,达 12 347.29 kg·hm-2,B5处理最低,仅为 9 853.83 kg·hm-2;抽穗期以后,B2处理干物质积累量达最大值,显著高于其他处理,且各处理间差异显著,出现这种现象的原因可能与多穗型品种随行距缩小、株距加大,分蘖数逐渐增加,B3、B4、B5处理拔节后群体过大,抽穗期有不同程度倒伏有关;开花期,B2、B1、B3处理间差异不显著,但显著高于B4、B5处理,较B1、B3、B4、B5处理分别高 2.73%、8.31%、13.75%、20.89%;成熟期B2处理干物质积累量最高,为34 875.08 kg·hm-2,与B1处理差异不显著,但显著高于B3、B4、B5处理,且B3、B4、B5处理间差异显著。说明多穗型品种在400万株·hm-2密度下行距越小越不利于群体的光合物质积累。
‘新冬50号’越冬前以B1处理最低,仅达 1 996.10 kg·hm-2,显著低于B5、B4、B3、B2处理;拔节期以B5处理最高,与B4、B3、B2处理间差异不显著,但显著高于B1处理;抽穗期各处理间差异显著,以B5处理最高,B1处理最低;开花期B5处理的干物质积累量最高,达24 061.21 kg·hm-2,其余处理依次为B4、B3、B2、B1;成熟期各处理间差异显著,均以B5处理最高,B1处理最低。单从干物质积累情况分析,密植条件下,缩小行距更有利于大穗型品种的干物质积累。
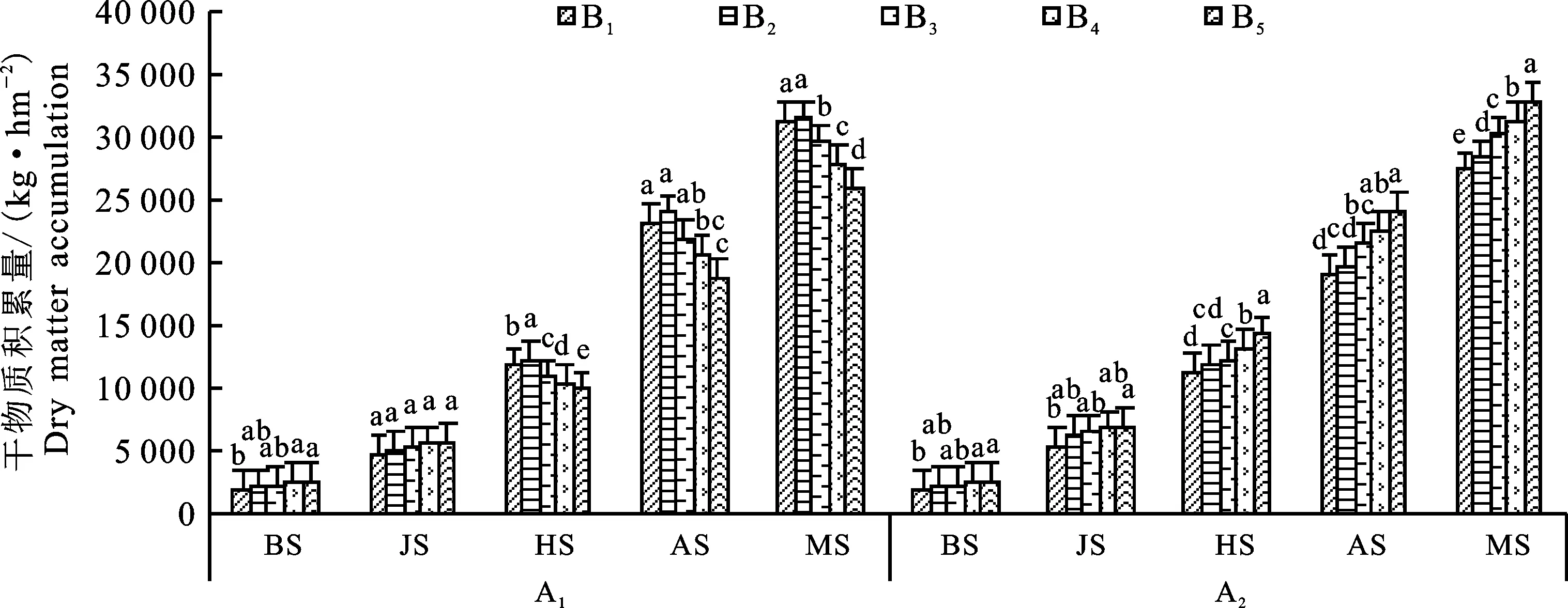
BS.越冬前;JS.拔节期;HS.抽穗期;AS.开花期;MS.成熟期。不同小写字母表示不同处理间差异显著(P<0.05)
2.2 株、行距配置对冬小麦各生育阶段干物质积累的影响
干物质是小麦光合作用产物的最终体现形式,全生育期干物质积累量因品种特性、株、行距配置等因素存在差异(表1)。

表1 不同处理冬小麦各生育阶段干物质积累量Table 1 Dry matter accumulation at different development stage of winter wheat
越冬前,‘新冬22号’干物质积累量占全生育时期8.02%左右,‘新冬50号’占全生育时期 7.93%左右,越冬前干物质积累是小麦安全越冬的基础。两穗型品种均以B5处理最高,分别为 2 596.13 kg·hm-2和2 684.13 kg·hm-2,B1处理最低,为1 972.1 kg·hm-2和19 996.1 kg·hm-2,B4处理略高于B3处理,B1均显著低于其他处理,且‘新冬50号’干物质积累量高于‘新冬22号’。说明在越冬前,行距的缩小对不同处理下两穗型品种的干物质积累影响较小,但仍表现为大穗型品种的干物质积累量多于多穗型 品种。
越冬-拔节阶段,‘新冬22号’干物质积累量占全生育时期10.48%左右,‘新冬50号’占全生育时期13.57%左右。‘新冬22号’以B5处理最高,B1处理最低,且各处理间无显著差异,干物质积累量占总干物质量的比例随行距缩小不断增加;‘新冬50号’以B5处理最高,达4 388.22 kg·hm-2,与其他4个处理差异不显著,但干物质积累量占总干物质量的比例随行距缩小呈先增后降的趋势。
拔节-抽穗阶段,干物质积累速度加快,随行距缩小,‘新冬22号’呈先增加后降低的趋势。其中以B2处理的积累量占全生育时期比例最大,达23.09%,与B1处理差异不显著,但显著高于的B3、B4、B5处理,这可能与多穗型品种B3、B4、B5处理拔节后群体过大,抽穗期有不同程度倒伏有关;‘新冬50号’随行距缩小不同处理的干物质积累量及其占总干物质量的比例均表现为先减小后增加,以B5处理最高,且各处理间差异不显著。
开花-成熟阶段,干物质积累量对最终产量形成起决定性作用。‘新冬22号’干物质积累量占全生育时期62.57%左右,该阶段B2处理干物质积累量最高,其次为B1处理,B3处理略高于B2处理,B5处理最低;‘新冬50号’干物质积累量占全生育时期积累量的57.98%左右,B5、B4、B3处理间差异不显著,但显著高于B2、B1处理,且干物质积累量占总干物质量的比例低于‘新冬22号’。
多穗型品种表现为随行距缩小,群体过大,个体间的竞争增加,抽穗期发生不同程度倒伏,不利于群体后期干物质的生产;大穗型品种随行距缩小干物质积累量呈增加趋势。
2.3 株、行距配置对冬小麦花前干物质转运和花后干物质生产量的影响
不同株、行距配置处理对不同穗型品种冬小麦的花前贮藏干物质转运、花后干物质生产量、转运及对籽粒产量的贡献率均有显著影响(表2)。
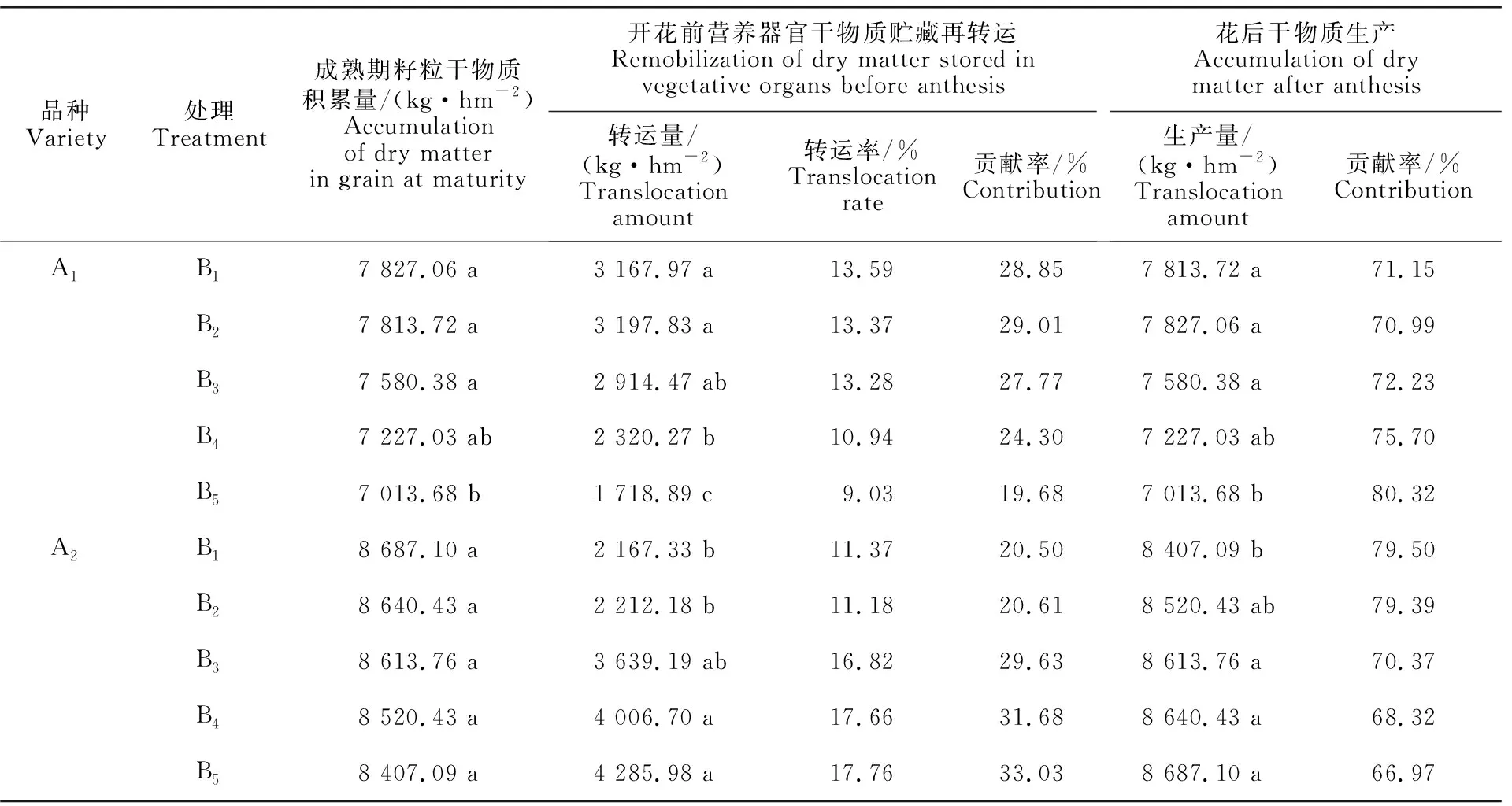
表2 不同处理下冬小麦营养器官的花前贮藏干物质转运量与花后干物质生产量Table 2 Storage dry matter transportation before flowering and photochemical accumulation after flowering in winter wheat under different treatments
随行距缩小,‘新冬22号’花前干物质转运量、花后干物质生产量均呈先增加后降低的趋势,且在B2处理达到最大,开花前B2处理为 3 197.83 kg·hm-2,与B1、B3处理差异不显著,但显著高于B4、B5处理;花后干物质生产量B2处理较B1、B3、B4、B5分别增加0.93%、 8.86%、27.44%、46.25%。‘新冬50号’品种花前干物质转运量、花后干物质生产量随行距缩小不断升高,B5处理的花前干物质转运量最大,与B4、B3、B2、B1分别对应增加6.52%、15.09%、48.39%、 49.43%,与B4、B3处理差异不显著,但显著高于B2、B1处理,花后干物质的生产量B1处理最低,仅为8 407.09 kg·hm-2,显著低于其他处理。表明适当缩小行距能显著提高冬小麦花前、花后的干物质转运。
‘新冬22号’花前干物质对籽粒贡献率为19.68%~29.01%,呈B2>B1>B3>B4>B5,干物质转运率随行距缩小逐渐降低,花后干物质的贡献率为70.9%~80.35%,呈B5>B4>B3>B1>B2;‘新冬50号’花前干物质转运率均为B5>B4>B3>B1>B2,对籽粒贡献率为20.50%~ 33.03%,花后干物质对籽粒贡献率为66.97%~ 79.50%,呈B5>B4>B3>B1>B2。说明花前干物质转运量、花后干物质生产量均会对籽粒产量产生影响,但花后干物质生产量的影响更大。
2.4 株、行距配置对冬小麦产量及其构成因素的影响
株、行距配置、品种对冬小麦产量及产量构成因素影响显著(表3)。‘新冬22号’单位面积收获穗数远高于‘新冬50号’,但小穗数、穗粒数远低于‘新冬50号’。
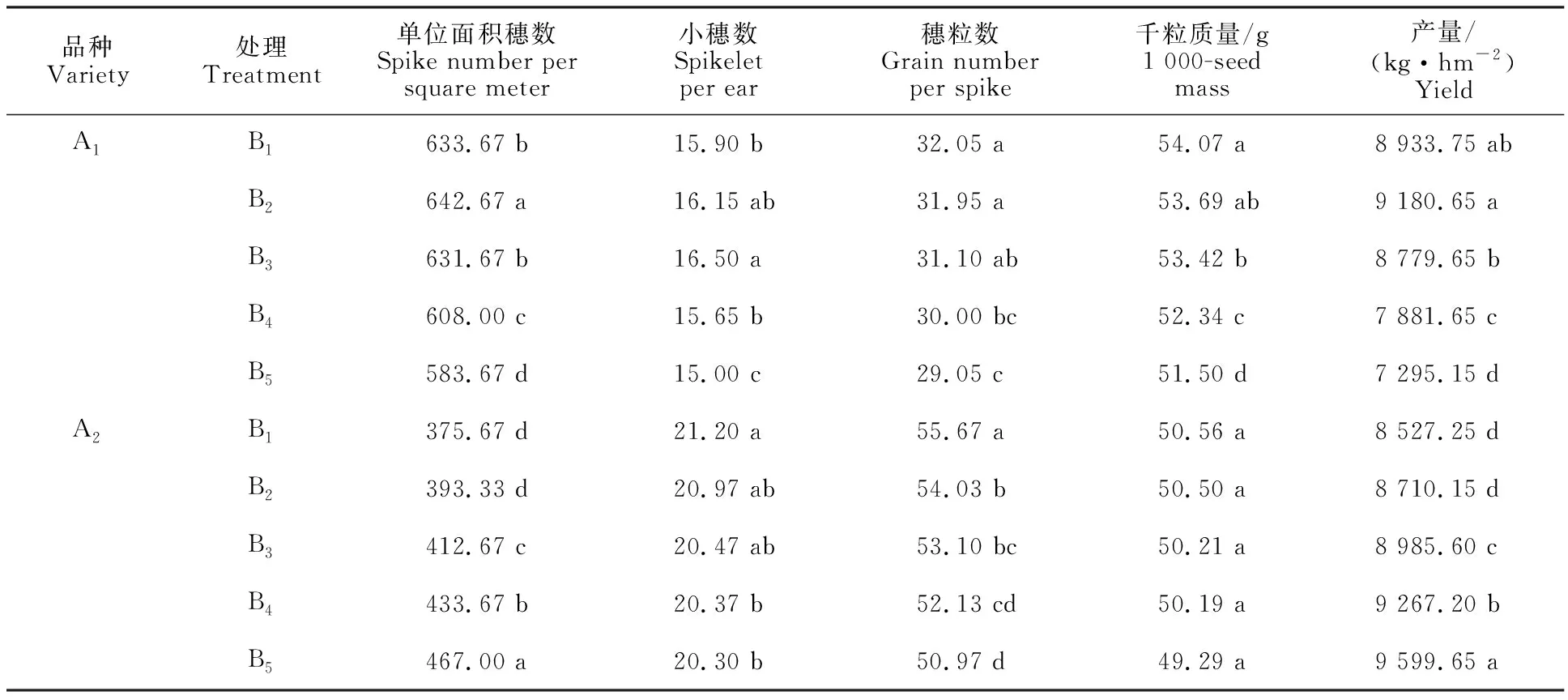
表3 不同处理冬小麦产量及其构成因素Table 3 Grain yield and yield components of winter wheat under different treatments
多穗型品种‘新冬22号’产量以B2处理最高,其次为B1处理,显著高于其他3个处理,B5处理均显著低于其他处理;单位面积收获穗数随行距减小呈先增加后减小的趋势,B2处理显著高于其他处理,B5处理显著低于其他处理,B1处理略高于B3、B4处理,与产量呈正相关;小穗数随行距减小呈先增加后减小的趋势,B3处理显著高于其他处理,B5处理最低;穗粒数、千粒质量均随行距减小而降低,B5处理显著低于其他处理,造成此现象的原因可能是B3、B4、B5处理拔节后群体过大,抽穗期有不同程度倒伏有关。
大穗型品种‘新冬50号’产量结果为B5处理显著高于其他处理,比B4、B3、B2、B1处理分别高3.58%、6.83%、10.21%、12.58%,且B1处理显著低于其他处理;单位面积收获穗数随行距缩小逐渐增加,B5处理显著高于其他处理;小穗数各处理间差异明显,随行距减小逐渐降低,B1处理最高,分别比B2、B3、B4、B5高1.10%、 3.57%、4.07%、4.43%;穗粒数、千粒质量均呈随行距缩小而降低,穗粒数处理间差异明显,千粒质量差异未达显著水平。说明缩小行距对冬小麦的增产效应主要因为单位面积穗数提高。
3 讨 论
3.1 株、行距配置对小麦干物质积累、转运的 影响
干物质是小麦光合作用产物的最终体现形式,它的积累、转运与产量的形成关系密切[12]。有研究指出,干物质积累会因品种特性、栽培措施等环境因素的不同而存在差异。抽穗期前的干物质主要用于营养器官和穗器官的构建,抽穗期后主要为了充实籽粒。大穗型品种在同一密度下以低行距效果最佳,表现出缩小行距抑制无效分蘖的作用[13],同时薛盈文等[14]也发现,在同一播种量下,小行距较大行距而言,株距拉大,植株个体间矛盾减小,导致干物质积累均随生育进程的推进呈增加趋势。邵敏敏等[15]通过设置不同行距和种植苗带对小麦群体大小进行调节,发现窄行距配置小苗带有利于小麦的干物质积累和提高花前营养器官对籽粒的贡献率。匀播种植条件下使小麦边行优势转化为单株优势,增加其分蘖数,促进干物质向籽粒产量分配,达到增产效果[16]。屈会娟等[17]研究发现开花前营养器官的贮藏干物质在开花后向籽粒的转运量对成熟期籽粒干物质积累量的贡献率较小,仅为21.00%~27.00%,花后干物质生产对籽粒的贡献率在64.00%以上。本研究结果表明,在相同密度下,小麦的干物质积累量随行距缩小逐渐增加。多穗型品种‘新冬22号’花前干物质对籽粒的贡献率为19.68%~ 29.01%,花后干物质的贡献率为70.90%~ 80.35%,大穗型品种‘新冬50号’花前干物质对籽粒的贡献率为 20.50%~33.03%,花后干物质对籽粒的贡献率为 66.97%~79.50%,可见冬小麦籽粒产量的形成同样以花后同化物转运为主,与前人研究结论一致。适当缩小行距能显著提高冬小麦的花前干物质转运和花后干物质积累,有利于提高小麦产量。
3.2 株、行距配置对小麦产量及其构成因素的 影响
小麦产量由单位面积穗数、穗粒数以及千粒质量共同决定,三因素之间既相互联系又相互制约,很难达到同步增长,往往存在负相关关系[18],冯伟等[19]研究发现密度、株、行距配置协调有利于调整个体空间分布,是解决超高产和倒伏问题的主要技术措施。在高密度条件下,缩小行距可提高分蘖成穗率,优化冠层结构,从而增加有效分蘖数,但穗粒数、千粒质量差异不显著。有研究发现,在行距缩小至10 cm时能通过增加有效分蘖而获得高产,继续缩小至7.5 cm超窄行距下仍然对产量起促进作用[20-21],而增大行距有利于千粒质量的增加;行距缩小,一定程度上抑制无效分蘖的形成,其成穗数不仅取决于分蘖数,还与群体质量有关,不同穗型品种,可结合当地生态条件,因地制宜选择合适的株、行距配置来达到高产的效果[14]。本研究认为改变株、行距距配置可以有效调节群体结构。大穗型品种‘新冬50号’株、行距距相等的B5处理产量显著高于其他处理,有效穗数与产量呈显著正相关,有效小穗数对产量有一定的影响,穗粒数、千粒质量与产量呈负相关。随着行距减小至匀播状态(株、行距相等),种子分布更加均匀,大群体单株竞争激烈,穗粒数、千粒质量有所降低。多穗型品种‘新冬22号’B3、B4、B5处理在抽穗期发生不同程度倒伏,使产量大幅度下降。其原因可能是‘新冬22号’分蘖能力较强,随着行距减小,分蘖增加,群体过大容易发生倒伏,中、低密度条件下株、行距配置对其干物质积累、运转及产量的影响如何尚待进一步研究。
4 结 论
随着行距缩小、株距扩大,植株在田间分布趋于均匀,小麦单株优势明显,群体茎蘖增加,虽然受穗粒数减少、千粒质量降低的影响,但在未发生倒伏的情况下群体收获穗数增加,最终产量呈增加趋势。
南疆400万株·hm-2种植密度条件下,冬小麦株距行距相等的均匀种植模式为最佳株、行距配置模式,但多穗型品种群体过大,存在倒伏的风险。
