铁线莲属21个类群的染色体核型分析
2022-03-17李明阳刘彦泽王鑫刘冬云
李明阳 刘彦泽 王鑫 刘冬云



摘 要: 铁线莲属(Clematis L.)为毛茛科大属之一,具有重要的园艺与药用价值。为探讨铁线莲属植物染色体组演化规律,揭示属下组间及种间的亲缘关系,该研究采用常规压片法对铁线莲属21个类群的根尖进行处理、压片,对染色体的形态特征进行观察及核型分析,同时利用Ward联接法进行聚类分析研究。狭裂太行铁线莲、毛果扬子铁线莲、卷萼铁线莲、中印铁线莲、钝萼铁线莲的染色体形态为首次报道。结果表明:铁线莲属21个类群均为二倍体,染色体数均为16(2n=2x=16),除中印铁线莲外其他类群均具随体;长瓣铁线莲、钝萼铁线莲、芹叶铁线莲、褐毛铁线莲、C. flammula、毛果扬子铁线莲的染色体为“2B”型,其他类群染色体为 “2A”型;铁线莲属核型不对称系数在60.29%~63.79%之间;铁线莲属植物染色体组表现的较为原始,种间核型存在广泛变异。综上结果表明,铁线莲属植物染色体数目应由二倍体向多倍体演化后通过多倍体的二倍化过程产生非整倍体方向演化。铁线莲属染色体的演化主要在二倍体水平上进行,通过产生染色体结构变异的方式实现,通过产生杂合染色体、加强核型不对称性、染色体类型改变以及随体染色体的变化四种途径进化。同时,核型特征在分组水平与物种水平上的划分与传统分类基本一致,说明核型分析可为铁线莲属下组一级分类提供一定的线索。该研究结果为铁线莲属植物系统分类、遗传演化与资源利用等研究领域提供了新的参考资料。
关键词: 铁线莲属, 染色体, 聚类分析, 核型分析, 细胞学
中图分类号: Q949.746.5
文献标识码: A
文章编号: 1000-3142(2022)01-0078-12
收稿日期: 2021-08-28
基金项目: 河北省铁线莲种质资源收集与新种质创新项目(1220256);现代林业学科群项目(XK1008601519) [Supported by Collection and Innovation of Clematis Germplasm Resources Project in Hebei Province(1220256); Modern Forestry Subject Group Project(XK1008601519)]。
第一作者: 李明陽(1996-),硕士研究生,主要从事园林植物遗传育种研究, (E-mail)1054468775@qq.com。
*通信作者: 刘冬云,博士,副教授,硕士生导师,主要从事地被植物资源收集及育种研究, (E-mail)dongyunliu@hebau. edu.cn。
Chromosome karyotype analysis of 21 Clematis taxa
LI Mingyang1, LIU Yanze2, WANG Xin1, LIU Dongyun1*
( 1. College of Landscape and Travel, Hebei Agricultural University, Baoding 071000, Hebei, China; 2. Management
Center of Wuling Mountain National Nature Reserve, Chengde 067300, Hebei, China )
Abstract: Clematis L. is one of the major genera of Ranunculaceae, and has important horticultural and medicinal values. In order to discuss the chromosome evolution law of Clematis plants, reveal the genetic relationship among sections and species, karyotype analysis in 21 Clematis taxa were observed. In this study, the root tips of 21 groups of Clematis were treated and pressed by conventional pressing method, the morphological characteristics of chromosomes were observed, karyotype was analyzed, and cluster analysis was carried out by Ward connection method. C. kirilowii var. chanetii, C. puberula var. tenuisepala, C. tubulosa, C. tibetana and C. peterae were reported for the first time. The results were as follows: All the 21 taxa of Clematis were diploid with x = 8 (2n = 2x = 16), the genome of each taxon had at least one pair of satellites except of C. tibetana. The chromosome of C. fusca, C. peterae, C. macropetala, C. aethusifolia, C. puberula var. tenuisepala, C. flammula were ‘2B’ type, C. acerifolia, C. fruticose, C. heracleifolia, C. intricate, C. grandidentata, C. brevicaudata, C. puberula var. tenuisepala, C. glauca, C. hexapetala, Clematis kirilowii, C. kirilowii var. chanetii, C. alpina var. ochotensis, C. tangutica, C. tibetana, C. tubulosa, C. vitalba were ‘2A’ type. The karyotype asymmetry coefficient ranged from 60.29% to 63.79%. The chromosomes of Clematis were primitive, and the karyotypes varied widely among species. Through the above research, we can draw the following conclusions: The chromosome number of Clematis should evolve from diploid to polyploid, and then aneuploidy be produced through the polyploid diploidization process. The chromosome evolution of Clematis is mainly carried out at the diploid level, which is realized by generating chromosome structural variation, and evolve through four ways: generating heterozygous chromosomes, strengthening karyotype asymmetry, chromosome type change and satellite chromosome change. At the same time, the karyotype characteristics of Clematis are basically consistent with the traditional classification at the section level and species level, which indicates that karyotype analysis can provide new reference basis for the classification of Clematis. The results of the present study have significant scientific values in the further studies on the taxonomy, phylogenetics and resources utilization in genus Clematis.
Key words: Clematis, chromosome, cluster analysis, karyotype analysis, cytology
铁线莲属植物具有重要的经济价值,一直以来都是我国常用的药用植物,其属下多种植物可起到镇痛、抗炎、抗肿瘤等作用(马迪,2010)。近年来,对其化合物方面的研究表明其含有三萜皂苷、黄酮、木脂素等成分(张亚梅等,2018)。铁线莲属植物花果形态优美,适应性强,是优良的垂直绿化植物,为国内外常用的景观花卉之一,具有极高的园艺观赏价值(高露璐等,2017)。铁线莲属(Clematis L.)分类学隶属于毛茛科(Ranunculaceae),为毛茛科中的世界性大属,有超过300个野生种类,广泛分布于全球各地,其中我国有150余种(Wang & Bartholomew, 2001; 王文采和李良千,2005)。该属种类繁多,在分类上一直是较难处理的一个类群。各国学者开展了大量的工作进行铁线莲属的系统学研究,但在不同分类系统中对其属下分类阶元的划分有较大区别(Tamura, 1995; Johnson, 1997; Grey-Wilson, 2000; 王文采和李良千,2005)。王文采和李良千(2005)基于形态特征将该属划分为四个亚属,四亚属下又分为15组。该研究为目前铁线莲属最为全面的分类系统,且属下分类群间的关系阐述清晰完整。然而,基于形态建立的分类系统对铁线莲属植物的起源演化的解释仍存在一定的问题,如属下组间亲缘关系仍不明确,而且分子系统学研究与形态分类存在巨大差别(Miikeda et al., 2006; Xie et al., 2011; Lehtonon et al., 2016)。因此,澄清铁线莲属的系统演化与属下类群关系问题仍需收集更多性状加以综合判断。
植物的核型反应了染色体演化的整体特征,研究物种的染色体及核型特征有助于判断物种间的亲缘关系和揭示其遗传进化机制(刘华敏等, 2010)。许多植物的核型特征在种级、属级乃至科级水平的分类中起到重要作用(王永,2007)。染色体与核型分析研究的重要性突出体现在毛茛科的分类学历史当中(Langlet, 1927; Gregory, 1941;Tamura, 1995;孔宏智等,1997;杨亲二,2000,2001a,b,2002)。二十世纪三四十年代,西方植物学家发现毛茛科中存在两种类型的染色体(Langlet, 1927; Gregory, 1941),即大型染色体(R type)与小型染色体(T type)。此后该特征成为毛茛科分亚科的重要参考性状(Tamura, 1995)。
铁线莲属隶属于银莲花族(Trib. Anemoneae),因而具有典型的大型染色体。因此,染色体及核型特征可能对铁线莲属分类与系统发育判断具有一定应用价值。但是,目前国内外针对染色体形态特征开展铁线莲属的细胞学研究并未清楚阐明其与基于形态的分类研究的关联,并有一定的局限性。同时铁线莲属在世界范围内广泛分布,前人的研究对部分类群覆盖范围不够完善,某些类群的染色体及核型特征数据不够完整。因此,本研究开展了针对21个铁线莲类群的染色体数目及核型研究工作,其中5个类群的染色体数据为首次报道。本研究观察了21个铁线莲类群的染色体,通过构建聚类树分析其亲缘关系,同时分析了已报道的铁线莲属染色体数目与倍性,拟探讨以下问题:(1)铁线莲属染色体演化规律;(2)核型特征在铁线莲属的分类学价值。
1 材料与方法
1.1 試验材料
试验材料为19种2变种铁线莲属植物,根据最新的分类系统,分属7组,包含华北地区铁线莲属植物组级的全部分类阶元。试验材料名称及来源见表1。所有凭证标本存放在河北农业大学植物标本室(HBAU)。试验材料均具活体植株,统一栽植于河北农业大学一分场试验地。
1.2 试验方法
于2020年8—9月期间,每种材料选取生长状况良好,根系发达的10株容器苗,上午8:00—9:00取贴近盆壁的侧根根尖分生区1~2 cm,取至少30条幼嫩根尖。将根尖置于冰水混合物中并在4 ℃冰箱预处理24 h;取出根尖用蒸馏水清洗 3~4 次,置于卡诺氏固定液 (无水乙醇∶冰醋酸 = 3∶1) 中固定 24 h;用蒸馏水冲洗,立即进行下一步解离或置于 70% 酒精中保存;将置于70%酒精中保存材料再次用蒸馏水清洗 3~4 次,加入1 mol·L-1的盐酸并在60 ℃恒温水浴锅中进行解离 10 min;将根尖用蒸馏水清洗干净,加入蒸馏水室温下低渗1 h,使细胞吸水胀大;取根尖0.2 mm的乳白色分生区于载玻片上,并用吸水纸在一侧吸干水分,滴改良卡宝品红染液染色10 min,盖上盖玻片,轻敲,压片,在Leica DM 4000显微镜下观察,并拍照。
1.3 数据获取与分析
每种铁线莲选出30个较好的分裂相,观察并统计染色体数目。再从中选取5个形态清晰且分散最好的分裂相,通过Photoshop 2020进行抠图配对,Image J测量臂长(徐炜桢等,2019),以这5个细胞各项数据的均值为参数值进行计算。根据Stebbins(1971)和李懋学等(1985)提出的核型分类标准,确定核型类型。Arano(1963)提出了核型不对称系数(asymmetrical karyotype coefficient, AS.K%)和Levan等(1964)提出了根据臂比确定着丝点位置和分类的方法。用上述方法对21个铁线莲类群的核型进行研究。并以核型不对称系数为纵坐标,平均臂比值为横坐标作21个铁线莲类群进化趋势图。
对21个铁线莲属植物的细胞学特征,包括核型不对称系数、最长染色体/最短染色体值、平均臂比、着丝点指数、有无随体、核型类型相关数值作为分类依据(盛璐,2011),选择Ward联接法,利用SPSS Statistics软件进行聚类分析。
2 结果与分析
2.1 铁线莲核型总体特征
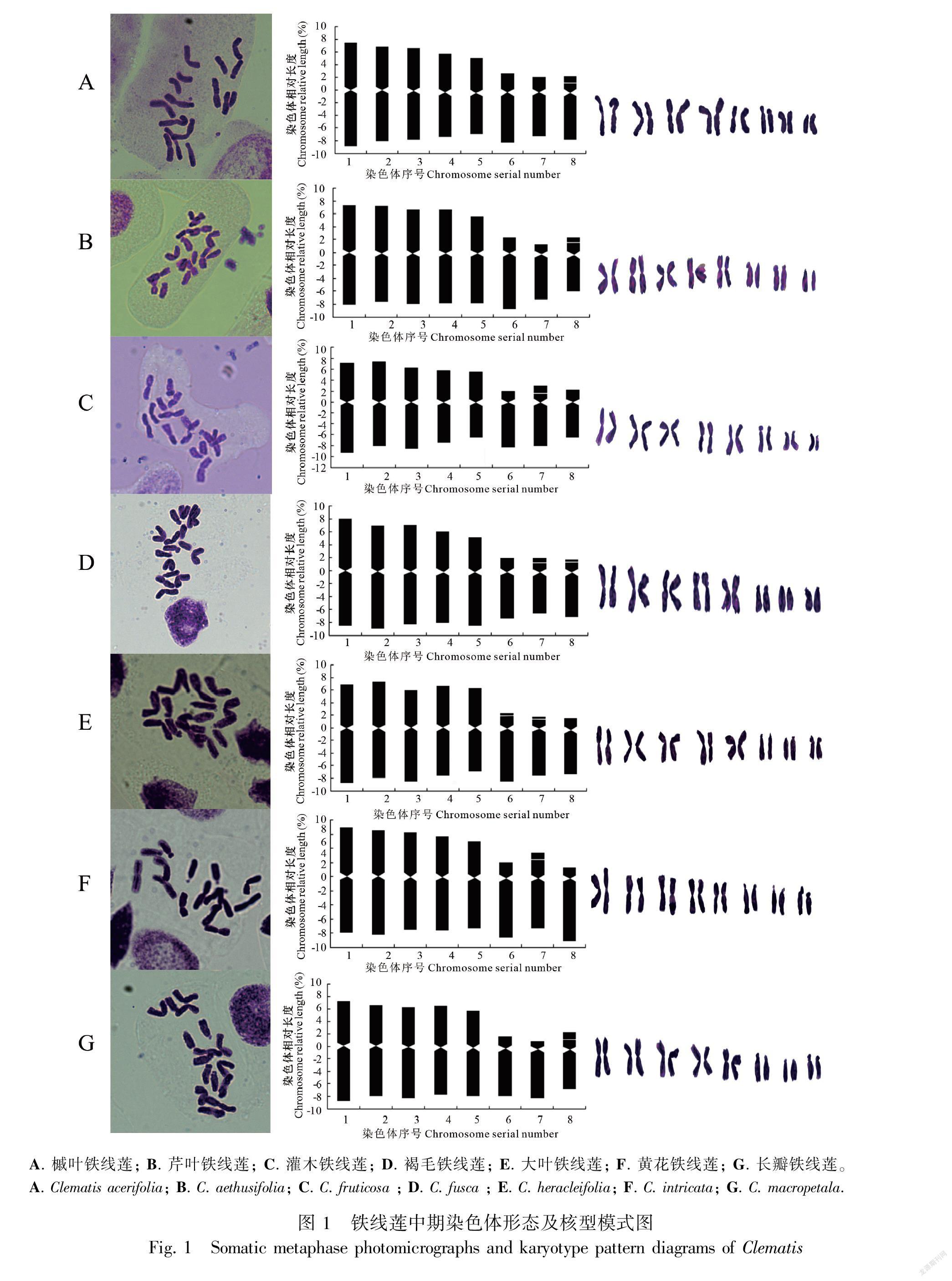
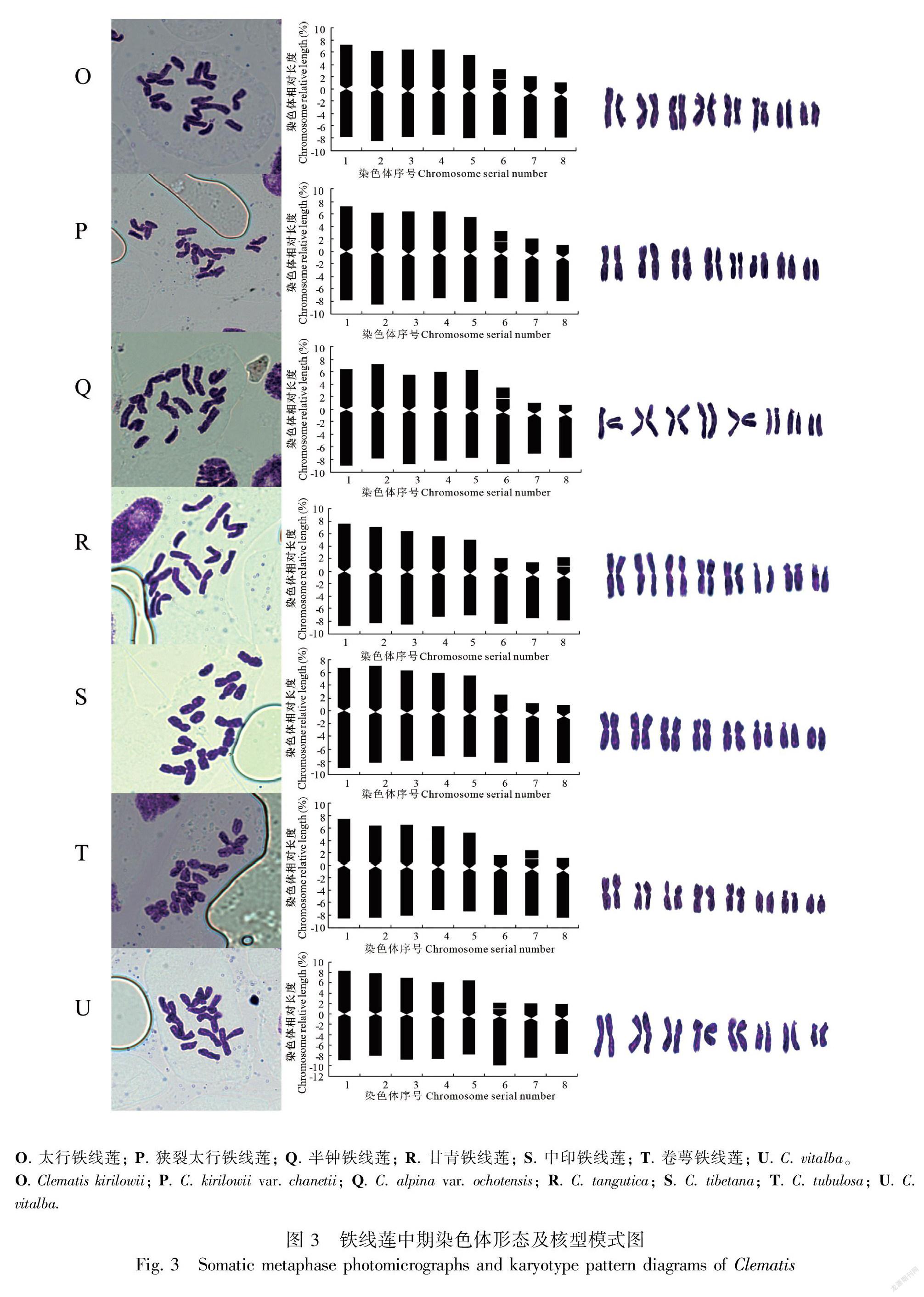
根据染色体分裂中期镜检图、染色体形态图及核型模式图(图1,图2,图3)可以看出供试的铁线莲的染色体数目稳定,观察的21个铁线莲属植物染色体数目一致,为2n=2x=16,x=8,均为二倍体植物。所有种类的染色体相对长度范围(8号染色体平均相对长度至1号染色体平均相对长度)、最长染色体与最短染色体比值、臂比值范围、核型类型、核型公式、核型不对称系数、着丝点指数、平均臂比等参数详见表2。供试植物染色体类型由m、sm、st、t组成,多数为m型。除中印铁线莲外,均具随体。供试材料染色体组中最长染色体与最短染色体比值平均值为1.89, 染色体组中臂比大于2∶1的染色体比例均为0.375, 平均臂比介于2.39~3.85之间。核型类型为“2A”和“2B”两种类型,其中,太行铁线莲、狭裂太行铁线莲、槭叶铁线莲、甘青铁线莲、黄花铁线莲、棉团铁线莲、粗齿铁线莲、灌木铁线莲、半钟铁线莲、粉绿铁线莲、大叶铁线莲、短尾铁线莲、卷萼铁线莲、中印铁线莲、Clematis vitalba的染色体为 “2A”型;长瓣铁线莲、钝萼铁线莲、芹叶铁线莲、褐毛铁线莲、C. flammula、毛果扬子铁线莲的染色体为“2B”型。核型公式可概括为2n=2x=16=10m+0~2sm+0~6st+0~4t。本试验中的21种铁线莲的核型不对称系数的范围是60.29%~63.79%。
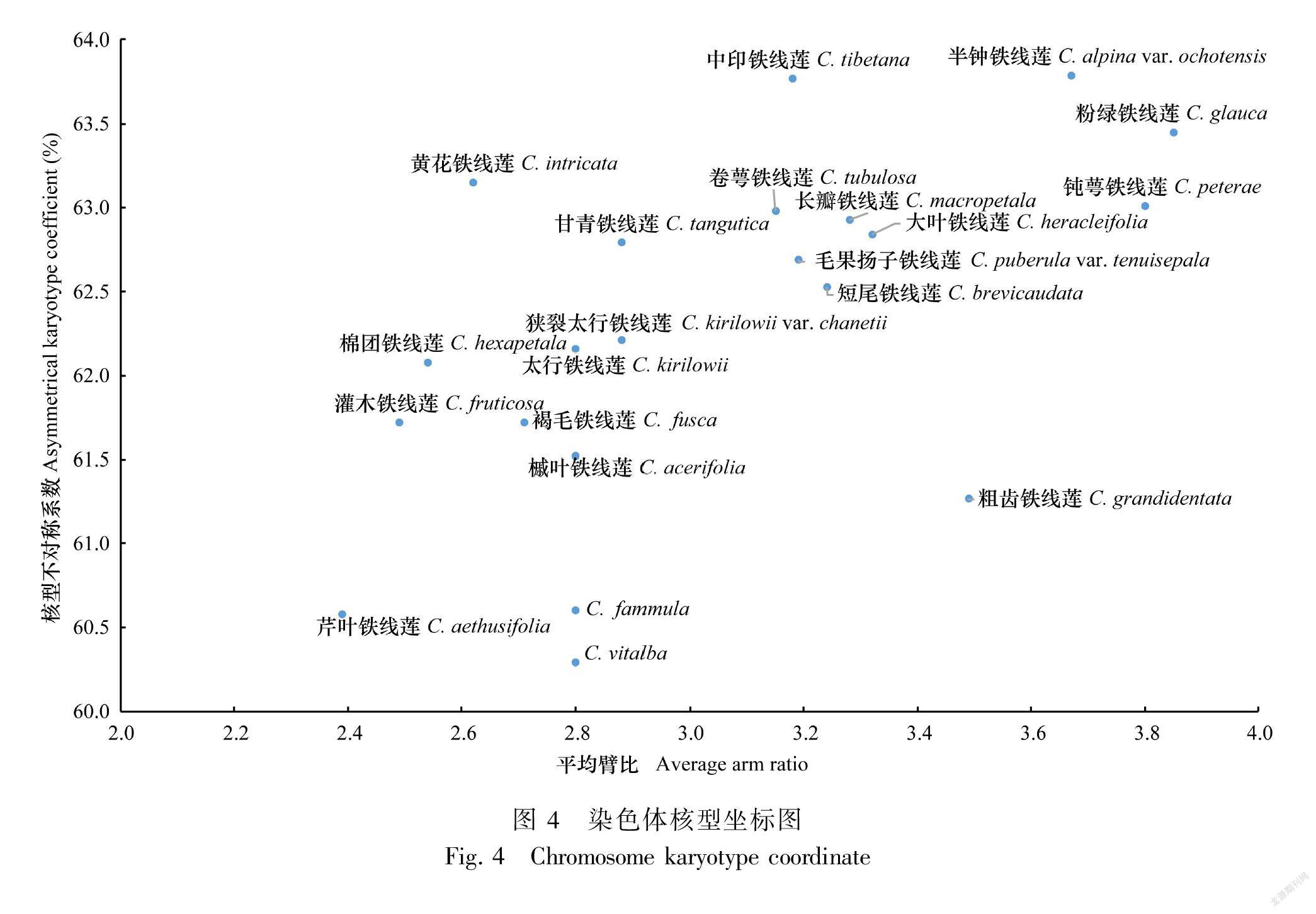
2.2 核型进化趋势分析
以平均臂比值为横坐标,核型不对称系数为纵坐标作染色体核型坐标图如图4。从中可以看出核型按双向进化趋势变化,粗齿铁线莲沿平均臂比方向进化趋势快,黄花组的中印铁线莲、黄花铁线莲、甘青铁线莲沿核型不对称性系数方向进化趋势快。同时,被子植物的染色体核型系统演化趋势是由对称向不对称发展(Stebbins,1971)。还可以看出越靠近右上方的类群,其进化程度越高。因此,芹叶铁线莲较靠近左下角,在核型上属于比较原始的类群,钝萼铁线莲、半钟铁线莲、粉绿铁线莲较靠近右上角,在核型上属于较为进化的类群。
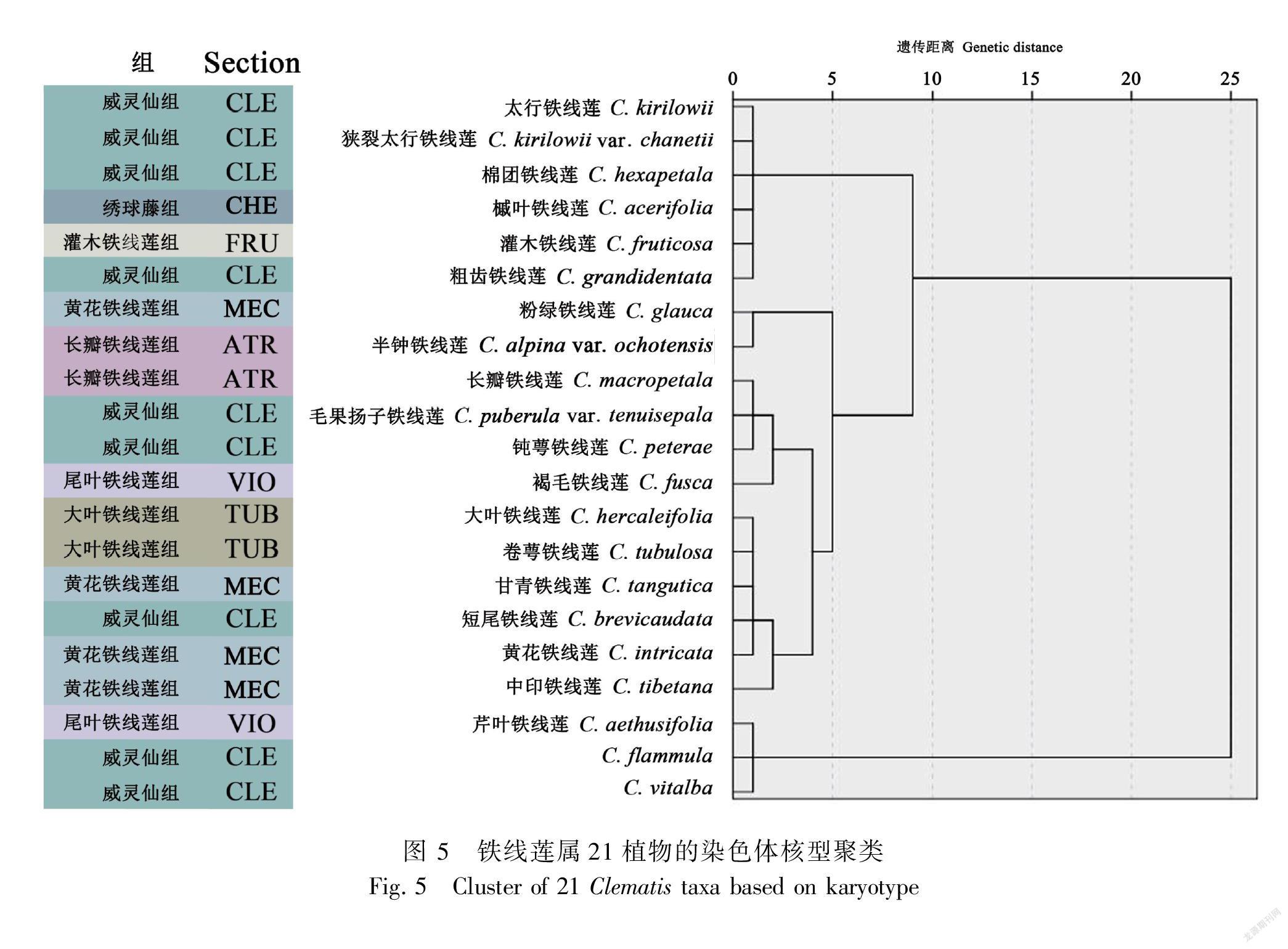
2.3 聚类分析
基于核型不对称系数、最长染色体与最短染色体比值、平均臂比、着丝点指数、有无随体、核型类型等核型参数,运用上述方法进行了聚类,聚类结果见图5。参照之前的分类处理(王文采和李良千,2005),由聚类结果可以看出,在欧氏距离10.0处,21个类群铁线莲被分为三类,第一类构成主要为威灵仙组成员,同时绣球藤组的槭叶铁线莲与灌木铁线莲组的灌木铁线莲也包含在内;第二类为黄花组、长瓣组、 大叶组以及威灵仙组的12个类群;第三类为威灵仙组的两个种以及尾叶组的一个种。灌木铁线莲组与绣球藤组各只有一种参与聚类,均与威灵仙组的四种铁线莲聚到了一起。尾叶铁线莲组的芹叶铁线莲、褐毛铁线莲在欧氏距离25.0处才聚在一起,其亲缘关系较远。此外,太行铁线莲与其变种狭裂太行铁线莲在核型相关参数相似度较高,同时在聚类结果上也支持其亲缘关系近。
3 讨论与结论
3.1 铁线莲属染色体演化规律
通过CCBD查询已报道的铁线莲属染色体数目可知,铁线莲染色体基数为8,并且存在二倍体、四倍体、六倍体、八倍体,分别对应2n=16、2n=32、2n=48、2n=64几种类型,还具2n=36、2n=42、2n=44、2n=49、2n=50等非整数倍体。在铁线莲属中, 2n=2x=16的二倍体占比最高,多倍化与非整数倍化现象并不多见,目前只在以下7个类群中有报道,如圆锥铁线莲、辣蓼铁线莲、吴兴铁线莲、拔葜叶铁线莲、C. bourdillonii、C. theobromina、C. chinensis var. fujisanensis。染色體数目演化包括多倍化和非整数倍化两种途径(王家坚等,2016),多倍体被认为比其二倍体具更好的环境适应能力(Stebbins,1971;Brochmann et al.,2004),非整倍体的形成则通常与多倍体后代形成配子过程中异常减数分裂有关(Comai,2000),因此,铁线莲属染色体数目应由二倍体向多倍体演化后通过多倍体的二倍化过程产生非整倍体方向演化。但上述七个类群均为二倍体与其他倍型的复合体,并未发现只存在多倍体或非整倍体的类群。因此,多倍体与非整数倍体在铁线莲属演化过程中作用较小。对于铁线莲属内多倍体及非整倍体的类群,其染色体数目演化与地理分布具有一定联系。在喜马拉雅地区(中国西南部、印度北部)及东亚地区,铁线莲属多倍化与非整倍体化现象频发。其中,八倍体圆锥铁线莲只在喜马拉雅地区有过报道,分布于我国其他地区的圆锥铁线莲则为四倍体。这与Ziman(1981)认为我国西南部是铁线莲属的起源与分化中心的观点相吻合。
铁线莲属染色体的演化通过产生染色体结构变异的方式主要在二倍体水平上进行。(1)产生杂合染色体,在太行铁线莲、狭裂太行铁线莲、长瓣铁线莲、灌木铁线莲、半钟铁线莲、槭叶铁线莲6个类群中均发现有一对杂合染色体,分别位于5号染色体、5号染色体、4号染色体、6号染色体、1号染色体、4号染色体。(2)核型不对称性加强,核型类型由“2A”演化为“2B”,如褐毛铁线莲、长瓣铁线莲、钝萼铁线莲、毛果扬子铁线莲。(3)染色体类型的改变,即m型染色体演化为sm型染色体,sm型染色体演化为st型染色体,st型染色体演化为t型染色体。铁线莲属染色体类型改变集中在第7和第8对染色体上,由st型向t型染色体转变。如大叶铁线莲、粗齿铁线莲、短尾铁线莲、毛果扬子铁线莲、粉绿铁线莲和卷萼铁线莲(龚维忠等,1985;张镱锂和贺士元,1990;杨亲二等,1994)。(4)随体染色体具有多态性,在数量以及类型上铁线莲属植物随体存在一定水平的变异。根据Brat (1965) 的标准,铁线莲属随体类型包含Ⅰ、Ⅱ、Ⅲ和Ⅳ型,其中Ⅲ型为主要类型。有的同时具有两种或多种演化途径。通过染色体结构演化可知,芹叶铁线莲较为原始,长瓣铁线莲、毛果扬子铁线莲、褐毛铁线莲、大叶铁线莲较为进化。这一结果得到了幼苗叶形态(Cheng et al., 2016)及系统发育(Miikeda et al., 2006; Lehtonon et al., 2016)等研究的支持。染色体演化特征对属下的系统亲缘关系研究具有重要参考价值。
3.2 铁线莲属染色体形态的分类学意义
毛茛科中铁线莲属与锡兰莲属具有密切的亲缘关系这一点早有认识(Miikeda et al., 2006; Xie et al., 2011),在毛茛科最近的分子系统研究中,也支持将锡兰莲属与互叶铁线莲属归并入铁线莲属,这恰与锡兰莲属和铁线莲属具相同的染色体数目相吻合。对银莲花族的单型属罂粟莲花属的研究显示,罂粟莲花具16条染色体,核型类型为“3A”,显示其与铁线莲属亲缘关系近,较铁线莲属“2A”或“2B”的核型类型,罂粟莲花属较为进化,获得分子系统研究(Zhang et al., 2015)的支持,结果显示罂粟莲花属与铁线莲属互为姊妹群。
21个铁线莲供试材料的聚类结果分析显示,染色体核型研究在铁线莲亚属一级未能获得分类学支持,在组一级的分类研究中,能起到一定的参考。在盛璐(2011)和彭绿春等(2012)的研究中,同为铁线莲组的湖州铁线莲、重瓣铁线莲、毛叶铁线莲、C. viticella聚为一类,同为绣球腾组的滑叶藤与金毛铁线莲聚为一类。尾叶铁线莲组的核型研究却与传统分类学(王文采和李良千,2005)产生了冲突,盛璐(2011)的聚类结果显示同为尾叶铁线莲组的宽柄铁线莲、日本铁线莲、褐毛铁线莲、C. crispa与单叶铁线莲、神农架铁线莲分别聚成两类,但尾叶组的全缘叶铁线莲与毛蕊铁线莲却并未表现出与上述类群亲缘关系较近;在彭绿春等(2012)的研究中,尾叶组的长花铁线莲与毛茛铁线莲聚为一类。本研究中同属尾叶组的芹叶铁线莲与褐毛铁线莲并未聚到一起,并显示出较远的亲缘关系,威灵仙组的10个类群也分别聚为三类。以上结果均不能很好地支持传统分类研究,却得到了幼苗叶形态研究与基于分子的系统发育研究的支持,暗示了尾叶铁线莲组与威灵仙组并非单系群。而分子生物学(穆琳和谢磊,2011; 盛璐,2015; Yan et al.,2016; Liu et al., 2018; Choi et al., 2021)的研究结果,同样支持中印铁线莲与甘青铁线莲、短尾铁线莲与大叶铁线莲与黄花铁线莲、棉团铁线莲与太行铁线莲的亲缘关系较近,说明染色体核型研究与幼苗叶形态、分子系统学研究有一定的一致性。同时,本研究与盛璐(2015)的聚类结果还显示太行铁线莲与狭裂太行铁线莲、芹叶铁线莲与宽芹叶铁线莲分别聚在一起,因此核型分析在种一级的分类上有一定的参考作用。对于不同居群的铁线莲,染色体核型分析可对其多样性提供线索(王娜等,2017)。
本研究中所采集的槭叶铁线莲核型类型为“2A”,在染色体结构上是较原始的类群,与传统分类中槭叶是较原始类群判断一致,但与北京采集的槭葉铁线莲(“2B”)在核型类型上存在差异;取自南京的大叶铁线莲比取自北京、河北的不对称性更低,在染色体结构上更为原始;山东采集的褐毛铁线莲与取自河北的核型类型也存在差异(龚维忠等,1985;王娜等,2017)。差异主要来自于不同居群进化程度不同,如环境发生改变,为在竞争中获得优势,就会比环境适宜的居群进化程度更高。本文中所采集的槭叶铁线莲居群为河北保定一新分布地,环境适宜,且在保护区中,人为干扰极少环境稳定,故其核型类型更加原始。
总之,染色体数目与核型分析在毛茛科科下的分类单元可提供一条明显的线索,在铁线莲属内显示大多数种类符合组一级的分类,但并不能够只依靠核型证据确定铁线莲在组一级的划分,在该属物种的划分上也能提供一定的参考,为物种鉴定提供依据。而在种间的核型分析中,则显示了不同居群具有较高的染色体形态变异。细胞学证据在铁线莲属系统与分类研究中可起到一定的帮助,在今后的研究中应该扩大取样并加强铁线莲属染色体方面的研究。
参考文献:
ARANO H, 1963. Cytological studies in subfamily Carduoideae(Compositae)of Japan [J]. Bot Mag Tokyo, 76(895): 32-39.
BRAT SV, 1965. Genetic systems in Allium L. chromosome variation [J]. Chromosoma ( Berl.), 16(4): 486-499.
BROCHMANN C, BRYSTING AK, ALSOS IG, et al., 2004. Polyploidy in arctic plants [J]. Biol J Linn Soc, 82(4): 521-536.
CHENG J, YAN SX, LIU HJ, et al., 2016. Reconsidering the phyllotaxy significance of seedlings in Clematis [J]. Phytotaxa, 265(2): 131-138.
CHEN YB, LI S, WU J, et al., 2021. A cluster analysis of karyotype resemblance-near coefficients in genus Epimedium L. and its evolutionary and systematic implication [J]. Guihaia, 41(1): 55-67. [陈永毕, 李双, 吴静, 等, 2021. 基于淫羊藿属(Epimedium L.)核型似近系数的聚类分析及其系统演化意义 [J]. 广西植物41(1): 55-67.]
CHOI KS, HA YH, GIL HY, et al., 2021. Two korean endemic Clematis chloroplast genomes: inversion, reposition, expansion of the inverted repeat region, phylogenetic analysis, and nucleotide substitution rates [J]. Plants, 10(2): 397-397.
COMAI L, 2000. Genetic and epigenetic interactions in allopolyploid plants [J]. Plant Mol Biol, 43(2): 387-399.
GAO LL, LI LF, MA YZ, et al., 2017. Analysis of florescence characteristics on Clematis cultivars group [J]. Acta Hortic Sin, 44(5): 921-932. [高露璐, 李林芳, 马育珠, 等. 铁线莲品种群的花期观赏性状分析 [J]. 园艺学报, 44(5): 921-932.]
GREY-WILSON C, 2000. Clematis, the Genus [M]. Oregon: Timber Press.
GONG WZ, LONG YY, LI MX, 1985. Karyotype of Clematis in Beijing [J]. J Wuhan Bot Res, 3(4): 371-379. [龚维忠, 龙雅宜, 李懋学, 1985. 北京地区铁线莲属植物的核型研究 [J]. 武汉植物学研究, 3(4): 371-379.]
GREGORY WC, 1941. Phylogenetic and cytological studies in the Ranunculaceae Juss [J]. Trans Amer Philos Soc, 31(5): 443-521.
JOHNSON M, 1997. Slktet Klematis [M]. Sdertlje: Magnus Johnson Plantskola.
KONG HZ, YANG QE, 1997. Karyomorphology and relationships of the genus Circaeaster maxim [J]. Acta Phytotax Sin, 35(6): 494-499. [孔宏智, 杨亲二, 1997. 星叶草属的核形态及其系统位置 [J]. 植物分类学报, 35(6): 494-499.]
LANGLET O, 1927. ber chromosome nverhltnisse und systematic der Ranunculaceae [J]. Svensk Bot Tidskr, 26: 381-400.
LEHTONEN S, CHRISTENHUSZ MJM, FALCK D, 2016. Sensitive phylogenetics of Clematis and its position in Ranunculaceae [J]. Bot J Linn Soc, 182: 825-867.
LEVAN A, FREDGA K, SANDBERG AA, 1964. Nomenclature for centromeric position on chromosomes [J]. Hereditas, 52(2): 201-220.
LI MX, CHEN RY, 1985. The standaraization about the karyotype analysis [J]. J Wuhan Bot Res, 3(4): 297-302. [李懋学, 陈瑞阳, 1985. 关于核型分析的标准化问题 [J]. 武汉植物学研究, 3(4): 297-302.]
LI MX, ZHANG ZP, 1996. Chromosome and research techniques of crops [M]. Beijing: China Agriculture Press: 292-297. [李懋學, 张赞平, 1996. 作物染色体及其研究技术 [M]. 北京: 中国农业出版社: 292-297.]
LIU HM, ZHI L, ZHAO LH, et al., 2010. Karyotype analysis of four wild Lilium species [J]. J Genet Genom, 11(4): 469-473. [刘华敏, 智丽, 赵丽华, 等,2010. 四种野生百合核型分析 [J]. 植物遗传资源学报, 11(4): 469-473.]
LIU QC, WANG KL, LU WP, et al., 2014. Research progress on genus Clematis in China [J]. J Genet Genom, 15(3):483-490. [刘庆超, 王奎玲, 卢婉佩, 等, 2014. 我国铁线莲属植物资源研究进展 [J]. 植物遗传资源学报, 15(3):483-490.]
LIU ZG, SHAO WL, SHEN YM, et al., 2018. Characterization of new microsatellite markers based on the transcriptome sequencing of Clematis finetiana [J]. Hereditas, 155(1) : 97-165.
MA D, 2010. Research on cold resistance of Clematis heracleifolia in Harbin [D]. Harbin: Northeast Forestry University. [ 马迪, 2010. 哈尔滨地区大叶铁线莲抗寒性的研究 [D]. 哈尔滨: 东北林业大学.]
MIIKEDA O, KITA K, HANDA T, et al., 2006. Phylogenetic relationships of Clematis (Ranunculaceae) based on chloroplast and nuclear DNA sequences [J]. Bot J Linn Soc, 152(2): 153-168.
MU L, XIE L, 2011. A preliminary study on the systematic position of Clematis acerifolia based on the analyses of ITS and plastid regions [J]. J Beijing For Univ, 33(5): 49-55. [穆琳, 谢磊, 2011. 槭叶铁线莲的系统位置初探——来自ITS和叶绿体DNA序列片段的分析 [J]. 北京林业大学学报, 33(5): 49-55.]
PENG LC, YU HJ, YU N, et al., 2012. Karyotype features of 10 species of Clematis and cluster analysis of karyotypies resemblance-near coefficient [J]. J Hunan Agric Univ (Nat Sci Ed), 38(6): 617-622. [彭绿春,于恒隽,余娜,等, 2012. 10种铁线莲的核型特征及核型似近系数聚类分析 [J]. 湖南农业大学学报(自然科学版), 38(6): 617-622.]
YAN SX, LIU HJ, LIN LL, et al., 2016. Taxonomic status of Clematis acerifolia var. elobata, based on molecular evidence [J]. Phytotaxa, 268(3): 209-219.
SHENG L, 2011. Karyotype analysis on sixteen of species genus Clematis [D]. Nanjing: Nanjing Forestry University. [盛璐, 2011. 铁钱莲属16种植物的核型分析 [D]. 南京: 南京林业大学.]
SHENG L, JI KS, YU LL, 2014. Karyotype analysis on 11 species of the genus Clematis [J]. Braz J Bot, 37(4): 601-608.
SHENG L, 2015. Karyotype analysis and molecular systematics of Clematis [D]. Nanjing: Nanjing Forestry University. [盛璐, 2015. 铁线莲属植物的核型分析及分子系统学研究 [D]. 南京: 南京林业大学.]
STEBBINS GL, 1971. Chromosomal evolution in higher plants [M]. London: Edward Arnold Press: 87-89.
TAMURA M, 1995. Ranunculaceae [N]//HIEPKO P. Die Natürlichen PflanzenFamilien. Berlin: Duncker & Humblot: 223-231.
WANG JJ, NIE ZL, MENG Y, 2016. Cytological advances on Tribe Polygonateae(Asparaganceae) [J]. Acta Bot Boreal Occident Sin, 36(4):834-845. [王家堅, 聂泽龙, 孟盈. 2016. 天门冬科黄精族细胞学研究进展 [J]. 西北植物学报, 36(4): 834-845.]
WANG N, WANG KL, LIU QH, et al., 2017. Karyotype analysis of seven wild Clematis species [J]. Acta Pratac Sin, 26(11): 123-130. [王娜, 王奎玲, 刘庆华, 等, 2017. 七种野生铁线莲属植物核型分析 [J]. 草业学报, 26(11): 123-130.]
WANG WC, LI LQ, 2005. A new system of classification of the genus Clematis (Ranunculaceae) [J]. Acta Phytotax Sin, 43(5): 431-488. [王文采, 李良千, 2005. 铁线莲属一新分类系统 [J]. 植物分类学报, 43(5): 431-488.]
WANG WC,BARTHOLOME B, 2001. Clematis [M]//Flora of China. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press, (6): 97-165.
WANG Y, 2007. Study on the cytology and flora component of Brassicaceae in Xinjiang [D]. Urumqi: Xinjiang Agricultural University. [王永, 2007. 新疆十字花科植物细胞学研究及其区系成分分析 [D]. 乌鲁木齐: 新疆农业大学.]
XIE L, WEN J, LI LQ, 2011. Phylogenetic analyses of Clematis (Ranunculaceae) based on sequences of nuclear ribosomal ITS and three plastid regions [J]. Syst Bot, 36: 907-921.
XU WZ, XU D, LIN QL, et al., 2019. Determination of rice appearance and shape parameters based on mobile phone and ImageJ software [J]. J Chin Cereal Oil Assoc, 34(10): 109-113. [徐炜桢, 许东, 林亲录,等, 2019. 基于手机拍照和ImageJ软件的大米外观形状参数的测定 [J]. 中国粮油学报, 34(10): 109-113.]
YANG QE, LUO YB, HONG DY, 1994. Karyotypes of six species of Ranunculaceae in Hunan Province [J]. Guihaia, 14(1): 27-36+99-101. [杨亲二, 罗毅波, 洪德元, 1994. 湖南6种毛莨科植物的核型研究 [J]. 广西植物, 14(1): 27-36+99-101.]
YANG QE, 2000. Ranunculus wangianus, a new species from NW Yunnan, China and its karyotype [J]. Acta Phytotaxon Sin, 38(6): 551-556. [杨亲二, 2000. 云南西北部毛茛属一新种——文采毛茛及其核型(英文) [J]. 植物分类学报, 38(6): 551-556.]
YANG QE, 2001a.Cytology of eleven species in the genus Ranunculus L. and five in its four related genera from China [J]. Acta Phytotax Sin, 39(5): 405-422. [杨亲二, 2001a. 国产毛茛属11种及其4个近缘属5种植物的细胞学研究(英文) [J]. 植物分类学报, 39(5): 405-422.]
YANG QE, 2001b. Cytology of 12 species in Aconitum L. and of 18 species in Delphinium L. of the tribe Delphineae ( Ranunculaceae) from China [J]. Acta Phytotaxon Sin,39 (6): 502-514. [杨亲二, 2001b. 国产12种乌头属和18种翠雀属植物的细胞学研究(英文) [J]. 植物分类学报, 39(6): 502-514.]
YANG QE, 2002. Cytology of the tribe Trollieae and of the tribe Cimicifugeae in the Ranunculaceae: a comparative study [J]. Acta Phytotax Sin, 40 (1): 52-65. [杨亲二, 2002. 毛茛科金莲花族和升麻族细胞学的比较研究(英文) [J]. 植物分类学报, 40 (1): 52-65.]
ZIMAN SN, 1981. Analysis of phylogenetic relations in Subtribe Clematidinae (Ranunculaceae) [J]. Ukrainskii Botanicheskii Zhurnal, 38: 4-13, 36-43.
ZHANG F, XING SY, HAN CJ, et al., 2011. Analysis on ksryotype and its evolutional trend of Ginkgo biloba var. epiphyllagerm-plasms resources [J]. Acta Hortic Sin, 38(12): 2245-2252. [张芳,邢世岩,韩晨静,等, 2011. 叶籽银杏种质资源染色体核型分析及进化趋势 [J]. 园艺学报, 38(12): 2245-2252.]
ZHANG YL, HE SY, 1990. Chromosome study of six Clematis species in China [J]. J Wuhan Bot Res, 8(2): 115-121. [张镱锂, 贺士元, 1990. 国产6种铁线莲的染色体研究 [J]. 武汉植物学研究, 8(2): 115-121.]
ZHANG YL, 1991. Studies on chromosomes of seven Clematis species [J]. J Wuhan Bot Res, 9(2): 107-111. [张镱锂, 1991. 7种铁线莲的染色体研究 [J]. 武汉植物学研究, 9(2): 107-111.]
ZHANG YM, MU ZJ, ZHANG PZ, et al., 2018. Ethnopharmacological investigation of Chinese Clematis plants [J]. Chin J Exp Trad Med Form, 24(2): 226-234. [张亚梅, 慕泽泾, 张普照, 等, 2018. 中国铁线莲属民族药用植物研究整理 [J]. 中国实验方剂学杂志, 24(2): 226-234.]
ZHANG Y, KONG HH, YANG QE, 2015. Phylogenetic relationships and taxonomic status of the monotypic Chinese genus Anemoclema(Ranunculaceae) [J]. Plant Syst Evol, 301(5):1335-1344.
(責任编辑 李 莉)
1722501186335
