模拟酸雨对耐冬山茶生理生态学特征的影响
2022-03-16王晖郭霄马翔李明燕柳健李有孙迎坤郝青李伟刘庆超刘庆华王奎玲
王晖,郭霄,马翔,李明燕,柳健,李有,孙迎坤,郝青,李伟,刘庆超,刘庆华,王奎玲
(1.青岛农业大学园林与林学院,山东青岛 266109;2.济南市林场,山东济南 250100)
酸雨是指pH小于5.6的降水[1]。近年来,由于化石燃料的燃烧,释放出大量的二氧化硫和氮氧化合物,在大气中形成酸性物质,这些酸性物质随雨雪降落到地面或植物体上,形成酸雨。目前我国已成为世界三大酸雨集中地之一。酸雨主要分布在我国东部平原、东南沿海和西南四川盆地,各地区因气候和环境存在差异,酸雨的pH和频率也有所不同[2]。对2003—2018年间全国291个城市的持续降水监测研究结果表明:全国降水酸度明显降低,其中,2003—2006 年降水酸度有所升高,而2006年之后则呈降低趋势;我国降水中的硝酸根与硫酸根离子物质的量比总体呈现升高趋势,酸雨类型正由硫酸型转变为硫酸、硝酸混合型[3]。
酸雨对植物的个体乃至整个生态系统造成不利影响[4-5]。酸雨会腐蚀植物体的外表保护层,改变细胞膜的通透性,进而侵入植物体内,破坏组织和器官,影响生理代谢功能,导致植物体发育不良[6]。主要表现为叶绿素含量降低、叶萎缩和畸形、光合作用受阻、生物量减少,甚至植物枯萎死亡[7-8]。目前,我国的酸雨范围明显扩大,资料显示我国北方地区酸雨污染形势明显加重[9],因此研究酸雨对北方树种的影响具有重要的意义。
耐冬山茶[Camelliajaponica(NaiDong)],山茶科,山茶属,常绿阔叶灌木或小乔木,是分布于胶东半岛地区的一个山茶原生种群,是世界上分布最北的山茶种群,耐低温性强,花期长,冬季开花,故当地称之为“耐冬”山茶[10]。资料显示,由于自然生境的改变、栖息地的破坏及人为干扰,大量耐冬山茶野生种群逐步减少,目前只剩500余株(丛),分别存在于长门岩岛、大管岛及崂山顶三处[11]。耐冬山茶作为北方地区原有的常绿阔叶树种,具有独特的遗传特性和重要的观赏价值,是北方地区仅有的常绿冬季开花植物,其栽培范围的扩大对于丰富北方园林冬季景观具有重要意义。
面对酸沉降日益加剧的环境,耐冬山茶幼苗对酸雨的响应机制尚不明确,且目前对植物生长影响的研究主要集中在酸雨酸度(即酸雨pH)上,关于酸雨酸度和酸沉降频率双因素对植物的生长影响研究较少。因此,本文通过温室控制试验模拟不同的酸雨酸度和酸沉降频率研究酸雨对耐冬山茶幼苗的生理生态学影响,为未来酸沉降环境下耐冬山茶资源的保护及应用提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为1年生耐冬山茶实生苗。2018年9月在青岛市植物园收集耐冬山茶种子,12月采用沙藏的方法对种子进行催芽,3月初将露白的种子种植到花盆(高15 cm、直径15 cm)中。栽培基质为草炭和园土体积比为1∶1的混合基质,6月初,筛选长势一致的耐冬山茶幼苗进行试验。
1.2 试验设计
2019年夏季在青岛农业大学温室内进行试验。试验采用全因子设计方法,设置3个模拟酸雨pH水平和3个添加频率水平。3个模拟酸雨pH分别是对照组(pH5.6)、轻度酸雨胁迫(pH4.0)和重度酸雨胁迫(pH3.0);3个添加频率: 每6 d一次为高添加频率、每12 d一次为中添加频率、每18 d一次为低添加频率,编号分别为F1、F2、F3。我国主要是以硫酸根型酸雨为主,试验选用硫酸、硝酸和氯化钙配置模拟酸雨母液,其中硫酸根离子、硝酸根离子和氯离子物质的量比为5∶1∶0.8,然后使用pHS-2C酸度计将模拟酸雨母液分别标定稀释成pH为3.0、4.0的溶液,稀释溶剂为蒸馏水,以pH5.6自来水作为对照(CK)。模拟酸雨喷洒采用小型喷雾器喷雾法,于2019年6月中旬开始喷洒,分别用pH3.0、4.0、5.6的模拟酸雨对各组植株进行胁迫处理,分别每6 d、12 d、18 d喷洒一次,每次喷洒以叶片滴液为标准,喷淋在16:00—18:00进行。本试验共9个处理组,每组7个重复,共处理63盆,试验处理时间共计108 d。整个试验过程中,采用自然光照,保证温室内通风良好,及时进行正常的养护管理工作,防止夏季经常出现的多风多雨天气的影响,定时除杂草、捉虫等。
1.3 试验指标的测定
1.3.1 生长指标的测定
在最后一次添加模拟酸雨溶液的7 d后,用精度为0.01 cm的游标卡尺测量基径,用精度为0.1 cm的直尺测量耐冬山茶幼苗的株高、冠面积。冠面积的计算采用Li等[12]的公式:
冠面积Scrown=0.25πab
其中:a为每株幼苗最大伸展处测量所得冠幅长,cm;b为在其平面垂直方向测量所得冠幅宽,cm。
摘取植株完全展开的健康成熟叶片,用电子天平称量每个叶片的鲜重(MLFW)并记录,然后用便携式叶面积仪(Yaxin-1241,北京雅欣理仪科技有限公司)测量叶片面积(SLA)并记录。最后将叶片放入烘箱中用85 ℃烘48 h使其完全脱水后称量其干重(MLDW)。根据公式计算比叶面积(SLA)和叶片含水量(LWC):
SLA=SLA/MLDM×100%
LWC= (MLFM-MLDM)/MLFM×100%
1.3.2 气体交换参数的测定
使用便携式CIRAS-3光合仪(PP SYSTEMS公司,美国)对植株从上往下的第2~3片叶进行气体交换参数的测定,有净光合速率(Pn)、蒸腾速率(E)、气孔导度(Gs)、细胞间隙二氧化碳浓度(Ci)等参数。测量时的参数设置为光强 1 000 μmol·m-2·s-1,叶室温度 27 ℃, 湿度为 60%,CO2浓度为 400 μmol·mol-1。每个处理测定7个重复。根据公式计算气孔限制值(Ls):
Ls=1-Ci/Ca,其中Ca为空气CO2浓度。
1.3.3 叶绿素含量的测定
用乙醇提取法和分光光度计法测定叶绿素95%乙醇提取液在665 nm和649 nm处的吸光度D665和D649,并根据公式计算相关的叶绿素含量:
Ca=13.95D665-6.88D649;
Cb=24.96D649-7.32D665;
叶绿体色素的含量CChl=(C×V)/Mch1;
总叶绿素含量:CT=Ca+Cb;
叶绿素a/b(Chl a/b )=Ca/Cb;
其中:C为色素浓度,mg/L;V为提取液体积,mL;Ca为叶绿素a含量,mg·g-1;Cb为叶绿素b含量,mg·g-1;Mch1为样品干重,g。
1.3.4 植株生物量指标的测定
以上参数全部测量完毕后,将试验所用的全部的63棵耐冬山茶幼苗整株挖出,切割分解为根、茎、叶三部分,分别放入烘箱中80 ℃ 烘48 h,最后使用电子天平称量根、茎、叶干重并计算总生物量、根生物量比、叶生物量比及根冠比。
总生物量MTB=MRB+MSB+MLB;
根生物量比(RMR)=MRB/MTB;
叶生物量比(LMR)=MLB/MTB;
根冠比(RS)=MRB/ (MLB+MSB)
其中:MRB为根生物量,MLB为叶生物量,MSB为茎生物量,MTB为总生物量。
1.4 数据处理
用Excel2019录入数据,釆用SPSS21对数据进行正态检验和方差齐性检验后进行单因素和双因素的ANOVA分析,比较各个参数分别在酸沉降和不同添加频率下的差异,以及两因素间是否存在交互作用。存在显著差异之后进行邓肯(Duncan)多重比较(α=0.05),并用字母进行标记。
2 结果与分析
对生长指标、叶形态特征、气体交换参数、叶绿素含量及生物量进行双因素方差分析与研究,所得结果如下:
2.1 生长指标对不同模拟酸雨的响应
模拟酸雨pH值、添加频率及二者交互作用对耐冬山茶株高和基径的影响不显著,冠面积受添加频率的影响极显著(表1),在不同pH下均在中添加频率时达到最大值。如图1所示,在pH4.0时,随着添加频率的下降,耐冬山茶幼苗的基径先减少后增大(图1B),而在pH5.6和pH3.0时,基径先增大后减少,但差异不显著。
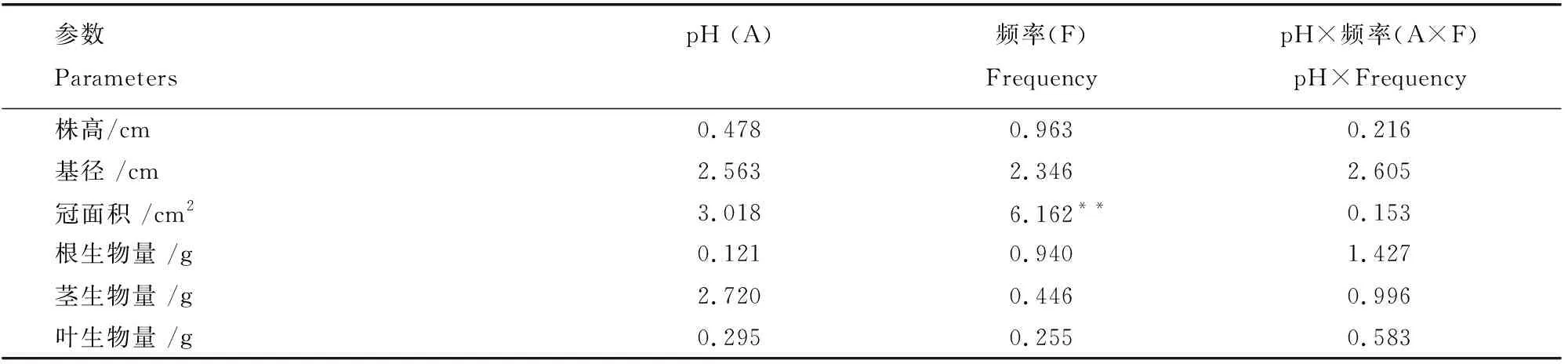
表1 耐冬山茶幼苗生理生态指标的双因素方差分析(以酸度和频率作为固定因子)Table 1 Results of two-way analysis of variance (ANOVA) for Camellia japonica (Naidong)seedlings on ecophysiological parameters, with acidity and frequency of acid deposition as fixed factors
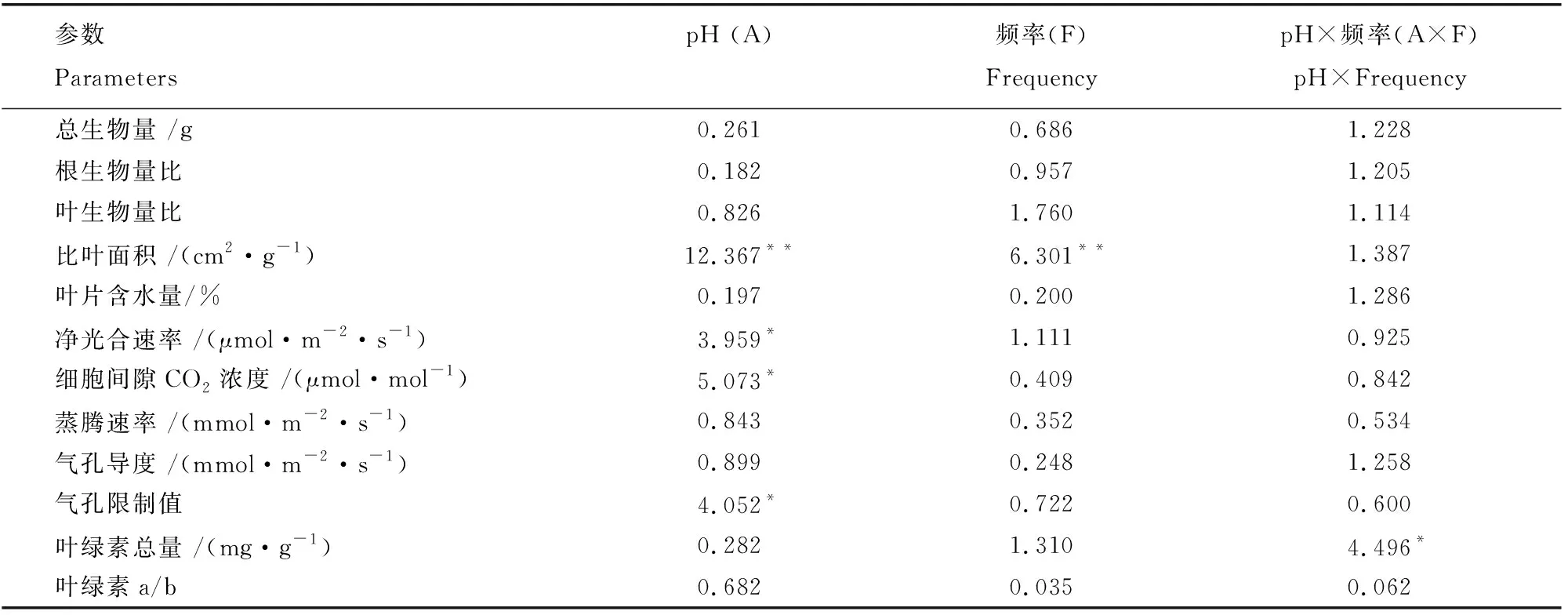
续表
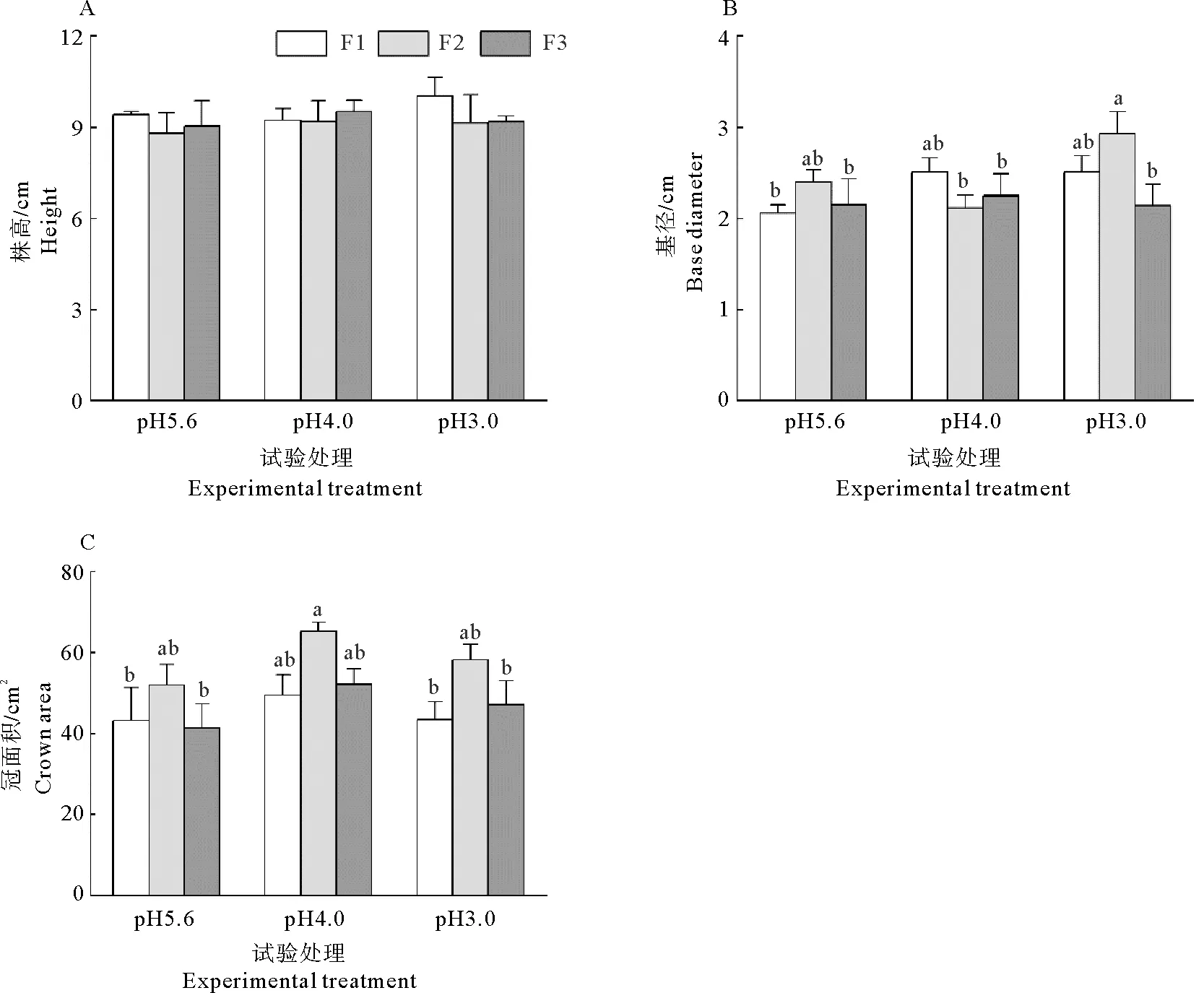
图1 不同的酸度和酸沉降添加频率对耐冬山茶幼苗生长指标的影响Fig. 1 Effects of different acidities and frequencies of acid deposition on growth parameters of C. japonica (NaiDong) seedlings注:不同小写字母表示不同处理间在P≤0.05水平差异显著,下同。
2.2 生物量对不同模拟酸雨的响应
耐冬山茶的生物量受模拟酸雨pH、添加频率及两者的交互作用的影响不显著(表1)。如图2A所示,在模拟酸雨pH3.0时,高添加频率下根生物量最大,这说明高频率的重度模拟酸雨胁迫使耐冬山茶幼苗增加了地下部分的生长。茎生物量、叶生物量和总生物量的变化不显著(图2B、2C、2D)。
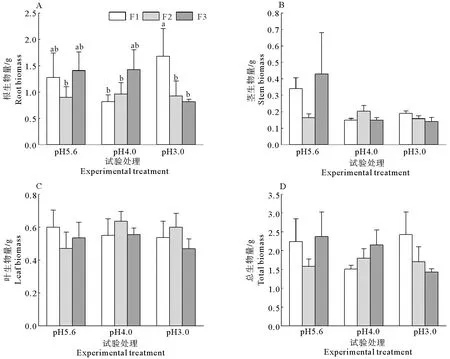
图2 不同的酸度和酸沉降添加频率对耐冬山茶幼苗生物量的影响Fig. 2 Effects of different acidities and frequencies of acid deposition on biomass parameters of C. japonica (NaiDong) seedlings
2.3 叶形态特征对不同模拟酸雨的响应
叶片含水量不受模拟酸雨pH、添加频率及两者交互作用的影响,而比叶面积则受到模拟酸雨pH和添加频率的影响,二者的交互作用对其无影响(表1,图3A、3B)。
由图3A可知,在同一pH值的模拟酸雨下,耐冬山茶比叶面积随着添加频率的降低先升高后下降,但差异并不显著。在添加频率为F2时比叶面积达到最大值,在pH5.6、pH4.0及pH3.0条件下分别为120.7、105.32、124.3 cm2·g-1。 总体看,随着酸雨pH的降低及模拟酸雨施加频率的降低,比叶面积增大。
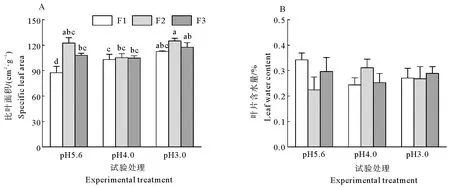
图3 不同的酸度和酸沉降添加频率对耐冬山茶幼苗叶形态特征的影响Fig. 3 Effects of acidities and frequencies of acid deposition on leaf morphological traits of C. japonica (NaiDong) seedlings
2.4 叶绿素含量对不同模拟酸雨的响应
耐冬山茶叶片的总叶绿素含量受模拟酸雨pH和添加频率二者交互作用的影响显著(表1)。由图4A可知,在高添加频率条件下, pH4.0和pH3.0处理下的总叶绿素含量与对照比分别提高了36.04%和132.92%;在中频添加处理下,总叶绿素含量随酸度增加则呈下降趋势,pH4.0和pH3.0处理下与对照比分别下降了42.96%和52.34%;低添加频率的处理下叶绿素总量不受模拟酸雨pH改变的影响,这说明,耐冬山茶幼苗对低频率的模拟酸雨有一定的抵抗作用。叶绿素a/b未受模拟酸雨pH影响(表1,图4B)。
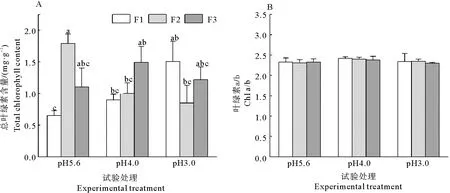
图4 不同酸度和酸沉降添加频率对耐冬山茶幼苗叶绿素含量的影响Fig. 4 Effects of acidities and frequencies of acid deposition on chlorophyll content of C. japonica (NaiDong) seedlings
2.5 气体交换参数对不同模拟酸雨的响应
耐冬山茶的气体交换参数不受添加频率和两者交互作用的影响,净光合速率(Pn)、细胞间隙CO2浓度(Ci)和气孔限制值等参数受模拟酸雨pH影响显著,蒸腾速率、气孔导度受模拟酸雨影响不显著(表1)。总体看,随着模拟酸雨pH的降低,净光合速率、气孔限制值降低,细胞间隙CO2浓度呈增加趋势(表1,图5A、5C、5D)。在同一添加频率下,随着模拟酸雨的pH值降低,耐冬山茶幼苗的净光合速率呈下降趋势,且pH3.0的处理低于其余两组,但不显著。在F1和F2添加频率下,Ci随着模拟酸雨pH的降低而增加,但增加并未达到显著水平,F2添加频率处理的叶片Ci在pH3.0模拟酸雨的胁迫下相较于pH5.6和pH4.0分别提升了177.67%和74.84%。在高、中添加频率处理下,随着模拟酸雨pH的降低,气孔限制值逐渐降低,而在低添加频率处理下,气孔限制值不受模拟酸雨pH的影响。
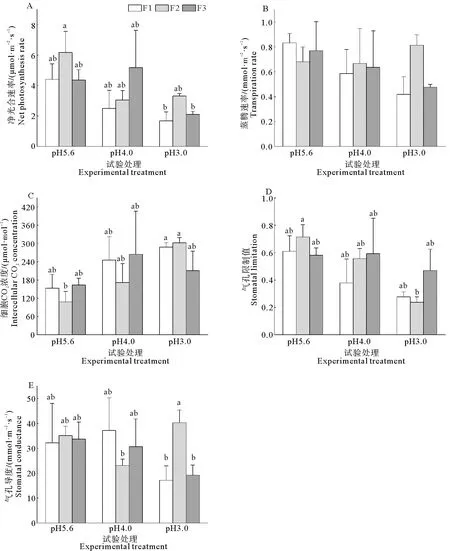
图5 不同的酸度和酸沉降添加频率对耐冬山茶幼苗气体交换参数的影响Fig. 5 Effects of acidities and frequencies of acid deposition on gas exchange parameters of C. japonica (NaiDong) seedlings
3 讨论
酸雨对植物的生长发育有着极大的影响[13],不同pH的酸雨对植物的影响是不同的,而酸沉降频率的差异会导致酸雨对植物的胁迫时间不同,也会对植物产生影响[14]。
酸雨对植物造成伤害的程度受植物种类和接触时间的影响,最重要的因素是酸雨的pH值。有研究表明,酸雨对植物的伤害主要表现在对叶片的直接伤害[15],酸雨有强烈的腐蚀性,随着酸雨pH的降低及胁迫时间延长以至超过了植物的耐受限度时,植物叶片表层结构上的蜡质和角质层将会受到破坏,保护组织的正常功能失效,进而大量的酸性物质就会通过表皮和气孔进入到植物叶片细胞内,使植物细胞中毒,植物正常的生理代谢和生长发育都会因此受到干扰[16]。本研究中,耐冬山茶总生物量不受模拟酸雨酸度、施加频率及二者交互作用的影响,说明耐冬山茶对短期模拟酸雨胁迫具有较强抗性与忍耐力。从耐冬山茶叶片的形态结构看,耐冬山茶叶片革质、表面光滑,且在外层有着较为厚实的蜡质层和角质层,酸雨无法在叶片上长时间滞留,酸雨对耐冬山茶的影响就会降低,因此模拟酸雨未对耐冬山茶总生物量造成显著影响。
双因素方差分析结果表明,添加频率显著影响了耐冬山茶的冠面积。随着添加频率的降低,耐冬山茶幼苗的冠面积先上升后下降,表明添加频率对耐冬山茶的影响存在一定的阈值。可能在较高的添加频率下,植株频繁接触酸雨,使植株的胁迫时间延长,导致植株受酸雨胁迫严重[17],进而影响了耐冬山茶正常的生长发育。前人多次研究表明,低浓度的酸雨并不会对植物的正常生长发育造成不利影响,部分植物还会因低浓度的酸雨而得到促进作用,但高浓度的酸雨则会抑制植物的生长[18-20]。本研究中,在同一模拟酸雨pH下,间隔较长时间的低添加频率条件下耐冬山茶幼苗的冠面积相比中等添加频率有所下降,原因可能是模拟酸雨中含有氮元素、硫元素等营养物质,适当频率的模拟酸雨或许会增加植株可利用的营养,进而促进植株生长。
比叶面积指的是叶的单位面积与其干物质质量的比值,是反映叶片厚度与植物生长速率的重要参数,受环境因素变化的影响。本研究中,模拟酸雨酸度增加了比叶面积,说明更高的模拟酸雨酸度,加速了对叶片表面的腐蚀作用,使叶片变薄;施加频率降低了比叶面积,其原因可能是频繁酸沉降条件下,耐冬山茶增加了向叶片细胞壁的投入,使叶片增厚以抵抗酸雨。
植物光合器官的功能与膜系统息息相关,随着酸雨pH的降低及胁迫时间延长以至超过了植物的耐受限度时,植物叶片表层结构就会遭到破坏,进而影响植物的细胞膜。酸雨破坏了植物叶片的组织形态学结构,改变了叶片生理生化特性,叶片中的酶、叶绿素等都会在酸性物质的侵蚀下失效分解,进而抑制植物的光合作用[21-23]。在本试验中,在高添加频率下,随添加模拟酸雨的pH降低,耐冬山茶叶片的总叶绿素含量相较于对照组增加,这与王玉魁等[24]对毛竹的研究结果相似,这有可能与山茶属植物自身的生活习性有关,山茶属植物喜酸,微酸的壤土和腐叶土最适合其生长,同时酸雨中含有大量的氮元素,促进了叶绿素的合成,并且轻度的酸雨对植物叶片的侵蚀作用不严重,导致了叶绿素的大量积累。
耐冬山茶的净光合速率随着模拟酸雨的pH的降低逐渐下降,其细胞间隙CO2浓度的变化趋势与净光合速率相反,按照Farquhar和Sharkey的理论[25],这也说明了耐冬山茶净光合速率的降低是非气孔限制型。
酸雨与添加频率的交互作用对耐冬山茶的影响并不显著,仅对叶绿素含量有影响。我们推测其可能有两方面的原因,一方面是耐冬山茶幼苗的生长发育缓慢,而酸雨对植物的影响是需要较长时间才能显现的,因此较短时间的酸雨处理不能对耐冬山茶幼苗的各项生理活动产生较大影响;另一方面,由于山茶属植物喜欢生长在偏微酸的环境当中,其对酸性的环境有着更高的耐受度,因此在面对酸雨胁迫时,短时间内的正常生理活性不会发生较大改变,这就使得酸雨和添加频率的交互作用对耐冬山茶植株的作用不明显。在轻度胁迫下,酸雨胁迫对叶肉细胞光合活性有一定的刺激作用,而采用高频度处理,胁迫次数增多,对叶肉细胞的伤害程度也大[26]。耐冬山茶叶片革质,表皮覆盖较厚蜡质层和角质层,对酸雨液滴的滞留时间较短,其结构形态特点对酸雨中有害离子侵入具有抵抗作用,重度酸雨胁迫下也未见明显伤斑[27]。
4 结论
模拟酸雨酸度主要影响耐冬山茶幼苗的比叶面积、净光合速率、细胞间隙CO2浓度和气孔限制值;添加频率影响了冠面积和比叶面积;模拟酸雨酸度和添加频率的交互作用仅影响总叶绿素含量。耐冬山茶对短期模拟酸雨胁迫具有较强抗性与忍耐力,其总生物量未受模拟酸雨酸度、施加频率及二者交互作用的影响。
模拟酸雨对耐冬山茶幼苗生长的影响,可为进一步开展酸沉降格局对陆地生态系统的影响提供科学数据和理论基础。
