棘头梅童鱼消化道形态与组织学结构特征
2022-03-16徐万土陶顺顺徐芳君
徐万土, 陶顺顺, 曾 肖, 徐芳君, 陈 裕
棘头梅童鱼消化道形态与组织学结构特征
徐万土, 陶顺顺, 曾 肖, 徐芳君, 陈 裕
(象山港湾水产苗种有限公司, 浙江 象山 315700)
为了解棘头梅童鱼消化道形态结构特点, 采用解剖及石蜡切片技术对其进行研究. 结果显示, 棘头梅童鱼的消化道由口咽腔(舌)、食道、胃、幽门盲囊和肠组成. 舌由基舌骨突出外覆黏膜而成, 黏膜上皮为复层扁平上皮, 内含少量味蕾及黏液细胞. 食道、胃、幽门盲囊及肠均可分为黏膜层、黏膜下层、肌层及浆膜层. 食道粗短, 黏膜层被覆复层扁平上皮, 内含大量黏液细胞. 胃呈卜型, 且具有体积较大的胃盲囊. 胃上皮为单层柱状上皮, 上皮之下的固有膜内分布有大量胃腺. 幽门盲囊位于胃与前肠连接处, 呈指状, 有12~17根, 其组织学结构与肠道相似. 肠呈“S”型盘曲, 肠指数为0.72±0.10, 黏膜层被覆单层柱状上皮, 其间分布有少量杯状细胞. 棘头梅童鱼消化道各部形态及组织学结构特征与其食性相适应.
棘头梅童鱼; 解剖学; 组织学; 食性

棘头梅童鱼()属鲈形目, 石首鱼科, 梅童鱼属(), 为我国重要的海产经济鱼类之一, 具有繁殖能力强, 生长速度快, 适盐、适温广等特点, 且在我国的沿海近岸广泛分布[8]. 棘头梅童鱼属典型的肉食性鱼类, 通常以小型虾类(糠虾、磷虾等)及桡足类为食, 并兼食鱼类及长尾类等[9]. 近年来, 棘头梅童鱼自然资源已出现严重衰退, 主要诱因有水环境污染、过度捕捞等, 因此, 其人工繁殖及增养殖开发引起重视. 目前, 有关棘头梅童鱼的生物学研究已涉及食性[9]、资源评估[10-11]、基因[12]、遗传多样性[13]等方面, 而其消化道形态及组织学结构特征尚未见报道. 因此本文采用解剖学及组织学技术对其进行研究, 旨在积累该鱼消化生理方面的基础资料, 为其人工养殖提供参考.
1 材料与方法
1.1 材料
2018年12月于浙江省宁波市慈溪附近海域捕获野生棘头梅童鱼21尾, 平均体长(11.19±0.9)cm, 体质量(19.93±4.25)g.
1.2 方法
取实验用鱼进行解剖, 观察其消化道形态后拍照, 并测量体重、体长、肠长等指标. 组织学样品主要取其舌、食道、胃、幽门盲囊、肠道等消化器官, 在波恩氏液中固定24h后, 于梯度酒精中脱水, 二甲苯透明, 石蜡包埋, 切片机(YD-202型)切片, 厚度6~7μm, 切片完成后于苏木精-伊红(HE)染液中染色, 并用中性树胶封片, 尼康显微镜(DS-Ri2型)观察并拍照. 消化道不同部位形态参数采用Photoshop软件进行测量, 黏液细胞密度以每次视野中300µm×300µm范围内的细胞个数为准(=3). 肠指数=肠长/体长.
1.3 数据处理
采用Excel对实验数据进行统计及处理, 统计数据均以平均值±标准差(Mean±SD)表示, 用SPSS 20.0软件中的单因素方差分析(One-way ANOVA)检测显著差异水平, 设=0.05.
2 结果
2.1 消化道外形特征
棘头梅童鱼的消化道由口咽腔(舌)、食道、胃、幽门盲囊及肠组成. 食道粗短; 胃卜型, 且具有体积较大的胃盲囊; 幽门盲囊呈指状, 位于胃与前肠相连处, 有12~17根(=21); 肠呈“S”型弯曲, 可以后2个转折处为界, 将其划分为前、中、后肠, 前肠肠径较中后肠大, 后肠肠长较前中肠稍长, 其末端以肛门开口于体外(图1). 测得21尾棘头梅童鱼肠指数为0.72±0.10.
2.2 消化道组织学结构
2.2.1 舌
舌由基舌骨突出外覆黏膜层而成(图2(A1)), 黏膜层可分为黏膜上皮及固有膜, 上皮为复层扁平上皮, 其中分布有黏液细胞及少量味蕾, 味蕾由固有膜隆起而成的乳头状突起所支撑, 固有膜之下为黏膜下层, 主要由疏松结缔组织构成(图2 (A2)、(A3)).
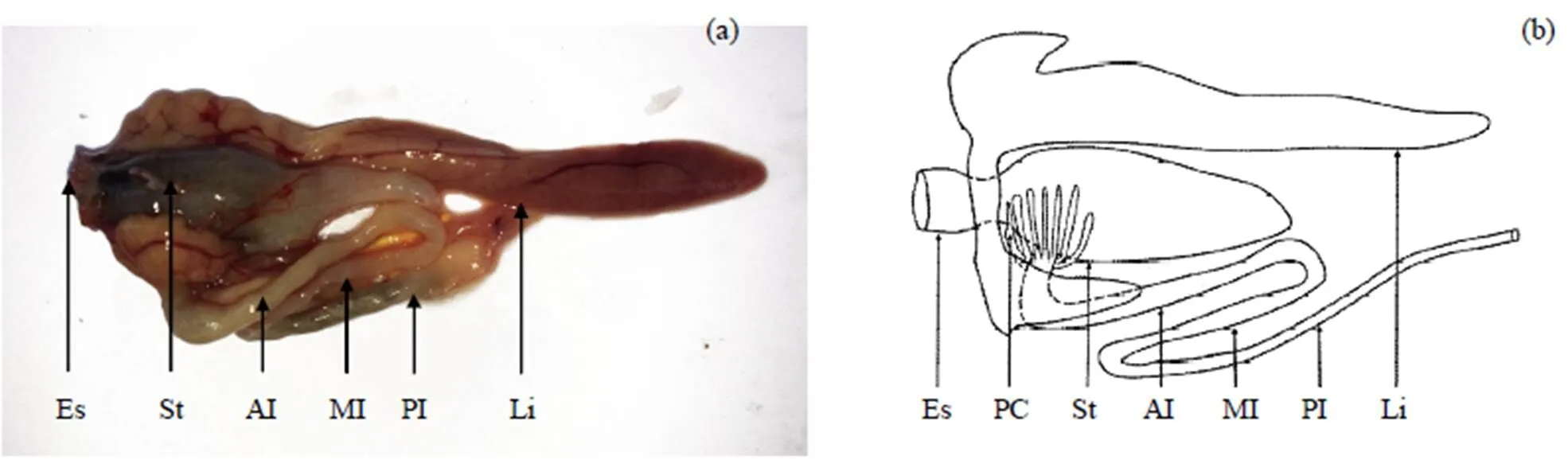
Es: 食道; St: 胃; AI: 前肠; MI: 中肠; PI: 后肠; Li: 肝脏; PC: 幽门盲囊.
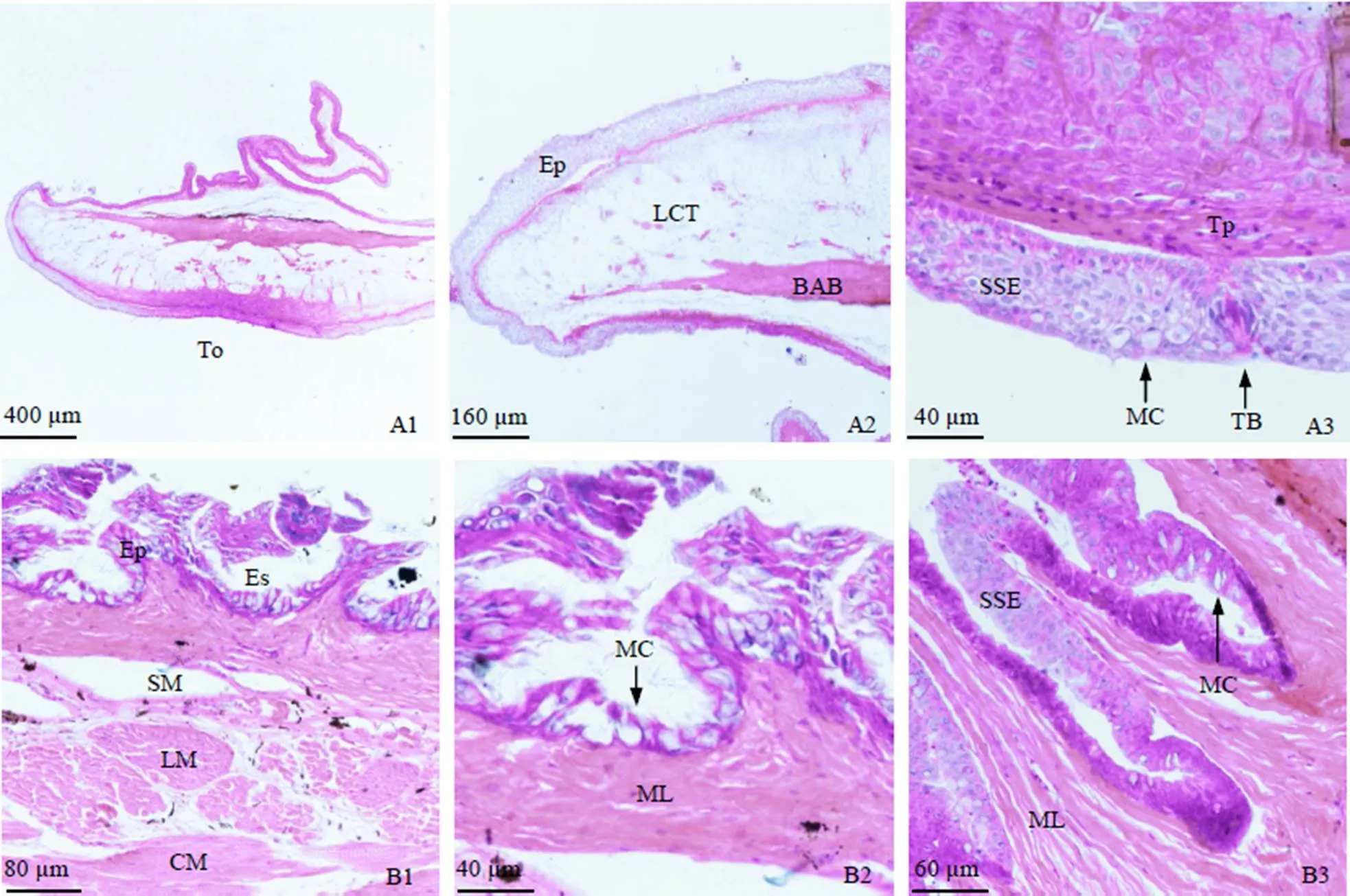
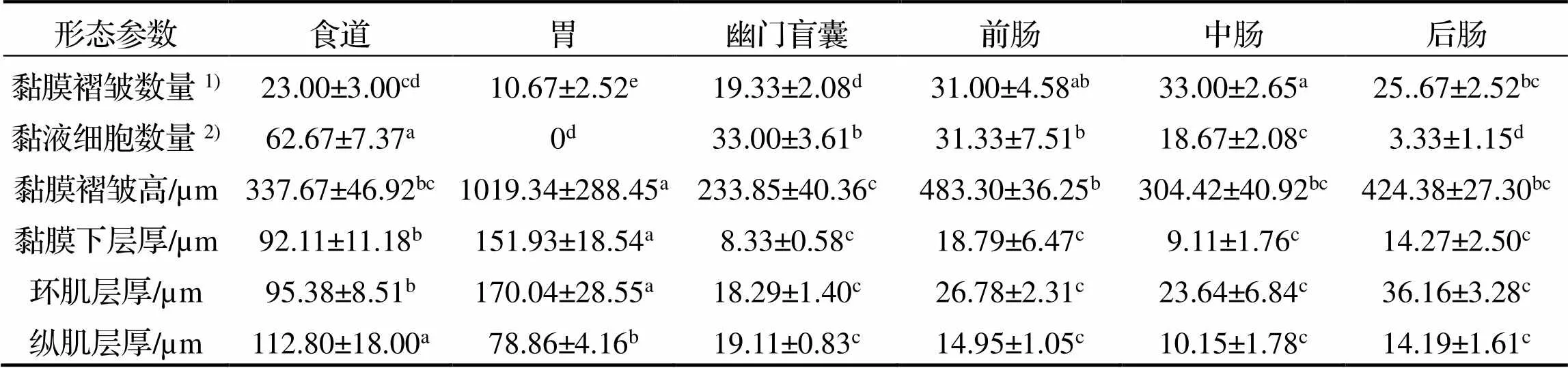
表1 棘头梅童鱼消化道组织形态测量参数
注: 1)单横切片面; 2)按0.09mm2中的个数进行计算; 同行上标字母不同表示差异显著,<0.05
2.2.2 食道
食道由内到外分别为黏膜层、黏膜下层、肌层及浆膜层(图2(B)). 黏膜层向食道腔内延伸, 形成较多树状突起, 被覆复层扁平上皮, 其间分布有大量黏液细胞(图2(B2)、(B3)). 黏膜下层主要为结缔组织, 可见少量血管分布; 肌层非常发达, 由外环肌和内纵肌构成; 食道最外层为薄层浆膜(图2 (B1)、(B2), 表1).
2.2.3 胃
胃由黏膜层、黏膜下层、肌层及浆膜层构成(图2(C)). 黏膜层由单层柱状上皮及固有膜构成, 上皮中可见单层柱状细胞紧密排列分布; 固有膜较厚, 其中分布有大量胃腺, 胃腺为单管状腺, 横切面由腺细胞围绕而成(图2(C2)、(C3)). 黏膜下层为疏松结缔组织, 也有少量血管分布(图2(C2)). 肌层属平滑肌, 由内环肌及外纵肌构成, 其中环肌非常发达; 浆膜层薄(图2(C2), 表1).
2.2.4 幽门盲囊
幽门盲囊由黏膜层、黏膜下层、肌层及浆膜构成(图3(D)). 黏膜层向盲囊腔内突起, 形成大量黏膜褶皱(图3(D1)). 黏膜褶皱上皮为单层柱状上皮, 分布有少量杯状细胞, 上皮游离面还分布有明显的纹状缘结构(图3(D3)); 黏膜下层含大量嗜碱性颗粒, HE染色呈深蓝色; 肌层由厚度相近的外纵肌和内环肌构成; 浆膜层薄(图3(D2), 表1).
2.2.5 肠
肠道的前、中、后肠均由黏膜层、黏膜下层、肌层及浆膜层构成(图3(E)、(F)、(G)). 其黏膜上皮为单层柱状上皮, 上皮间分布有杯状细胞, 其游离面具明显的纹状缘结构; 肠道的黏膜下层由疏松结缔组织构成, 较薄, 且与黏膜层分界不清; 肌层由内环肌及外纵肌构成, 其中环肌较纵肌发达; 浆膜层薄(图3(E2)、(E3)、(F2)、(F3)、(G2)、(G3)). 肠道单个横切面内黏膜褶皱数量以前中肠较多, 后肠较少; 黏液细胞的数量由前往后逐渐减少; 而黏膜褶皱高度、黏膜下层厚度及肌层厚度均以前肠和后肠较高, 而中肠较低(图3(E2)、(E3)、(F2)、(F3)、(G2)、(G3), 表1).

3 讨论
3.1 消化道外形与食性的相关性

3.2 消化道各部组织结构特点
棘头梅童鱼舌、食道表面被覆复层扁平上皮, 具有较好耐磨性, 可防止机械损伤. 舌上皮内分布有味蕾, 有助于甄别及选择食物. 食道上皮内含大量黏液细胞, 分泌的黏液可润滑食物, 减少机械摩擦, 这与大黄鱼()[2]、小黄鱼()[3]以及美洲黑石斑鱼()[22]等鱼类相似.
棘头梅童鱼胃黏膜层上皮为单层柱状上皮, 组织学方法未观察到杯状细胞或者其他类型的黏液细胞在上皮中分布, 但是在条石鲷()[23]、波纹唇鱼()[24]、小黄鱼()[3]等多种鱼类的胃上皮中均能观察到杯状细胞或黏液细胞的分布. 有研究报道, 含中性黏多糖的杯状细胞或黏液细胞可在胃黏膜层表层分泌一层稳定的中性黏液物质, 可防止胃酸等强酸性物质对胃黏膜的侵蚀, 进而起到保护胃黏膜的作用[25-26]. 棘头梅童鱼胃中是否含此类黏液细胞, 有待通过组织化学等方法进行检测. 此外, 棘头梅童鱼胃黏膜层中还分布有大量胃腺, 这与大黄鱼()[2]、小黄鱼()[3]、真鲷()[4]等肉食性鱼类相同. 胃腺具有分泌胃蛋白酶原、盐酸等物质的作用, 可促进食物转化为食糜, 并使食物中的蛋白质在胃内进行消化吸收[4].
棘头梅童鱼胃与前肠连接处分布有12~17根指状盲囊, 其组织结构与肠道类似, 可增加肠道消化吸收的表面积, 具有辅助肠道吸收的作用, 因而被认为是由于体腔狭小而不能增加肠长的鱼类以及肉食性鱼类为增加其肠表面积的一种适应性进化[27]. 棘头梅童鱼肠道黏膜层上皮为单层柱状上皮, 上皮游离面具明显的纹状缘结构, 据报道, 纹状缘结构与碱性磷酸酶等消化酶联系紧密[28], 例如在欧洲无须鳕()中, Kozariæ等[29]用组织化学方法检测到碱性磷酸酶在纹状缘部位活性最高; 而在尼罗罗非鱼()中, Tengjaroenkul等[30]检测到麦芽糖酶、氨基肽酶、脂肪酶、非特异性酯酶以及碱性磷酸酶的主要分布区域都在纹状缘上, 这说明纹状缘结构在肠道的消化吸收过程中起非常重要的作用. 棘头梅童鱼前、中肠黏膜褶皱的数量及杯状细胞的密度均高于后肠, 而前、中、后肠的黏膜褶皱高度无显著差异(表1). 据报道, 前、中肠为大部分鱼类肠道发挥消化吸收作用的主要场所, 后肠消化吸收作用较小, 主要起抑菌及促排便等作用[1,31], 本文中的棘头梅童鱼是否也是前中肠消化吸收能力更强, 有待通过组织化学或酶学等方法进行验证.
4 结论
棘头梅童鱼口裂较大, 胃卜型, 含较多幽门盲囊(12~17根), 肠指数较小(0.72±0.1). 舌含味蕾和黏液细胞; 食道粗短, 复层上皮内含大量黏液细胞, 可润滑食物并防止机械损伤; 胃部体积较大, 胃腺发达, 有助于食物的储存及消化吸收; 幽门盲囊结构与肠道类似, 增加了肠道消化吸收的表面积; 肠道在食物的消化吸收方面也起重要作用. 总之, 棘头梅童鱼消化道形态及其组织学结构特点与其肉食食性相适应.
[1] 徐革锋, 陈侠君, 杜佳, 等. 鱼类消化系统的结构、功能及消化酶的分布与特性[J]. 水产学杂志, 2009, 22(4): 49-55.
[2] 林树根, 陈文烈, 钟秀容, 等. 大黄鱼消化道器官显微与亚显微结构[J]. 水产学报, 2002, 26(5):396-401.
[3] 陈寅儿, 郑学斌, 高心明, 等. 小黄鱼()消化道形态与组织学结构特征及其消化酶活性的研究[J]. 海洋与湖沼, 2019, 50(5):1116-1126.
[4] 喻子牛, 孔晓瑜, 孙世春. 真鲷消化道的组织学和形态学研究[J]. 水产学报, 1997, 21(2):113-119.
[5] 陈惠群, 焦海峰, 竺俊全, 等. 龙头鱼的外形特征与消化系统结构[J]. 宁波大学学报(理工版), 2004, 17(4): 389-392.

[7] Hassan A A. Anatomy and histology of the digestive system of the carnivorous fish, the brown-spotted grouper,(Pisces; Serranidae) from the Red Sea[J]. Life Science Journal, 2013, 10(2):2149-2164.
[8] 吴常文, 王伟宏. 浙江近海棘头梅童鱼的分布生物学与资源变动[J]. 海洋渔业, 1991, 13(1):6-10.
[9] 贺舟挺, 薛利建, 金海卫. 东海北部近海棘头梅童鱼食性及营养级的探讨[J]. 海洋渔业, 2011, 33(3):265-273.
[10] 胡艳, 张涛, 杨刚, 等. 长江口近岸水域棘头梅童鱼资源现状的评估[J]. 应用生态学报, 2015, 26(9):2867- 2873.
[11] 黄良敏, 李军, 谢仰杰, 等. 闽江口及其附近海域棘头梅童鱼资源的研究[J]. 台湾海峡, 2010, 29(2):250-256.
[12] Song W, Jiang K J, Zhang F Y, et al. Characterization, molecular cloning, and expression analysis of Ecsit in the spinyhead croaker,[J]. Genetics and Molecular Research: GMR, 2016, 15(1):1-10.
[13] 宋炜, 孟永永, 蒋科技, 等. 棘头梅童鱼七个野生群体遗传多样性的微卫星分析[J]. 水产学报, 2017, 41(1): 31-39.
[14] Andrade I M, Guimarães J P, Rotundo M M, et al. Morphology of the digestive tract of the Whitemouth croaker(Desmarest, 1823) (Perciformes: Sciaenidae)[J]. Acta Zoologica, 2017, 98(2): 136-143.
[15] 王迎春, 周勤, 段晓英. 八种海产硬骨鱼类消化系统的比较解剖研究[J]. 海洋湖沼通报, 1997(3):46-51.
[16] 闻海波, 马学艳, 徐跑, 等. 淡水石首鱼幼鱼外部形态与内部解剖学特征[J]. 中国水产科学, 2018, 25(6):14- 24.
[17] 毕冰, 孙中武, 毛天强, 等. 鲤、鲢、鳙、草鱼消化道结构与食性的研究[J]. 水产学杂志, 2011, 24(1):26-29.
[18] 杨元昊, 王立新, 李学智, 等. 兰州鲇与鲇消化系统的形态学及组织学比较研究[J]. 水生生物学报, 2017, 41 (1):174-181.
[19] 罗江波, 戴海平, 龙玲利, 等. 光唇鱼消化道的形态结构特征[J]. 生物学杂志, 2013, 30(6):59-61; 65.
[20] Yang L, Fang J, Peng X, et al. Study on the morphology, histology and enzymatic activity of the digestive tract ofHerzenstein[J]. Fish Physiology and Biochemistry, 2017, 43(4):1175-1185.
[21] 殷江霞, 张耀光, 李萍, 等. 华鲮消化道组织学与组织化学的初步研究[J]. 淡水渔业, 2005, 35(6):7-10.
[22] 李海燕, 竺俊全, 陈飞, 等. 美洲黑石斑鱼消化道的形态结构[J]. 生物学杂志, 2011, 28(4):31-34; 46.
[23] 王健鑫, 石戈, 李鹏, 等. 条石鲷消化道的形态学和组织学[J]. 水产学报, 2006, 30(5):618-626.
[24] 区又君, 廖光勇, 李加儿. 波纹唇鱼消化道的形态学和组织学[J]. 热带海洋学报, 2012, 31(6): 83-89.
[25] 彭迁迁, 权梦雪, 罗莹, 等. 鳜鱼消化道黏液细胞和6种酶的组织化学定位[J]. 激光生物学报, 2016, 25(2): 161-169.
[26] Wilson J M, Castro L F C. Morphological diversity of the gastrointestinal tract in fishes[J]. Fish Physiology, 2010, 30: 1-55.
[27] 尾崎久雄. 鱼类消化生理(下册)[M]. 李爱杰, 沈宗武, 译. 上海: 上海科学技术出版社, 1983:331-414.
[28] Nasruddin N S, Azmai M N A, Ismail A, et al. Histological features of the gastrointestinal tract of wild Indonesian shortfin eel,, (McClelland, 1844), captured in Peninsular Malaysia[J]. The Scientific World Journal, 2014, 2014:312670.
[29] Kozariæ Z, Kužir S, Nejedli S, et al. Histochemical distribution of digestive enzymes in hake,L. 1758[J]. Veterinarski Arhiv, 2004, 74(4): 299-308.
[30] Tengjaroenkul B, Smith B J, Caceci T, et al. Distribution of intestinal enzyme activities along the intestinal tract of cultured Nile tilapia,L[J]. Aquaculture, 2000, 182(3/4):317-327.
[31] 刘亚秋, 李新辉, 王志坚. 大鳞副泥鳅肠道黏液细胞及消化酶活性分布特征研究[J]. 水生生物学报, 2017, 41(5):1048-1053.
Morphology and histological structure of digestive tract of
XU Wantu, TAO Shunshun, ZENG Xiao, XU Fangjun, CHEN Yu
( Gangwan Aquatic Fingerlings Limited Company of Xiangshan, Xiangshan 315700, China )
The morphological and histological features of digestive tract ofwere determined in the present study. The results show that the digestive tract consisted of five parts including oropharyngeal cavity, esophagus, stomach, pyloric caecum and intestine. The oropharyngeal cavity contained a short tongue and is formed by the projecting part of basihyoid covered by mucosa. The mucosa epithelium of tongue was stratified squamous epithelium and contained several taste buds or mucous cells. The esophagus, stomach, pyloric caeca and intestine were consisted of the mucosa, sub mucosa, muscle layer and serosa tissues. The esophagus was short and wide and present with many mucous cells in the stratified squamous epithelium. The stomach was λ-shaped and abundant in gastric glands. There were 12-17 finger-shaped pyloric caeca located to the connective part of stomach and intestine. The histological structure of pyloric caecum was similar to the intestine and covered with single columnar epithelium that contained a small number of mucous cells. In addition, the intestine was short and S-shaped, with the intestine index being 0.72±0.10. This study reveals that the features of morphology and histological structure of digestive tract ofwas in consistency with its feeding habits and digestive functions.
; anatomy; histology; feeding habit
S917.4
A
1001-5132(2022)02-0015-07
2021−10−17.
宁波大学学报(理工版)网址: http://journallg.nbu.edu.cn/
徐万土(1973-), 男, 浙江象山人, 工程师, 主要研究方向: 海水鱼类繁育与遗传育种. E-mail: xuwantu@163.com
(责任编辑 章践立)
