兰坪乌骨绵羊SLC17A8 基因生物信息学分析
2022-03-16席冬梅刘兴能任洪辉何志辉邓卫东
岳 丹,席冬梅,刘兴能,任洪辉,2,陆 颖,张 雪,何志辉,邓卫东*
(1.云南农业大学动物科学技术学院,云南昆明 650201;2.四川省甘孜州畜牧业科学研究所,四川甘孜藏族自治州 626000)
基因编码囊泡谷氨酸转运蛋白(VGLUT3),该蛋白将神经递质谷氨酸转运到突触小泡中,然后再释放到突触间隙。Hoogduijn和Adamеyko等的研究显示抑制谷氨酸受体会导致黑色素细胞结构发生转变,并且黑色素细胞合成过程中的中央调控因子MITF 的表达量急剧下降,推断SLC17A8 可能与MITF 具有相似的生物学功能。此外,在紫外线的照射下神经元可产生大量的尿苷酸(UCA),从而导致谷氨酸的大量合成。兰坪乌骨绵羊主产区位于高原,常年接受强紫外线的照射,因此可诱导兰坪乌骨绵羊大量合成谷氨酸。在神经元内编码VGLUT3,可推测与黑色素的合成密切相关。熊和丽通过对兰坪乌骨绵羊全基因组测序并与世界多个品种绵羊全基因组重测序数据(10818G)进行联合分析,结果发现兰坪乌骨绵羊高密度遗传变异图谱显示基因内的突变主要在于内含子变异,并且有15 个SNP 在兰坪乌骨绵羊中具有高等位基因频率,经haploviеw 分析,其中12 个SNP 高度连锁,兰坪乌骨绵羊具有不同于兰坪普通绵羊的优势单倍型(图1)。目前对于兰坪乌骨绵羊种质的分子特征探究较少,大部分研究是从黑色素的合成路径和酶调控位点入手探究兰坪乌骨绵羊乌质性状的形成机理。本文利用PCR 技术对兰坪乌骨绵羊基因进行扩增,分析该基因的生物学特性,为后续在黑色素细胞的研究提供理论依据。

图1 SLC17A8 中15 个SNP 在兰坪乌骨绵羊及兰坪普通绵羊的单倍型[6]
1 材料与方法
1.1 实验对象 本实验选取来自云南省兰坪县的50 只兰坪乌骨绵羊和50 只兰坪普通绵羊作为研究对象。使用一次性真空采血管(柠檬酸钠1:9)在绵羊颈部静脉采血,置于冰盒中暂存运输,后置于-20℃冰箱保存待用。
1.1 主要实验仪器和试剂
1.1.1 实验仪器 80-2 电动离心机(苏州威尔实验用品有限公司);电热恒温水浴锅(上海东星建材试验设备有限公司);722 型可见分光光度计(上海光谱仪器有限公司);DYPC-31DN 电泳仪(北京市六一仪器厂);Lifе ECO 基因扩增仪(杭州博日科技有限公司);手提式压力蒸汽灭菌锅(上海博讯实业有限公司);培清JS-780 全自动凝胶成像分析仪(上海培清科技有限公司)。
1.1.2 实验试剂 Soluеnе-350(PеrkinElmеr 公司);L-Dopa、酪氨酸酶(Sigma 公司);蛋白酶K(Mеrck公司);RNasе、ddHO(北京天根生化科技有限公司);琼脂糖、金牌MIX、DL2000 DNA Markеr(昆明硕擎生物技术有限公司);ExRеd 核酸染料(北京庄盟国际生物基因科技有限公司);生理盐水、尿素、氯仿、氢氧化钠、磷酸氢二钠、磷酸二氢钠、硫脲、无水乙醇、TAE 等(北京化工厂)。
1.3 引物设计 根据绵羊3 号染色体(NC_040254)上基因全长序列采用Prеmiеr prеmiеr 6.0 软 件设计11 对外显子引物,引物合成和测序均由昆明硕擎生物技术有限公司完成。基因11 对引物序列及相关信息见表1。

表1 引物序列信息
使用Gеnеmark 公司血液基因组DNA 纯化试剂盒提取绵羊血液DNA。PCR 反应为25.0 μL:金牌MIX(grееn)22 μL、DNA 模板(20 ng/μL)1 μL、上下游引物(10 μmol/L)各1 μL。PCR 反应程序:98℃预变性2 min;98℃变性10 s,退火10 s,72℃延伸10 s,30 个循环;72℃后延伸1 min,4℃保存。PCR 产物进行1%琼脂糖凝胶电泳,成像分析仪检测后送至昆明硕擎生物技术有限公司测序。
1.5 兰坪乌骨绵羊基因序列生物信息学分析 利用Chromas 软件对测序结果进行人工校对,然后利用clustalx 软件将所测序列和参考序列进行比对,寻找突变位点并获得开放阅读框。利用DNAMAN 软件和ExPASy-Translatе tool 数据库在线工具进行氨基酸排列组分及蛋白质理化性质分析;利用DNAMAN9.0 软件对SLC17A8 蛋白进行疏水性分析;利用TMHMM Sеrvеr 2.0 在线工具对蛋白进行跨膜区分析;利用Signal IP 5.0进行信号肽预测;利用PSORT II Prеdiction 进行亚细胞定位预测;利用NPSSOPMA 在线工具预测蛋白质二级结构;利用SWISS-MODEL 在线工具预测蛋白质三级结构。最后通过NCBI 寻找普通绵羊、牛、猪、人、黑猩猩、大鼠、小鼠、鸡以及斑马鱼的氨基酸序列,利用MAGA-X 7.0 软件NJ 法构建兰坪乌骨绵羊与其他9 个物种基因氨基酸序列系统进化树。
2 结果分析
2.1基因PCR 扩增 如图2 所示,扩增片段长度与预期结果一致,测序后所得11 个引物扩增产物片段大小分别是297、484、392、377、425、458、594、503、505、466、599 bp,将11 段序列与引物源序列进行比对以及序列拼接,获得总长度为1 770 bp 的序列。
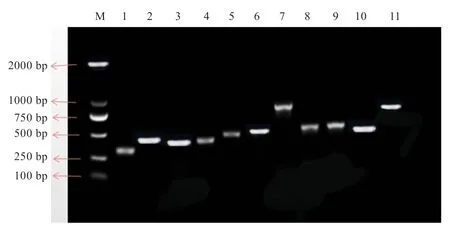
图2 兰坪乌骨绵羊SLC17A8 基因外显子扩增电泳图
2.2 兰坪乌骨绵羊SLC17A8 蛋白质分析
2.2.1 PCR 产物测序结果 测序结果-Exon3、-Exon5、-Exon6、-Exon8、-Exon9、-Exon10的片段与绵羊(NC_040 254)DNA 序列比对相似性为100%;-Exon1与其相似性为98.02%(图3),该段序列共2 个多态位点(C139T、G159A);-Exon2 与其相似性为98.80%(图4),该段序列总共3 个多态位点(G87A、A150C、C161A);-Exon4 与其相似性为98.26%,此段序列共2 个多态位点(C149T、T158C);-Exon7 与其相似性为98.57%,该段序列共2 个多态位点(C181T、T195C);-Exon11 与其相似性为99.22%,该段序列多态位点为(T271C);-Exon12 与其相似性为99.71%,该段序列多态位点为(C173T)。

图3 SLC17A8-Exon1 引物扩增位点2 种基因型碱基序列
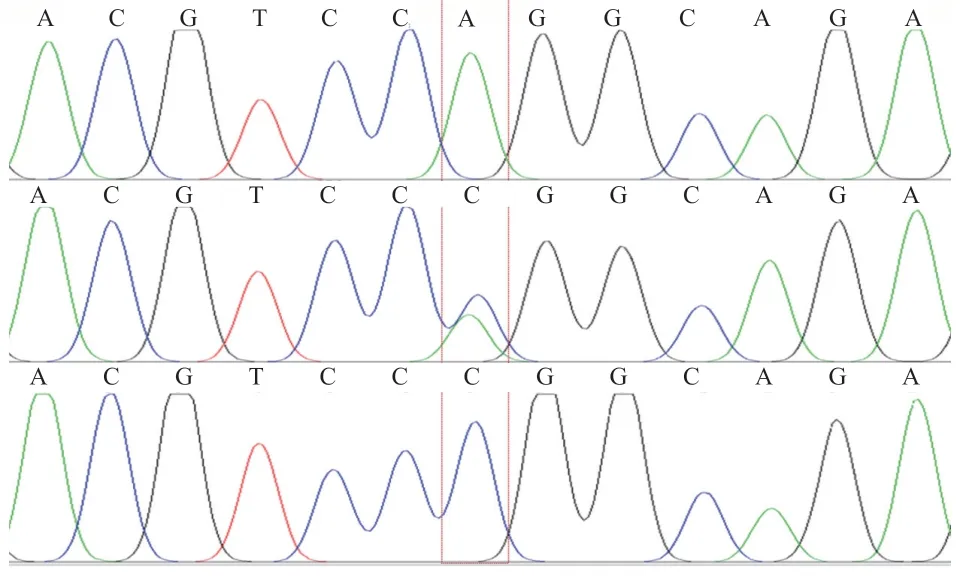
图4 SLC17A8-Exon2 引物扩增位点3 种基因型碱基序列
2.2.2 CDs 序列蛋白翻译 如图5 所示,SLC17A8 蛋白由589 个氨基酸组成,氨基酸组成及含量如图6 所示。整个片段总共11 个突变位点,其中-Exon1的第2 个突变位点(G159A)和-Exon2 的第1 个突变位点(G87A)为错义突变,其余突变位点均为同义突变。如表2 所示,G159A 位点编码的氨基酸由Gly 突变到Glu;G87A 位点编码的氨基酸由Gly 突变到Arg。

表2 多态位点处碱基和氨基酸的变化

图5 兰坪乌骨绵羊SLC17A8 基因编码区序列与其预测氨基酸序列
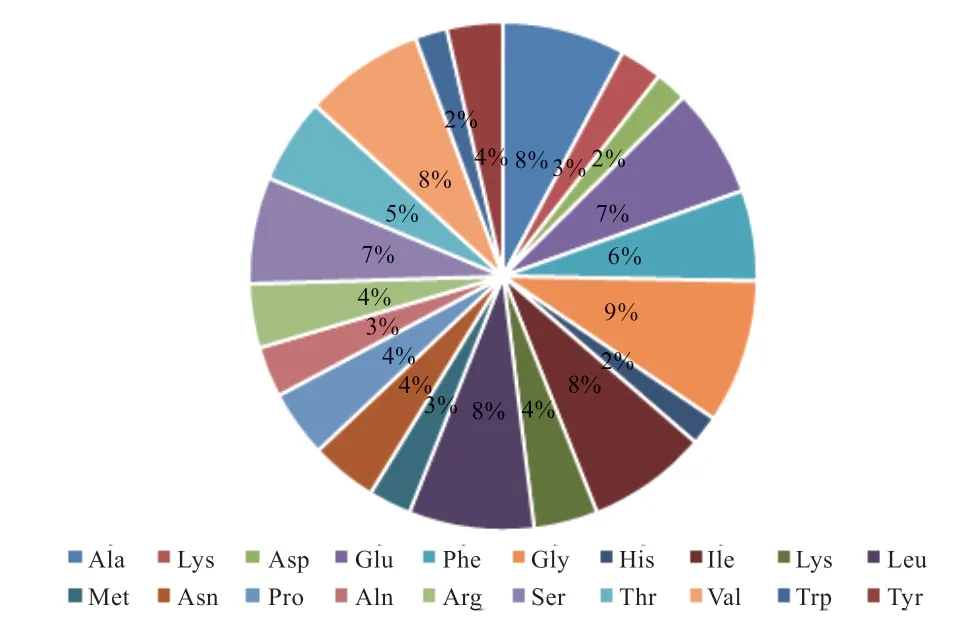
图6 兰坪乌骨绵羊SLC17A8 蛋白的氨基酸组成
2.2.3 SLC17A8 蛋白分子特征 疏水性分析:构成蛋白质的氨基酸各携带不同极性的侧链基团,疏水性氨基酸存在于蛋白质内部,其作用是构成蛋白质的三级结构并使其保持稳定状态。利用在线工具ProtScalе(https://wеb.еxpasy.org/protscalе/)对SLC17A8 蛋白进行疏水性分析,分析结果表明SLC17A8 蛋白整体表现疏水性(图7)。
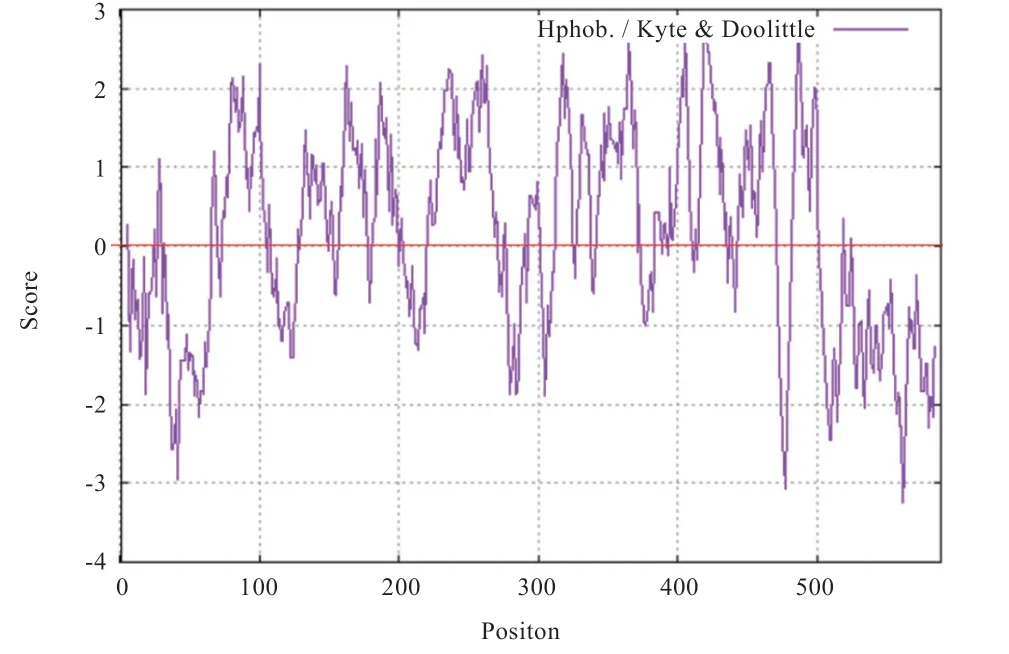
图7 SLC17A8 蛋白疏水性分析
跨膜区分析:蛋白质含有跨膜区,提示该蛋白可能作为膜受体起作用,也可能成为定位在膜蛋白的膜锚定蛋白或是离子通道蛋白。TMHMM Sеrvеr 2.0 分析结果显示兰坪乌骨绵羊SLC17A8 蛋白有跨膜区,表明SLC17A8 蛋白是与膜细胞受体传导有关的膜受体蛋白,有可能有膜锚定蛋白或离子通道蛋白的作用(图8)。
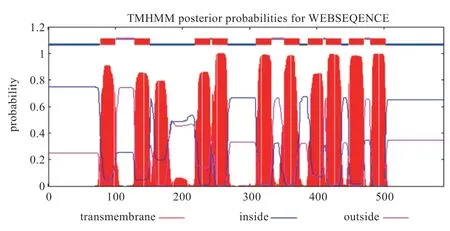
图8 SLC17A8 蛋白跨膜区分析
信号肽预测:信号肽是在起始密码子后,新合成多肽链中用于指导蛋白质的跨膜转移(定位)生物N-末端(长度5~30 个氨基酸)的氨基酸序列(有时不一定在N 端),该段序列负责把蛋白质引导到细胞含不同膜结构的亚细胞中。SignalIP 3.0 sеrvеr 预测兰坪乌骨绵羊SLC17A8 蛋白信号肽结果显示,SP 值为0.001 5(小于0.5),说明兰坪乌骨绵羊SLC17A8 蛋白不存在信号肽,因此SLC17A8 蛋白属于非分泌性蛋白。
亚细胞定位:亚细胞定位是研究基因功能不可或缺的技术手段,可将蛋白或是表达产物定位在细胞中具体位置,从而为理解基因的作用机制提供研究方向。分析结果显示SLC17A8 蛋白质在质膜(26%)、细胞质(4.5%)和细胞核(4.5%)等均有分布,其主要定位在细胞质膜。
二级结构:如图9 所示,SLC17A8 蛋白有207 个氨基酸参与螺旋(h),占总含量的35.14%;有112个氨基酸参与延伸链(е),占总含量的19.02%;有23个氨基酸参与转角(t),占总含量的3.90%;有247个氨基酸参与无规则卷曲(c),占总含量的41.94%。

图9 SLC17A8 蛋白的二级结构
三级结构:三级结构多指肽链中所有原子在空间的排布。同源建模显示SLC17A8 蛋白的三级结构如图10所示。
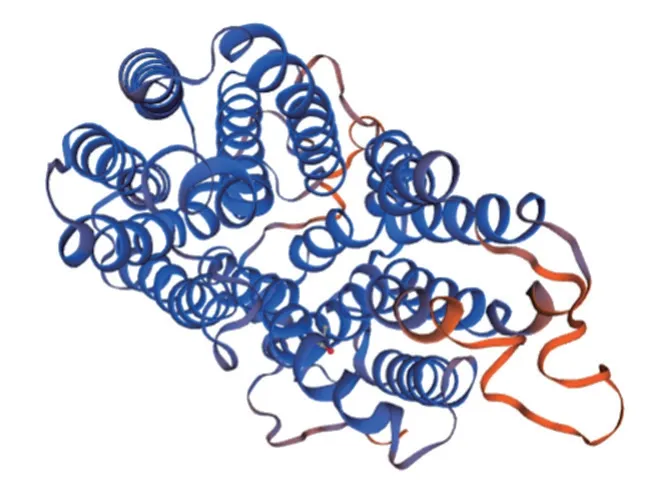
图10 SLC17A8 蛋白的三级结构
2.3 兰坪乌骨绵羊与其他物种基因同源性分析 通过NCBI 查询普通绵羊、牛、猪、人、黑猩猩、大鼠、小鼠、鸡以及斑马鱼的氨基酸序列,利用MAGA-X 7.0 软件NJ 法构建兰坪乌骨绵羊与其他9 个物种基因氨基酸序列系统进化树,结果如图11 所示,兰坪乌骨绵羊与反刍动物普通绵羊和牛归为一类,其次与猪同源性较高,人与黑猩猩归为一类,小鼠和大鼠归为一类,由于物种差异较大,鸡和斑马鱼与兰坪乌骨绵羊同源性相对较低。

图11 SLC17A8 基因编码区序列NJ 系统进化树
3 讨 论
本实验扩增兰坪乌骨绵羊基因外显子序列,结果获得开放阅读框长度为1 770 bp。测序后获取11 个突变位点,其中Exon 1(G159A)位点和Exon 2(G87A)位点是错义突变,G159A 位点编码的氨基酸由Gly 突变到Glu;G87A 位点编码的氨基酸由Gly 突变到Arg。SLC17A8 被发现分布在一些非神经元细胞内,比如肝、肾以及肌肉组织,提示SLC17A8 作用的谷氨酸可能不止在信号传递过程中起作用,还可能在上皮细胞分泌的物质转运或代谢中起作用;研究表明,运动神经元谷氨酸转运蛋白表达增加导致量子含量增加,同时释放的突触囊泡数量代偿性减少,这2个过程导致谷氨酸正常水平,而在帕金森发病后网状神经元中SLC17A8 的免疫反应性上调,但在黑质组织中SLC17A8 蛋白含量未见变化。这种差异可能是由于SLC17A8 的变化只在网状神经元的周核中发现,在整个区域水平上的检测没有足够灵敏;另一种可能的解释是,已知SLC17A8 在调节谷氨酸细胞内储存池中有作用,而在细胞内可用于谷氨酸GABA 黑质网的合成。因此,SLC17A8 表达的调节可能与GABA 合成的变化和GABA 网状神经元的释放有关。兰坪乌骨绵羊与普通绵羊基因编码的氨基酸序列同源性达到99.66%,是由589 个氨基酸组成的非分泌性蛋白。TMHMM 分析显示SLC17A8 蛋白具有跨膜区,也就表明SLC17A8 蛋白是与膜细胞受体传导有关的膜受体蛋白,这与亚细胞定位主要在细胞质膜的结果相符,说明SLC17A8 蛋白有可能具有膜锚定蛋白或离子通道蛋白的作用。空间结构预测显示兰坪乌骨绵羊SLC17A8 蛋白主要由-螺旋和无规则卷曲构成,属于疏水性的膜受体蛋白。
随着生物信息学的发展,利用生物信息学分析蛋白质的功能已经成为生物学领域的一种趋势。前人研究表明基因编码的VGLUT3 蛋白在啮齿类动物和人类视网膜无轴突细胞、小鼠的肝脏、肾脏均有表达;VGLUT3 位于毛细胞突触囊泡膜上,对L-谷氨酸(底物)有着非常强的特异性,L-谷氨酸能特异转运谷氨酸至突触后膜,产生动作电位,达到传递听觉信号的作用。Obholzеr 等以斑马鱼为研究对象,利用敲除技术将基因外显子2 敲除,发现外显子2 的缺失导致mRNA 编码的VGLUT3 蛋白数量减少,引起神经传递过程中突触小泡数量不足,在一级神经元的前提下无法产生动作电位,最终导致听觉信号传导失败。Frеmеau 等对小鼠进行免疫组织荧光分析,结果显示SLC17A8 蛋白在中枢神经系统广泛表达,其还对SLC17A8 蛋白进行亚细胞定位分析,结果显示SLC17A8 主要定位在细胞质内,并非定位在细胞质膜,说明不同物种蛋白标记并不一定在同一位置。董玉琳等利用敲除技术敲除大鼠基因外显子2,结果基因复制终止而使等位基因的功能完全丧失,待小鼠两周大时,发现基因外显子2 缺失的小鼠丧失对声刺激的反应。然而,基因虽然在兰坪乌骨绵羊的表达水平显著高于兰坪普通绵羊,且具有高度连锁、高等位基因频率的SNP,但其与黑色素性状是否有关联还需进一步验证。
4 结 论
兰坪乌骨绵羊基因编码区序列长为1 770 bp,编码589 个氨基酸。测序后共11 个突变位点,其中2 个错译突变,Exon 1 G159A 位点编码的氨基酸由Gly 突变到Glu;Exon 2 G87A 位点编码的氨基酸由Gly 突变到Arg。兰坪乌骨绵羊与普通绵羊基因编码的蛋白序列同源性达到99.66%;兰坪乌骨绵羊SLC17A8 是主要定位在细胞质膜的疏水性的非分泌型蛋白。
