植物生长调节剂对花后渍水遮阴小麦籽粒淀粉合成和干物质积累的影响
2022-03-16王贝贝徐旭赵艳侯君佑杨丽胡正马尚宇樊永惠黄正来张文静
王贝贝 徐旭 赵艳 侯君佑 杨丽 胡正 马尚宇 樊永惠 黄正来 张文静
摘要: 為探明喷施植物生长调节剂6-苄氨基腺嘌呤(6-BA)和水杨酸(SA)对小麦花后渍水遮阴胁迫后籽粒淀粉合成和干物质积累的影响,以渍水敏感型品种皖麦52和渍水迟钝型品种扬麦18为材料,人工模拟花后7 d、11 d和15 d 3个渍水遮阴时长,于处理结束当天喷施6-BA和SA,研究喷施植物生长调节剂对小麦籽粒淀粉合成关键酶活性、淀粉积累、干物质积累及产量的影响。结果表明,渍水遮阴后喷施6-BA和SA能不同程度上缓解渍水遮阴对小麦籽粒淀粉葡萄糖焦磷酸化酶(AGPase)、颗粒结合态淀粉合成酶(GBSS)、可溶性淀粉合成酶(SSS)和淀粉分支酶(SBE)活性造成的影响,对籽粒SSS活性修复的效果最为显著,其次是AGPase和SBE,GBSS活性受外源6-BA的影响较小。与对照相比,喷施6-BA和SA提高了淀粉积累速率,增加了淀粉的最终积累量,延长了花后渍水遮阴后小麦灌浆持续时间,提高了平均灌浆速率,增加了千粒质量和花后渍水遮阴7 d、11 d处理的穗粒数,减轻了花后渍水遮阴对小麦产量的不利影响。另外,两种植物生长调节剂间比较,6-BA的修复效果要优于SA。喷施6-BA和SA均能缓解花后渍水遮阴对小麦籽粒淀粉合成关键酶活性造成的伤害,有利于籽粒淀粉合成和干物质积累。
关键词: 小麦;渍水遮阴;6-苄氨基腺嘌呤(6-BA);水杨酸(SA);淀粉;干物质积累
中图分类号: S512.101 文献标识码: A 文章编号: 1000-4440(2022)01-0009-11
Abstract: This study aimed to investigate the effects of spraying plant growth regulator 6-benzylamino adenines (6-BA) and salicylic acid (SA) on starch synthesis and dry matter accumulation of wheat grains under waterlogging and shading after anthesis. The waterlogging-sensitive variety Wanmai 52 and the waterlogging dull variety Yangmai 18 were used as materials, and three waterlogging shading periods of 7 d, 11 d and 15 d after anthesis were artificially simulated. The 6-BA and SA were sprayed on the day of the end of the treatment. The effects of spraying plant growth regulators on the activities of key enzymes in starch synthesis, starch accumulation, dry matter accumulation and yield in wheat were studied. The results showed that spraying 6-BA and SA after waterlogging and shading could alleviate the effects of waterlogging and shading on the activities of starch glucose pyrophosphorylase (AGPase), granule-bound starch synthase (GBSS), soluble starch synthesis (SSS) and starch branching enzyme (SBE) in wheat grains to varying degrees. Among them, the effect of repairing SSS activity in the grain was the most significant, followed by AGPase and SBE. The activity of GBSS was less affected by exogenous 6-BA. Compared with the control, spraying 6-BA and SA increased the rate of starch accumulation, increased the final accumulation of starch, extended the duration of wheat filling after waterlogging and shading, raised the average filling rate, added the thousand-grain weight and the number of grains per spike treated with shading and waterlogging for 7 d and 11 d, alleviated the adverse effects of waterlogging and shading on wheat yield. In addition, the remediation effect of 6-BA was better than that of SA. Spraying 6-BA and SA can alleviate the damage caused by waterlogging and shading to the activities of key enzymes in starch synthesis in wheat grains, which is beneficial to starch synthesis and dry matter accumulation.
Key words: wheat;waterlogging and shading;6-benzylamino adenine (6-BA);salicylic acid (SA);starch;dry matter accumulation
近年来,随着极端气候频繁发生,渍害已成为长江中下游地区稻茬小麦生产的主要限制因子[1-3],小麦常在生育后期遭受连续阴雨,严重影响了小麦产量[4-5]。同时持续性降水带来的弱光效应,使小麦在生育后期受寡照影响严重[6-8]。因此,针对长江中下游地区小麦生育后期遭受连阴雨的现象,探索有效缓解伤害的栽培技术措施具有重要意义。
小麦生长发育后期主要是籽粒形成时期,淀粉是籽粒的主要成分,占籽粒干质量的65%左右,与产量和品质的关系极为密切[9]。淀粉的积累过程受一系列淀粉合成有关酶的调控,主要包括葡萄糖焦磷酸化酶(AGPase)、颗粒结合态淀粉合成酶(GBSS)、可溶性淀粉合成酶(SSS)、淀粉分支酶(SBE)等[10]。研究结果表明,小麦在花后遭受渍水遮阴时,淀粉积累过程中关键酶的活性受到抑制,导致淀粉积累的平均速率降低,显著降低了籽粒中淀粉含量,改变了淀粉组分,使淀粉的品质发生变化[11-12]。在作物遭受非生物胁迫时,通常采用喷施植物生长调节物质等措施来减轻逆境对植物的影响。小麦开花期在渍水前喷施6-BA可以降低旗叶丙二醛含量,提高旗叶光合速率,延缓叶片衰老,减少产量损失[13-14]。在小麦开花期遮阴前喷施6-BA能够延缓旗叶的衰老,增加花后光合产物,有效提高产量[6]。李建坤等[15]研究结果表明,在玉米苗期淹水前、后各喷施一次6-BA可以有效改善玉米各器官超氧化物歧化酶(SOD)活性,缓解受涝叶片的膜脂过氧化,提高植株的耐渍性。Larkindale等[16]发现,在拟南芥体内添加SA可以缓解高温带来的氧化损伤,提高耐热性。在小麦幼苗期喷施SA能在一定程度上缓解水分胁迫对叶片的损伤[17]。此外,在小麦开花期喷施SA能够增加淀粉的积累量,有利于籽粒质量的提高[18]。
目前有关喷施6-BA、SA缓解逆境伤害方面的研究多集中在水分胁迫、高温胁迫等单一胁迫方面,且研究内容也大多是关于光合特性、产量等方面,从淀粉合成及干物质积累方面研究小麦开花期渍水遮阴后喷施植物生长调节剂缓解效应的报道较少。本研究拟选用渍水敏感型和迟钝型2个小麦品种,分析渍水遮阴胁迫后喷施6-BA、SA对小麦籽粒淀粉合成关键酶活性、淀粉积累、干物质积累及产量形成的影响,为探索有效缓解小麦花后受连阴雨伤害的栽培技术措施提供理论依据。
1 材料与方法
1.1 试验材料与试验设计
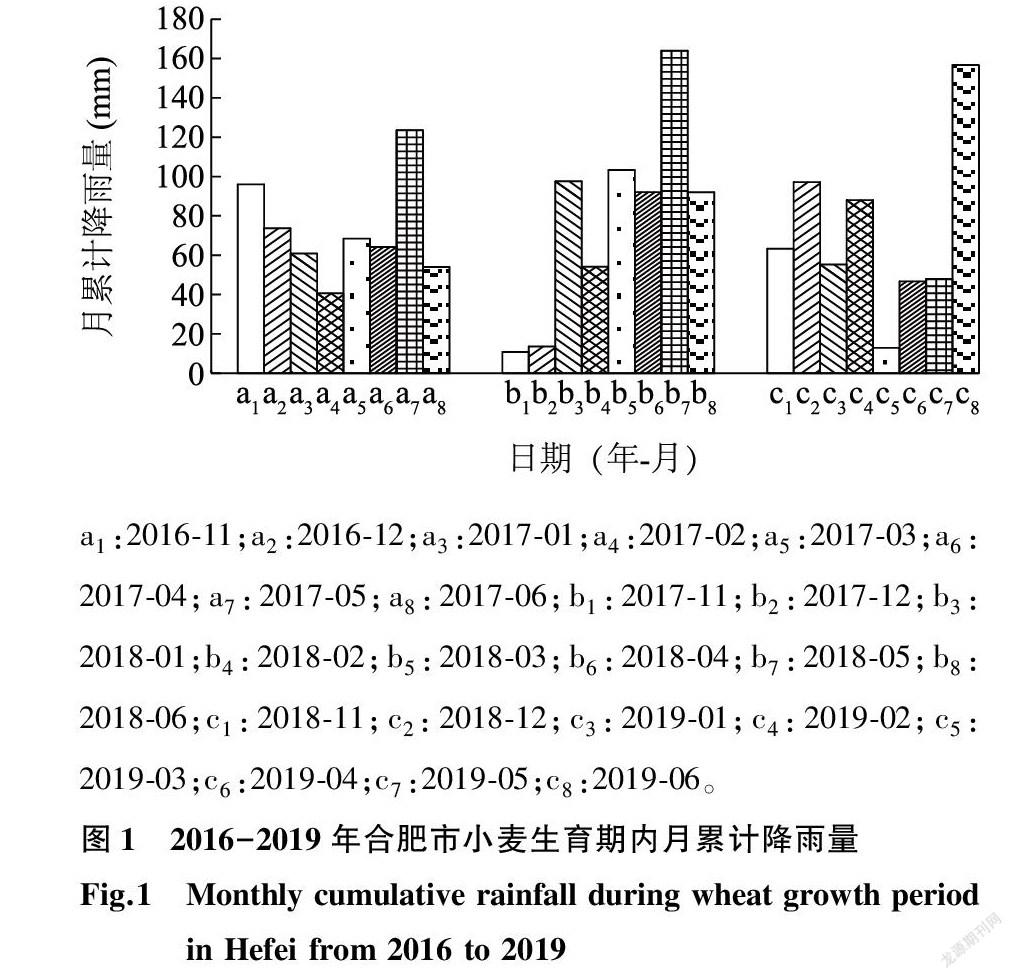
试验于2018年10月-2019年6月在安徽农业大学校内试验基地农萃园(31°52′N,117°17′E)进行,试验点属亚热带季风湿润气候区,年降水量约1 000 mm,试验地2016-2019年小麦生育期内月累计降雨量情况见图1。根据本课题组前期研究结果,选用2个耐渍能力不同的小麦品种(渍水迟钝型品种扬麦18和渍水敏感型品种皖麦52),采用盆栽+池栽的种植方式,盆钵直径28 cm、高26 cm。试验用土取自0~20 cm耕作层,土壤中有机质含量为15.51 g/kg,全氮含量为0.91 g/kg,速效氮、速效磷和速效钾含量分别为102.93 mg/kg、20.82 mg/kg和150.52 mg/kg,过筛后每盆填土10.0 kg。播种前每盆施有机肥75.0 g、纯氮2.3 g、P2O5 2.7 g、K2O 4.5 g,于小麦拔节期每盆追施纯氮1.1 g。试验播种日期为2018年10月31日,每品种种植180盆。试驗设置7 d、11 d和15 d 3个渍水遮阴时长,分别记为WS7、WS11和WS15。在渍水遮阴结束当天喷施6-BA、SA和等量蒸馏水,每个品种共9个处理,相同处理埋于同一试验池中,试验池长6 m,宽4 m,深1.2 m,盆上沿与池面平齐,齐苗后每盆留长势均匀的麦苗8株,田间管理与高产栽培要求一致。
于小麦开花后进行渍水遮阴处理,渍水期间保持水层高出盆钵内土面0.5~1.0 cm,遮阴处理采用固定式铝合金拱棚上铺设开放式遮阳网(50%透光率)进行,遮阳网下沿距地表0.5 m,保证通风良好。根据前期试验筛选结果,选用15 mg/L的6-BA和250 mg/L的SA,在渍水遮阴结束当日喷施,喷施量以叶片均匀沾满药水为准,以喷施等量蒸馏水的植株作为对照(CK)。3个渍水遮阴处理时间段(7 d、11 d、15 d)内降雨时不作遮雨处理;7 d和11 d渍水遮阴处理结束后用透明塑料薄膜作遮雨处理,并保持通风良好,确保不同渍水时长处理之间的差异性;当3个渍水遮阴处理全部结束后(15 d后),不作遮雨处理。
1.2 样品的获取
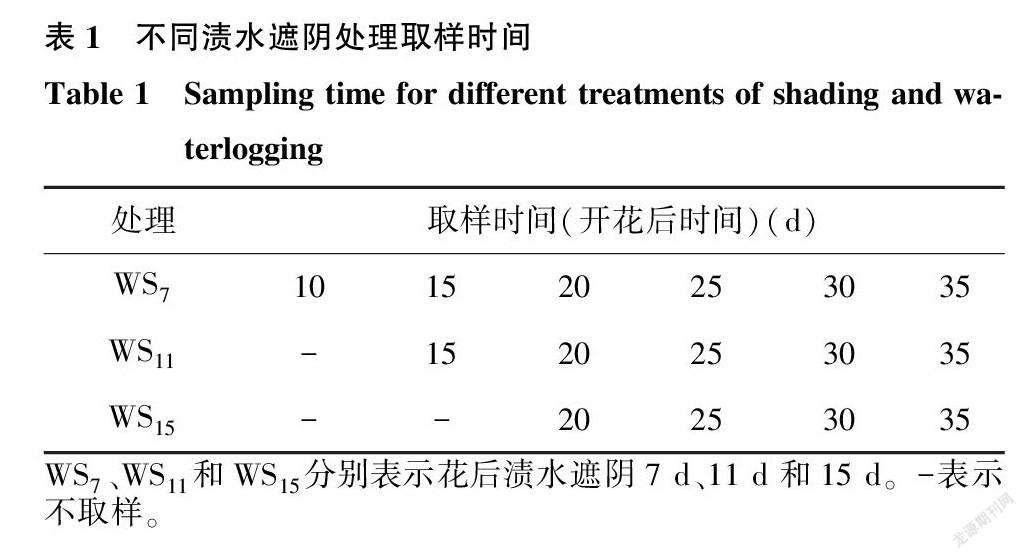
在小麦开花时,各处理中选择同一天开花、长势均匀的小麦单茎进行标记,从花后10 d开始取样,每隔5 d于各处理中随机取大小一致的麦穗8~10个(各处理取样时间见表1),分别用于测定淀粉合成相关酶活性以及淀粉和干物质积累。
1.3 测定项目及方法
1.3.1 籽粒淀粉合成关键酶活性 酶液的提取参照程方民等[19]的方法。AGPase活性测定参照Smyth等[20]的方法,SSS和GBSS活性测定参照Nakamura等[21]的方法,SBE活性测定参照李太贵[22]等的方法。
1.3.2 淀粉含量 参照何照范[23]的双波长比色法,根据Logistic方程Y=K/(1+eA+Bt)(Y为籽粒淀粉含量,t为开花至观察时的天数,A、B为参数,K为拟合的淀粉最大积累量)拟合淀粉积累过程,得到淀粉积累相关特征参数:淀粉积累持续期(T),T=(4.595 12+A)/(-B);淀粉积累活跃期(D),D=[ln(1/9)-A]/B;淀粉平均积累速率(Rmean),Rmean=K/D;淀粉最大积累速率(Rmax),Rmax=-KB/4。
1.3.3 籽粒干物质积累速率 从各处理中取小麦穗15~20个,105 ℃杀青15 min,60 ℃烘至恒质量后剥出籽粒称质量,用Logistic方程W=C/(1+eZ+Mn)(W为籽粒千粒质量,n为开花至观察时的天数,Z、M为参数,C为拟合的最大千粒质量)拟合籽粒灌浆过程,对该方程进行一阶求导,得到籽粒积累速率方程,进而推导出以下参数:灌浆持续期(N,籽粒质量达99%所需天数)N=(4.595 12+Z)/-M ;最大灌浆速率出现时间(N3),N3=N-N1-N2;灌浆渐增期(Nmax),Nmax=-Z/M;灌浆快增期( N1), N1=(Z-1.317)/M;灌浆缓增期(N2),N2=(Z+1.317)/M-(Z-1.317)/M。
1.3.4 产量及其构成因素 待小麦成熟后,每处理取未取样的小麦10盆,用于测定穗粒数、千粒质量并计算出单株产量。
1.4 数据处理
采用SPSS 22.0软件进行方差分析,Dunca 法对测定各项数据进行多重比较,用Origin 2017 作图。
2 结果与分析
2.1 6-BA和SA对花后渍水遮阴小麦籽粒淀粉合成关键酶活性的影响
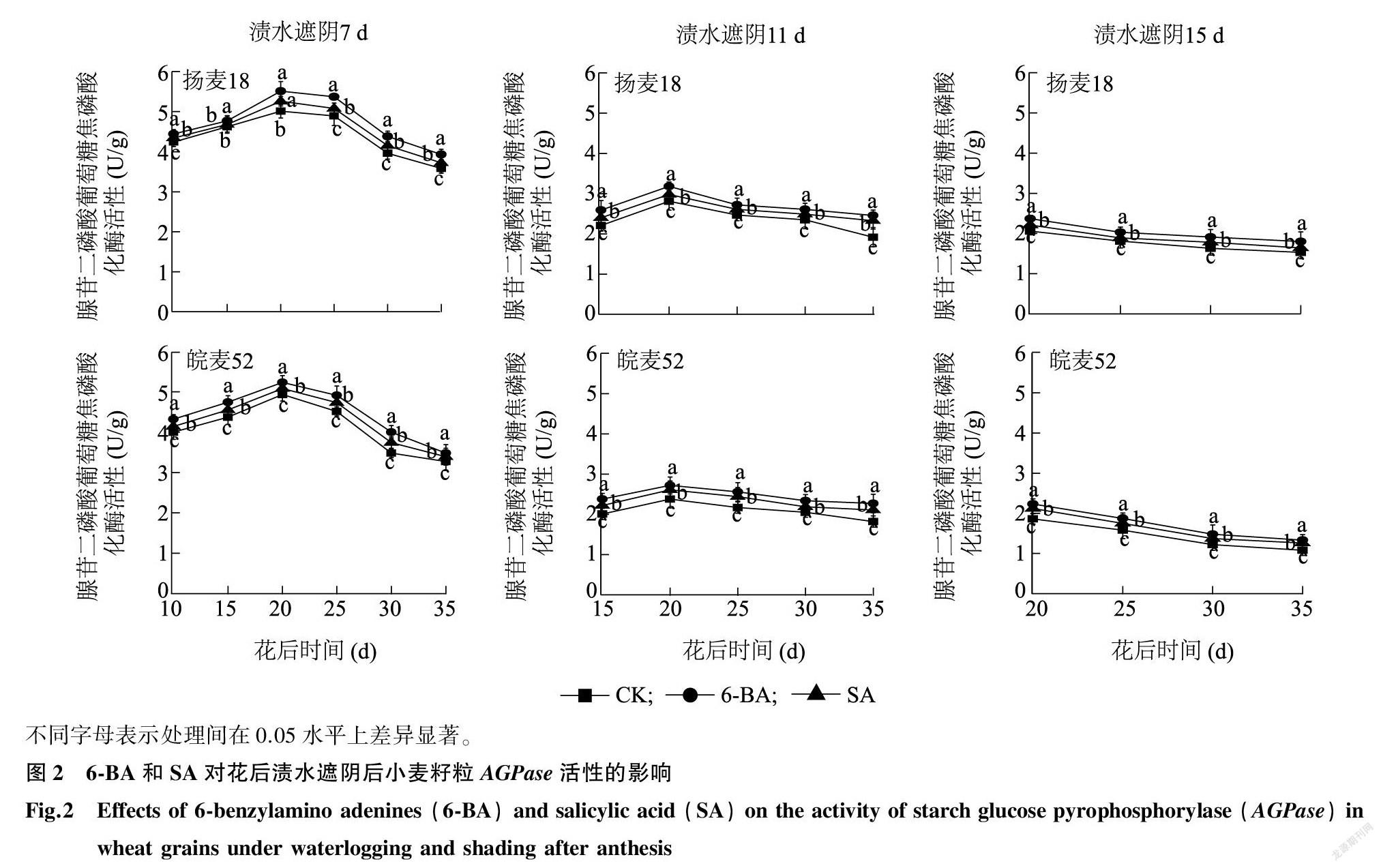
2.1.1 籽粒葡萄糖焦磷酸化酶(AGPase)活性 AGPase主要负责催化淀粉合成的底物腺苷二磷酸葡萄糖(ADPG)的形成[24]。由图2可知,花后渍水遮阴7 d和11 d后,小麦籽粒AGPase活性变化呈单峰曲线,在花后20 d左右达到峰值;渍水遮阴15 d后,籽粒AGPase活性呈下降趋势。渍水遮阴后喷施6-BA与SA提高了小麦籽粒AGPase活性,与喷施蒸馏水的对照间差异达到显著水平(P<0.05,除渍水遮阴7 d后扬麦18花后15 d)。喷施6-BA对AGPase活性损伤的缓解效果优于SA。以皖麦52花后20 d为例,在渍水遮阴7 d、11 d和15 d后,与对照相比,喷施6-BA后籽粒AGPase活性分别提高了6.1%、14.3%和19.9%,喷施SA后籽粒AGPase活性分别提高了3.0%、9.7%和14.0%。
2.1.2 籽粒可溶性淀粉合成酶(SSS)活性 SSS主要负责支链淀粉的合成[25]。从图3可知,花后渍水遮阴7 d、11 d后,小麦籽粒SSS活性变化呈单峰曲线,峰值出现的时间约为花后20 d;渍水遮阴15 d后,籽粒SSS活性呈下降趋势。渍水遮阴后喷施6-BA与SA提升了小麦籽粒SSS活性,与对照间差异达到显著水平(P<0.05)。喷施6-BA对SSS活性损伤的缓解效果优于SA。以皖麦52花后20 d为例,在渍水遮阴7 d、11 d和15 d后,与喷施蒸馏水的对照相比,喷施6-BA的籽粒SSS活性分别提高了30.8%、36.8%和39.8%,喷施SA的籽粒SSS活性分别提高了16.2%、26.9%和29.7%。
2.1.3 籽粒淀粉分支酶(SBE)活性 SBE是参与合成支链淀粉的关键酶,也是将直链淀粉转化为支链淀粉的催化剂[26]。由图4可知,花后渍水遮阴7 d、11 d后,小麦籽粒SBE活性变化呈单峰曲线,峰值出现的时间约为花后20 d。花后渍水遮阴15 d后,籽粒SBE活性呈下降趋势。渍水遮阴后喷施6-BA和SA皆提高了籽粒SBE活性,且喷施6-BA处理籽粒SBE活性与对照间差异达显著水平,喷施SA后扬麦18有部分处理与对照间差异未达到显著水平(P<0.05)。喷施6-BA对SBE活性损伤的缓解效果优于SA。以皖麦52花后20 d为例,渍水遮阴7 d、11 d和15 d后,与喷施蒸馏水的对照相比,喷施6-BA的籽粒SBE活性分别提高了12.7%、15.3%和19.4%,喷施SA的籽粒SBE活性分别提高了6.9%、8.0%、12.0%。
2.1.4 籽粒颗粒结合型淀粉合成酶(GBSS)活性 GBSS主要催化直链淀粉的合成,能够将ADPG上的葡萄糖残基添加到淀粉引物上,加长淀粉的分子链长[27]。从图5可知,花后渍水遮阴7 d、11 d后,小麦籽粒GBSS活性变化呈单峰曲线,且在花后20 d左右达到峰值;渍水遮阴15 d后,籽粒GBSS活性呈下降趋势。渍水遮阴后喷施6-BA和SA均提高了小麦籽粒GBSS活性,与喷施蒸馏水的对照间差异达到显著水平(P<0.05)。喷施6-BA对籽粒GBSS活性损伤的缓解效果优于SA。以皖麦52花后20 d为例,渍水遮阴7 d、11 d和15 d后,与喷施蒸馏水的对照相比,喷施6-BA的籽粒GBSS活性分别提高了7.2%、11.5%和12.4%,喷施SA的籽粒GBSS活性分别提高了4.8%、5.4%和6.2%。
2.2 6-BA和SA对花后渍水遮阴小麦籽粒淀粉积累的影响
由图6可知,在淀粉合成过程中,两品种小麦籽粒淀粉积累量变化呈“S”型曲线,从花后15 d开始淀粉含量迅速增长,花后25 d之后积累量趋于稳定。渍水遮阴后喷施6-BA和SA皆提高了籽粒淀粉的积累量,且各处理间差异达到显著水平(P<0.05)。喷施6-BA对籽粒淀粉累积量损伤的缓解效果优于SA。以皖麦52为例,渍水遮阴7 d、11 d和15 d后,与喷施蒸馏水的对照相比,喷施6-BA的籽粒淀粉的最终积累量分别增长了16.3%、14.4%和10.5%,SA处理使淀粉的最终积累量分别增长了6.7%、9.4%和5.9%。
用Logistic方程对小麦籽粒淀粉积累过程进行拟合(表2),各方程的决定系数均大于0.99。喷施6-BA和SA提高了渍水遮阴后小麦籽粒淀粉最大累积速率(Rmax)和平均累积速率(Rmean)。渍水遮阴7 d后皖麦52喷施6-BA和SA有效延长了淀粉积累持续期(T)和淀粉积累活跃期(D),但渍水遮阴11 d、15 d后,喷施6-BA和SA对淀粉积累持续期和淀粉积累活跃期的影响较小。渍水遮阴后,喷施6-BA和SA提高了淀粉最大积累速率和淀粉平均积累速率,以喷施6-BA的效果较好,并且渍水遮阴胁迫时间越长,喷施6-BA和SA后淀粉最大积累速率和淀粉平均积累速率增加的幅度越大。以皖麦52为例,渍水遮阴7 d、11 d和15 d后,喷施6-BA使淀粉最大积累速率和淀粉平均积累速率分别提高了6.2%、8.0%、8.2%和9.5%、13.3%、17.7%。喷施SA使淀粉最大积累速率和淀粉平均积累速率分别提高了2.1%、5.2%、4.8%和4.1%、8.9%、10.4%。对于扬麦18,渍水遮阴7 d后喷施6-BA、SA缩短了淀粉积累持续期和淀粉积累活跃期,渍水遮阴11 d、15 d后喷施6-BA延长了淀粉积累持续期和淀粉积累活跃期,喷施SA縮短了淀粉积累活跃期,却延长了淀粉积累持续期,但总体而言渍水遮阴处理后喷施6-BA、SA对淀粉积累持续期和淀粉积累活跃期的影响较小。
不同字母表示处理间在0.05水平上差异显著。
2.3 6-BA和SA对花后渍水遮阴小麦籽粒干物质积累的影响
用Logistic方程对小麦籽粒干物质积累过程进行拟合,各方程的决定系数均大于0.990(表3)。喷施6-BA和SA延长了渍水遮阴后两品种小麦籽粒灌浆持续天数(N)和提高了籽粒灌浆平均速率(V)。渍水遮阴后扬麦18喷施6-BA后延长了籽粒灌浆快增期(N2)和灌浆缓增期(N3),对灌浆渐增期(N1)的影响不大;喷施SA主要延长了灌浆渐增期,对灌浆快增期、灌浆缓增期影响较小。渍水遮阴后,皖麦52喷施6-BA和SA后延长了籽粒灌浆渐增期、灌浆快增期、灌浆缓增期(渍水遮阴11 d灌浆渐增期除外),以喷施6-BA的修复效果较好。以皖麦52为例,渍水遮阴7 d、11 d和15 d后,与对照相比,喷施6-BA后籽粒平均灌浆速率分别提高了18.8%、22.7%和22.7%,喷施SA后籽粒平均灌浆速率分别提高了10.3%、9.3%和10.2%。两种植物生长调节剂之间相比较,喷施6-BA对花后渍水遮阴对小麦籽粒干物质积累损伤的缓解效果优于SA。
2.4 6-BA和SA对花后渍水遮阴小麦产量构成因素的影响
从表4可以看出,与对照相比,渍水遮阴后7 d、11 d后喷施6-BA和SA显著提高了两品种小麦的穗粒数、千粒质量。对于扬麦18,与对照相比,渍水遮阴后喷施6-BA和SA穗粒数达到显著性差异,喷施6-BA和SA两处理间无显著差异,但千粒质量在各处理间的差异均达到显著水平。对于皖麦52,穗粒数和千粒质量在各处理间差异都达到显著水平。渍水遮阴15 d后喷施6-BA和SA显著提高了两品种小麦的千粒质量,对于穗粒数的影响未达显著水平。以皖麦52为例,渍水遮阴7 d、11 d后喷施6-BA穗粒数分别提高了22.5%、19.2%,喷施SA穗粒数分别提高了10.8%、13.9%;渍水遮阴7 d、11 d、15 d后喷施6-BA千粒质量分别提高了21.7%、27.1%和32.1%,喷施SA千粒质量分别提高了11.3%、13.6%和17.6%。两种植物生长调节剂之间相比较,喷施6-BA对花后渍水遮阴对小麦产量损伤的缓解效果优于SA。
3 讨论
3.1 6-BA和SA对渍水遮阴后小麦籽粒淀粉合成关键酶活性和淀粉积累的影响
研究者认为,参与小麦籽粒淀粉合成的关键酶主要有AGPase、SSS、GBSS和SBE[28-29]。在籽粒充实过程中,外界环境的变化会影响酶的活性,导致籽粒中淀粉的合成受到影响[30] 。如渍水降低了小麦GBSS、SSS活性[31];弱光胁迫显著减弱了玉米花粒期的AGPase、SSS、GBSS活性,降低了淀粉的最终累积量[32]。前人研究发现,喷施植物生长调节剂可以对淀粉合成关键酶进行调控,增加淀粉累积量,如在水稻灌浆期喷施6-BA和SA显著上调了籽粒GBSS、SSS和SBE相关基因的表达量[33]。本试验研究结果表明,喷施6-BA、SA缓解了渍水遮阴胁迫对小麦籽粒淀粉合成关键酶(AGPase、SSS、GBSS和SBE)活性造成的影响,胁迫时间越长,喷施植物生长调节剂的修复效果越明显。前人研究结果表明,在逆境条件下小麦籽粒中SSS活性受影响最大[34-35]。对水稻喷施CPPU[N-(2-氯-4-吡啶基)-N-苯基脲]和PP333(多效唑)后,籽粒中SSS和SBE活性显著提高,GBSS则具有相对稳定性[36]。本试验研究结果表明,开花期渍水遮阴后喷施6-BA和SA对籽粒中SSS活性的修复效果最为显著,其次是AGPase和SBE,GBSS活性受外源植物生长调节剂的影响较小,说明逆境条件下喷施6-BA和SA更有利于支链淀粉的合成,喷施外源植物生长调节剂对于淀粉组分的影响还需进一步探究。
小麦籽粒的充实过程,主要是淀粉的合成和积累过程。逆境胁迫下籽粒淀粉合成关键酶活性将会受到影响,进而导致籽粒淀粉积累速率下降[37]。前人研究发现,喷施6-BA能有效促进小麦籽粒中胚乳细胞分裂和淀粉的生物合成[38];开花期喷施适宜浓度的SA可以有效提高小麦淀粉产量[39]。本研究发现,渍水遮阴后喷施6-BA和SA提高了小麦淀粉积累速率(淀粉平均积累速率和淀粉最大积累速率),增加了淀粉的最终积累量。在淀粉积累进程中,AGPase活性的变化与淀粉积累的关系极为密切,AGPase活性与淀粉积累呈显著或者极显著正相关[40]。3-磷酸甘油酸能够激活谷物中AGPase活性,有效增加淀粉积累量,PP333可以促进甘薯根块AGPase活性的提升,加快淀粉积累速率,增加淀粉积累量[41-42]。GBSS活性与直链淀粉积累速率呈显著正相关关系[43];SSS活性与支链淀粉积累速率呈显著正相关, 并与总淀粉积累速率呈显著正相关[44]。本研究中,与对照相比,渍水遮阴后喷施6-BA和SA,显著提高了籽粒中淀粉合成关键酶活性、淀粉积累量和淀粉积累速率。喷施6-BA和SA在一定程度上还影响了小麦淀粉积累持续期和活跃期,其中对皖麦52渍水遮阴7 d处理的影响较大,其他处理淀粉积累特性可能更大程度受遗传基因的控制[45]。
3.2 6-BA和SA对渍水遮阴后小麦籽粒干物质积累和产量的影响
小麦籽粒的最终质量主要由灌浆速率和灌浆持续期所决定[46]。前人研究发现喷施植物生长调节剂会影响小麦灌浆进程,如在小麦开花期喷施6-BA和ABA改变了籽粒中的激素水平,促进了籽粒灌浆[47]。喷施6-BA缓解了渍水、遮光胁迫对小麦功能叶片造成的伤害,增加光合作用能力和花后同化物的转运量,进而提高产量[6,14]。本试验研究发现,喷施6-BA和SA促进了花后光合产物的积累与转运,缓解了渍水遮阴胁迫对平均灌浆速率造成的伤害,同时延缓了功能叶片的的衰老进程,延长了籽粒灌浆持续期,且胁迫时间越长,延缓效果越明显。喷施6-BA和SA对于其他灌浆参数(渐增期、快增期和緩增期)的影响由于不同品种对药剂敏感程度存在一定差异而不同。总体而言,喷施6-BA和SA对平均灌浆速率的影响较大。
前人研究发现,开花后渍水遮阴胁迫增加了小麦的不孕小花数,影响了小麦籽粒灌浆,最终减少了小麦的穗粒数和千粒质量[12]。在小麦拔节后25 d喷施6-BA可以显著降低小麦穂部可孕小花的败育速率,提高单穂可孕花结实率,提高小麦穗粒数[48]。在小麦开花期喷施SA能够延缓叶片的衰老,有利于灌浆速率和籽粒质量提高[49]。此外在对水稻的研究中也发现,在水稻开花期喷施SA可以缓解高温胁迫造成的伤害,增加了穗粒数和结实率[50]。本研究中小麦渍水遮阴7 d、11 d后喷施6-BA和SA提高了穗粒数和千粒质量,对产量的恢复效果显著。这可能是因为喷施6-BA和SA减少了小麦穗部小花的退化率从而提高了结实率,同时一定程度上延缓了小麦功能叶片的衰老进程,增强了花后光合作用,提高了花后同化物的转运量及对籽粒的贡献率。渍水遮阴15 d后由于小麦穗粒数已经基本确定,喷施6-BA和SA对小麦穗粒数的影响较小,但是可以有效缓解渍水遮阴胁迫对千粒质量的影响。不同植物生长调节剂之间相比较,喷施6-BA对产量的修复效应优于SA。
4 结论
小麦花后渍水遮阴后喷施6-BA和SA均能缓解渍水遮阴胁迫对小麦籽粒中淀粉合成关键酶活性造成的伤害,缓解渍水遮阴胁迫对淀粉积累速率(淀粉最大积累速率和淀粉平均积累速率)造成的影响,有利于籽粒淀粉合成,还能延长籽粒灌浆期,提高平均灌浆速率,增加籽粒质量。喷施6-BA和SA可缓解渍水遮阴胁迫对穗粒数和千粒质量造成的影响。两种植物生长调节剂间比较,喷施6-BA的修复效应优于SA。
参考文献:
[1] 李 琪,王 清,王连喜,等. 花后遮光渍水对南京冬小麦光合特性和产量的影响[J].江苏农业学报,2017,33(3):555-560.
[2] SINGH S P, SETTER T L. Effect of waterlogging on element concentrations, growth and yield of wheat varieties under farmers sodic field conditions[J].Proceedings of the National Academy of Sciences, India Section B: Biological Sciences,2017,87(2):513-520.
[3] BORU G,GINKEL M V,KRONSTAD W E, et al. Expression and inheritance of tolerance to waterlogging stress in wheat[J].Euphytica,2001,117(2):91-98.
[4] SHAO G C,LAN J J,YU S E, et al. Photosynthesis and growth of winter wheat in response to waterlogging at different growth stages[J].Photosynthetica,2013,51(3):429-437.
[5] 馬尚宇,王艳艳,黄正来,等. 渍水对小麦生长的影响及耐渍栽培技术研究进展[J].麦类作物学报,2019,39(7):835-843.
[6] 李刘龙,卫茗梅,李 秀,等. 外源6-BA对不同时期遮光下江汉平原稻茬小麦产量的影响[J].应用生态学报,2019,30(11):3753-3761.
[7] 顾蕴倩,刘 雪,张 巍, 等. 灌浆期弱光逆境对小麦生长和产量影响的模拟模型[J].中国农业科学,2013,46(5):898-908.
[8] 牟会荣,姜 东,戴廷波,等. 遮光对小麦籽粒淀粉品质和花前贮存非结构碳水化合物转运的影响[J].应用生态学报,2009,20(4):805-810.
[9] 许振柱,于振文,张永丽. 土壤水分对小麦籽粒淀粉合成和积累特性的影响[J].作物学报,2003,29(4):595-600.
[10]姜 东,于振文,李永庚,等. 高产小麦强势和弱势籽粒淀粉合成相关酶活性的变化[J].中国农业科学,2002,35(4):378-383.
[11]王晨阳,张艳菲,卢红芳,等. 花后渍水、高温及其复合胁迫对小麦籽粒淀粉组成与糊化特性的影响[J].中国农业科学,2015,48(4):813-820.
[12]汪 敏,王邵宇,吴佳佳,等. 花后阴雨对小麦籽粒淀粉合成和干物质积累的影响[J].中国生态农业学报,2020,28(1):76-85.
[13]WANG X, LIU D, WEI M, et al. Spraying 6-BA could alleviate the harmful impacts of waterlogging on dry matter accumulation and grain yield of wheat[J].PeerJ,2020,8(7):e8193.
[14]杜厚江,王小燕,赵晓宇. 6-BA对小麦开花期渍害的缓减效应[J].麦类作物学报,2014,34(12):1672-1676.
[15]李建坤,汪宗立. 涝渍逆境下玉米叶片各细胞器超氧物歧化酶活性的变化及6-BA的保护作用[J].江苏农业学报,1992,8(3):20-24.
[16]LARKINDALE J, KNIGHT M R.Protection against heat stress-induced oxidative damage in arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid[J].Plant Physiology,2002,128(2):682-695.
[17]许明丽,孙晓艳,文江祁. 水杨酸对水分胁迫下小麦幼苗叶片膜损伤的保护作用[J].植物生理学报,2000,36(1):35-36.
[18]刘海英,郭天财,朱云集,等. 开花期喷施水杨酸对不同类型专用小麦品种籽粒淀粉及产量的影响[J].麦类作物学报,2006,26(4):123-127.
[19]程方民,钟连进,孙宗修. 灌浆结实期温度对早籼水稻籽粒淀粉合成代谢的影响[J].中国农业科学,2003,36(5):492-501.
[20]SMYTH D A, PRESCOTT H E. Sugar content and activity of sucrose metabolism enzymes in milled rice grain [J].Plant Physiology,1989,89(3):893-896.
[21]NAKAMURA Y, YUKI K, Park S Y, et al. Carbohydrate metabolism in the developing endosperm of rice grains [J].Plant & Cell Physiology,1989,30(6):833-839.
[22]李太贵,沈 波,陈 能,等. Q酶在水稻籽粒垩白形成中作用的研究[J].作物学报,1997,23(3): 84-90.
[23]何照范. 谷物淀粉组分分离及测试方法评述[J].粮食储藏,1985(6):32-38.
[24]YANG J.Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling[J]. Plant Physiology,2004,135(3):1621-1629.
[25]趙步洪,张文杰,常二华,等. 水稻灌浆期籽粒中淀粉合成关键酶的活性变化及其与灌浆速率和蒸煮品质的关系[J].中国农业科学,2004,37(8):1123-1129.
[26]ACHENBACH T V, BRUNNER B, HEERMEIER K.Oligonucleotide-based knockdown technologies: antisense versus RNA interference[J].Chembiochem,2010,4(10):928-935.
[27]RAHMAN S, ABRAHAMS S,ABBOTT D, et al. Erratum: A complex arrangement of genes at a starch branching enzyme I locus in the D-genome donor of wheat[J].Genome,1998,41(1):139.
[28]HSU Y C, TSENG M C, WU Y P, et al. Genetic factors responsible for eating and cooking qualities of rice grains in a recombinant inbred population of an inter-subspecific cross [J].Molecular Breeding,2014,34(2): 655-673.
[29]LUO J, JOBING S A, MILLAR A, et al. Allelic effects on starch structure and properties of six starch biosynthetic genes in a rice recombinant inbred line population[J].Rice,2015,8(1):15.
[30]杨 毅,李 昱,康建宏,等. 花后高温胁迫对春小麦籽粒淀粉合成的影响[J].麦类作物学报,2015,35(11):1535-1541.
[31]范雪梅,姜 东,戴廷波,等. 花后干旱和渍水下氮素供应对小麦籽粒蛋白质和淀粉积聚关键调控酶活性的影响[J].中国农业科学,2005,38(6):1132-1141.
[32]贾士芳,董树亭,王空军,等. 玉米花粒期不同阶段遮光对籽粒品质的影响[J].作物学报,2007,33(12):1960-1967.
[33]金正勋,王思宇,王 珊,等. 外源激素对水稻籽粒碳氮代谢相关酶基因表达影响[J].东北农业大学学报,2020,51(7):1-9,26.
[34]AHMADI A , BAKER D A.The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat [J].Plant Growth Regulation,2001,35:81-91.
[35]HURKMAN W J, MCCUE K F, ALTENBACH S B, et al. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm[J].Plant Science,2003,164(5):873-881.
[36]唐湘如,谭中文,李之林,等. CPPU和PP333对杂交稻3个淀粉合成酶活性和米质的影响[J].杂交水稻,2002,17(3):47-49.
[37]COMMURI P D, JONES R J. High temperatures during endosperm cell division in maize: A genotypic comparison under in vitro and field conditions[J].Crop Science,2001,41(4):1122-1130.
[38]戴忠民.喷施6BA和ABA对冬小麦籽粒胚乳细胞增殖和淀粉积累的影响[J].麥类作物学报,2008,28(3):484-489.
[39]刘海英,郭天财,朱云集,等. 开花期喷施水杨酸对不同类型专用小麦品种籽粒淀粉及产量的影响[J].麦类作物学报,2006,26(4):123-127.
[40]刘 霞,尹燕枰,姜春明,等. 花后不同时期弱光和高温胁迫对小麦旗叶荧光特性及籽粒灌浆进程的影响[J].应用生态学报,2005,16(11):113-117.
[41]TUNCEL A, CAKIR B, HWANG S K, et al. The role of the large subunit in redox regulation of the rice endosperm ADP-glucose pyrophosphorylase[J]. Febs Journal,2015,281(21):4951-4963.
[42]陈晓光,史春余,王振林,等. 多效唑对食用甘薯北京553块根淀粉积累及相关酶活性的影响[J].中国农业科学,2012,45(1):192-198.
[43]KEELING P L, WOOD J R, TYSON R H, et al. Starch biosynthesis in developing wheat grain: evidence against the direct involvement of triose phosphates in the metabolic pathway[J].Plant Physiology,1988,87(2):311-319.
[44]NAKAMURA Y.Towards a better understanding of the metabolic system for amylopectin biosynthesis in plants: rice endosperm as a model tissue[J].Plant & Cell Physiology,2002,43(7):718-725.
[45]PANOZZO J F, EAGLES H A.Cultivar and environmental effects on quality characters in wheat I Starch[J].Australian Journal of Agricultural Research,1998,49(5):629-636.
[46]李文阳,尹燕枰,闫素辉,等. 不同粒型小麦品种籽粒内源激素变化与籽粒灌浆特征的比较[J].华北农学报,2007,22(1):5-8.
[47]杨东清,李玉玲,倪英丽,等. 外源ABA和6-BA对不同持绿型小麦籽粒灌浆进程及蛋白质含量影响[J].作物学报,2014,40(2):301-312.
[48]郑春风,任 伟,朱云集,等. 冬小麦小花发育及结实特性对叶面喷6-BA的响应[J].植物营养与肥料学报,2017,23(3):774-780.
[49]RASKIN I. Salicylate, a new plant hormone[J]. Plant Physiology,1992,99(3):799-803.
[50]杨 军,蔡 哲,刘 丹,等. 高温下喷施水杨酸和磷酸二氢钾对中稻生理特征和产量的影响[J].应用生态学报,2019,30(12):4202-4210.
(责任编辑:张震林)
