面向植物抗旱性研究的多源表型信息采集和分析技术
2022-03-14张慧春杨琨琪李杨先边黎明郝明灼
张慧春 杨琨琪 李杨先 边黎明 郝明灼
(1.南京林业大学机械电子工程学院, 南京 210037; 2.南京林业大学林学院, 南京 210037; 3.南方现代林业协同创新中心, 南京 210037)
0 引言
干旱是全球最普遍和最严重的自然灾害,全球每年因干旱造成经济损失达80亿美元,远超其他气象灾害[1]。近些年,全球气温不断上升,各地区水资源逐渐匮乏,严重影响植物的生长和产量。干旱环境中,植物自身进化出一系列自我保护和适应与抵制不利环境胁迫的机制,其表型特征会发生显著改变,来尽可能降低逆境环境对其生长发育的影响[2-5],例如,植物可通过降低株高来缩短地下水分到达地上部分的距离,有利于水分的上下传输等[6]。表型是指受基因和环境决定或影响的复杂植物性状,包括生长、发育、耐性、抗性、生理、结构、产量等。可测量的表型参数分为形态结构表型参数和生理生化表型参数,其中形态结构参数包括植物高度、叶形态、根形态、生物量、果实特征;生理表型参数包括叶片水分利用率、叶绿素、细胞膜透性、光合速率等[7-11]。因此,研究植物在干旱胁迫下的表型变化,解析植物对不同环境变化的适应机制,将会有助于在应对全球气候变化时培育耐旱品种,对保障农林可持续发展具有重要意义。
植物生长过程中受多种因素共同作用,因此干旱等逆境造成的植物表型性状体现出复杂性。尽可能多的获取植物的表型参数有助于实时了解植物在干旱胁迫下的长势,从而对植物的生长发育进行指导管理。随着图像处理技术的迅速发展,表型图像采集的方法种类也愈来愈多,成像传感器的应用越来越广泛。每种成像方式都从植物获取某些信息,不同的成像方法可以提供同一植物不同方面的信息[12]。植物具有特殊的变异性、随机性和复杂性,它的生长发育、生理生化变化都受到各种因素影响,完整而丰富的植物表型信息无法依赖某一个传感器来获取,单独从某一种图像中也无法得到全面的表型信息。在植物表型研究中通过多个传感器的协同来获取多源表型信息,并采用一定的数据关联性分析,可以得到植物的形态表型参数及生理参数。因此,构建表型信息采集平台,使用多个传感器采集同一目标植物的信息,经过适当处理,将采集的多源表型信息关联分析、综合互补,是获取准确客观的植物表型信息的有效方法。
簸箕柳(SalixsuchowensisCheng)是杨柳科柳属植物,基因组大小为356 Mb,具有易繁殖、速生丰产和轮伐期短等特点,是重要的生物质能源和造林用材树种[13]。本文以模式树种杨树的姐妹属植物簸箕柳苗木为研究对象,构建一套多源信息采集系统,对获取到的表型信息进行关联分析,实时、连续监测簸箕柳苗木的生长状况,指导抗性育种中水分的施用和抗旱基因型的快速筛选,以期为簸箕柳苗木的抗旱性表型指标筛选提供研究平台和技术支撑[14-15]。
1 材料与方法
1.1 供试材料
供试材料选取直径(4±0.2) mm、高度(12±0.3) cm的40段簸箕柳枝条进行扦插式种植,以南京林业大学林区有机土壤为基质,放入花盆(直径10 cm,高11.5 cm)中,并置于人工气候箱(RXZ型,宁波江南制造厂)中培养,人工气候箱的环境参数设置如下:相对湿度65%、温度28℃、光照强度278 μmol/m2、昼夜时间比16∶8。扦插种植第12天,将长势相同的簸箕柳分为2组,每组20株。其中一组20株进行正常施水处理,另一组20株进行干旱胁迫处理。正常施水处理组,通过水分控制模块与称量模块,保持每盆总质量为950 g。干旱胁迫处理试验分2个阶段进行,第1阶段是从第26天开始到第42天结束,每4 d随机选取2株进行破坏性取样,第26天第1次取样测量,16 d共记录5次,共计10株,每次将取样得到的植株立即称量以获得植株鲜质量(Fresh weight,FW),之后将该植株置于70℃的电热恒温鼓风干燥箱(DHG-9101-2SA型)中保存24 h,称量获取植株干质量(Dry weight,DW)。干旱胁迫试验第2阶段是从扦插后第42天开始到第62天结束,剩余未进行破坏性取样的10株植物继续在培养箱中生长并每2 d进行成像。这一阶段不进行破坏性取样,直至试验结束。
1.2 多源表型信息采集平台设计与图像获取
本文构建的多源表型信息采集平台由图像采集模块(GO-5000C-PGE型,分辨率2 560像素×2 048像素,LM8HC-SW定焦镜头)、热成像仪(FLIR T460型,热灵敏度小于0.03℃@+30℃)、自动称量模块、水分精确控制模块、光源及平台框架组成,如图1所示。其中平台框架的尺寸为1 m×1 m×1.5 m,目标植物位于平台中心处。图像采集模块中2个垂直摆放的CCD 相机从不同角度采集植物形态结构表型信息。热成像仪用于获取植物的冠层热成像图像。自动称量、水分精确控制模块主要包括压力传感器、A/D转换芯片、继电器、电磁阀、水泵等器件。自动称量模块获取植株的质量。水分精确控制模块根据获取的植株质量信息与预先设定的质量之间的差值来精确施加定量的水分。光源用于确保拍摄环境的稳定可靠,尽量减少噪点对后期图像处理的影响。
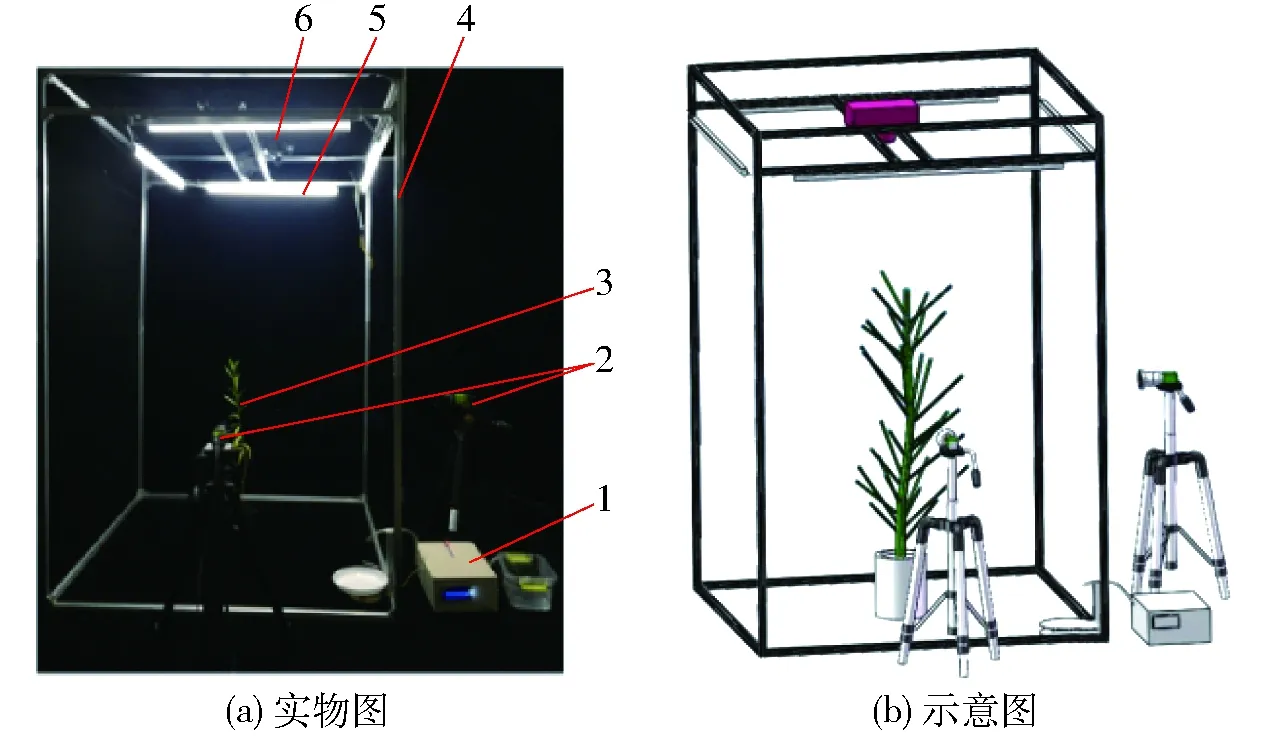
图1 多源表型信息采集平台Fig.1 Multi-source phenotypic information collection platforms1.水分精确控制模块和自动称量模块 2.图像采集模块 3.目标植物 4.平台框架 5.光源 6.热成像仪
1.3 图像处理算法
1.3.1彩色图像处理算法
为了便于提取彩色图像中植株信息,需要将彩色图像中植株部分分割出来。目前较为成熟的目标分割多采用图像处理技术,该方法可以通过阈值分割算法实现前景和背景的分离,实现目标识别,图2为利用图像处理技术检测簸箕柳的流程图。但是该方法受光照不均、反光或背景复杂等因素影响,存在最优阈值求解困难、适应性差、分割效率低下等缺点。

图2 图像处理算法分割簸箕柳效果Fig.2 Effect drawings of image processing algorithm segmentation of Salix suchowensis Cheng
针对传统图像处理技术存在的缺点,本文提出了一种新的自动图像分割算法,在对目标图像进行自动分割之前,基于目标检测算法YOLO v3(You only look once v3)对目标区域进行定位与识别[16-20],定位与识别到图像中植物与红色标定板区域并设置为感兴趣区域(Region of interest,ROI),再基于颜色信息分割出植物和红色标定板,图3为利用YOLO v3检测簸箕柳的方法示意图。
考虑到簸箕柳叶片和茎的生长具有随机性,且深度学习算法需要大量的数据,本文使用CCD相机采集簸箕柳图像共3 600幅,其中90%用于训练,10%用于测试。所有的训练和测试均在处理器为AMD Ryzen 7 2700,内存16 GB,GPU为NVIDIA 1660Ti的台式计算机上完成,并使用Tensorflow 1.6.0的深度学习框架进行训练。使用基于动量的随机梯度下降法来训练数据,动量设置为0.9,初始学习率设为0.001,共训练了200个周期(epoch)。模型在前15个epoch中迅速拟合,损失值快速减少,到达第100个epoch后逐渐稳定。为了避免模型出现过拟合,本文选用第100个epoch的模型权值进行试验。

图3 利用YOLO v3检测簸箕柳图像方法示意图Fig.3 Image method schematic of Salix suchowensis Cheng detected by YOLO v3
利用输出的目标检测模型进行试验,可以检测到簸箕柳和红色标定板区域。设置图中簸箕柳和红色标定板为感兴趣区域,其中的红色标定板为了提取表型参数而设立[21]。为了便于提取图像中的信息,通过2G/(R+B)和2R/(R+B)这2种色彩参数将2个感兴趣区域转换为单波段图像[22],并通过超绿色灰度算法对簸箕柳区域进行灰度化处理。经过超绿色灰度化处理后的图像包含噪点,为获取去噪后簸箕柳的精确图像,本文采用了形态学操作中的开运算对图像进行降噪处理。之后利用边缘检测算法对图像中的簸箕柳边缘提取、标注和连通,从而获取到簸箕柳的分割图像。CCD相机采集的簸箕柳图像分割过程及效果如图4所示。
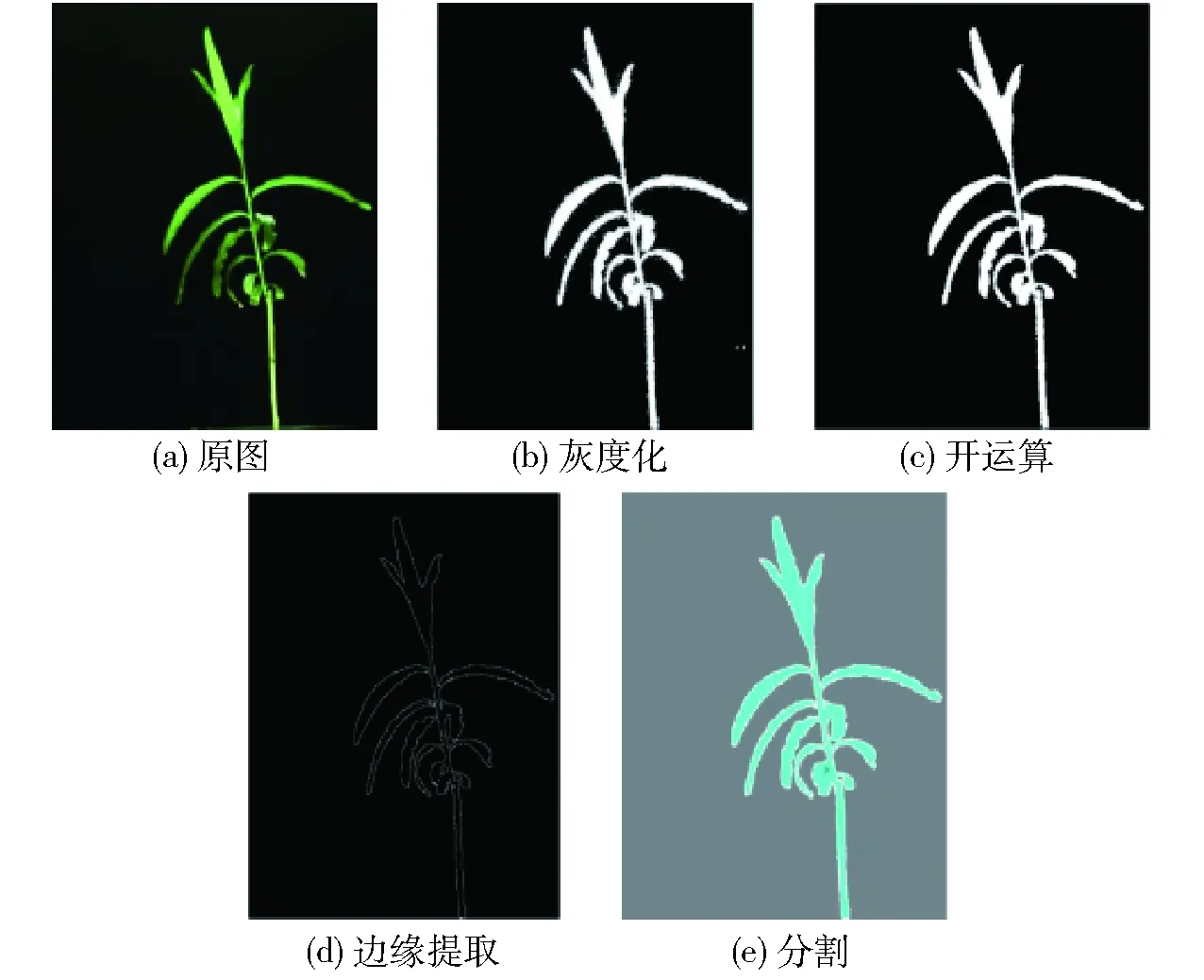
图4 CCD相机采集的簸箕柳图像分割过程示意图Fig.4 Image segmentation processing diagrams of Salix suchowensis Cheng by CCD camera
对比使用传统图像处理算法与本文提出的自动图像分割算法对采集到的图像中簸箕柳进行分割的效果,如图5所示。由图5可知,基于传统图像处理算法会因为叶片尺寸不同、花盆材料反光、背景复杂等原因,存在阈值设定不合理造成簸箕柳图像出现叶片局部断裂、边界轮廓不连续、整体模糊、分割不完全等现象,影响了后期植物表型参数的精准提取。而本文先基于目标检测算法YOLO v3对目标区域进行定位与识别,再通过图像处理算法进行图像处理,避免了传统图像处理算法分割时出现的问题,由此可见本文算法适应性更好,精度更高,提升了识别算法的鲁棒性,提高了表型数据的获取效率,保证了数据的客观准确性。
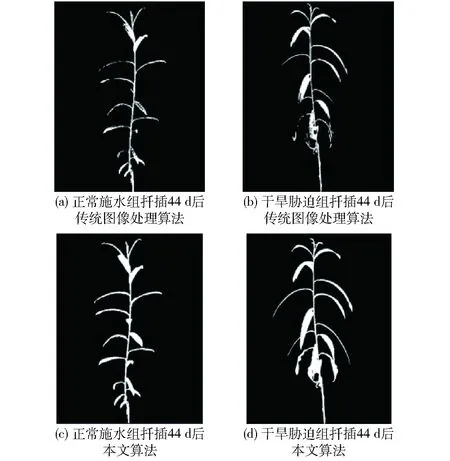
图5 两种算法的簸箕柳图像分割效果对比Fig.5 Comparison of two algorithms for image segmentation effect of Salix suchowensis Cheng
1.3.2红外热图像处理算法
采用红外热成像检测技术来检测簸箕柳不断散发出的热辐射能,用于研究簸箕柳体内水分与冠层温度之间的关系。采集簸箕柳红外热图像过程中,由于图像对比度较小以及噪声的存在,使得图像中很多细节比较模糊。采用直方图均衡算法进行图像增强,经过直方图均衡化后的簸箕柳热红外图像去除了周围的环境温度,突出了簸箕柳的红外热成像信息。但是周围仍然存在比较明显的噪声,为获取准确的冠层信息,需要去除周围的噪声,采用中值滤波算法去除直方图均衡化后的红外热图像噪声,并通过Matlab中人工交互式选区的方法去除花盆区域,最后将图像转换为二值化图像,图6为红外热成像仪采集的簸箕柳图像处理效果。在FLIR Tools软件中将整幅红外热图像的温度信息导出,每个像素点都对应着其所在位置的温度信息。通过提取二值化图像中白色区域(即像素值为1)的像素位置,然后将这些像素位置所对应的温度信息提取出来并计算平均值,便可以将冠层温度提取出来。
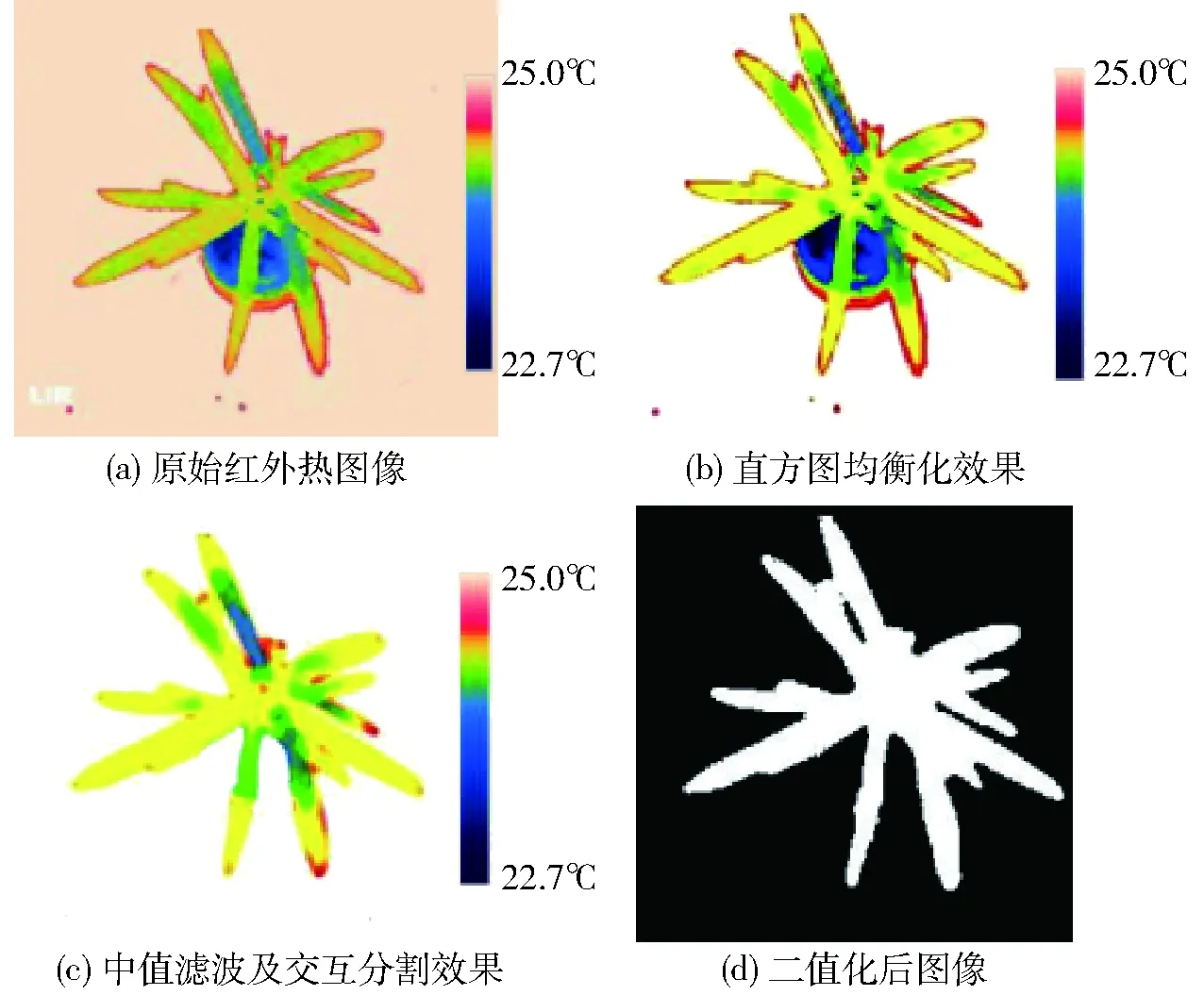
图6 红外热成像仪采集的簸箕柳图像处理效果Fig.6 Image processing flows of Salix suchowensis Cheng by infrared thermal image
2 植物表型参数提取
2.1 投影叶面积与株高
经过图像分割得到簸箕柳图像后,通过像素计数法计算侧视投影叶面积(Projected area,PA)所占像素区域面积。绘制簸箕柳图像的最小包围矩形,利用已知尺寸(2 cm×2 cm)的红色正方形标定板进行比例缩放,从而得到植株投影叶面积与株高,为了减小误差,结果取2个角度所得的平均值。簸箕柳的侧视投影叶面积计算方法为
(1)
式中SPA——实际簸箕柳的侧视投影叶面积,cm2
Slabel——标定板实际面积,取4 cm2
Nlabel——二值图像中标定板投影区域像素数
Nplant——二值图像中簸箕柳投影区域像素数
簸箕柳的株高计算方法为
(2)
式中Hplant——实际簸箕柳的株高,cm
Clabel——标定板的实际周长,8 cm
2.2 叶片数量
本文对原始的霍夫直线变换算法进行改进,首先在簸箕柳的二值化图像中使用直线检测算法识别到簸箕柳的茎,之后根据识别到的2条直线设置左、右距离从而限定出茎的范围,将线段内的所有像素点去除,便可以将茎去除。并基于二值图像寻找簸箕柳叶片轮廓,并将每一个轮廓用彩色显示出来。最后通过骨架提取算法将簸箕柳骨架提取出来。在骨架提取的基础上,对叶片数量(Leaf number,LN)进行提取,图7e中红色圈内为有交点的叶片,如果叶片发生了交叉,则交叉的叶片为一个区域。如果叶片没有交叉,则单独的叶片为一个区域。叶片数量的计算方法为:所有的叶片区域数目加交点个数。普通非交点处像素周围最多有2个值为1的像素点,而交点周围的像素值为1的点至少有3个(如图7e中细节放大图)。因此检测交点的方法为:遍历骨架提取二值化图像的所有像素点,如果像素值为1,则此处为白色,然后遍历该像素点周围的8个像素点。如果有3个像素的值为1,则此处为交点。此算法可能会在交点处检测到2个点,保留1个即可。该叶片数量提取算法面对一般情况的叶片交叉具有较好的鲁棒性,基本满足叶片数目提取需求。

图7 簸箕柳去茎处理及骨架提取效果图Fig.7 Flow charts of stem removal and skeleton extraction of Salix suchowensis Cheng
2.3 耗水量和水分利用效率
耗水量可以定义为蒸腾、土壤水分蒸发蒸腾损失总量或系统总水量的输入[23],是研究植物体内水分的一个重要表型参数。水分利用效率是灌溉和耐旱研究的重要指标,通常定义为生物量积累与耗水量的比值[24]。本试验中水分利用效率定义为每单位水分损失所产生的地上部分干物质量[25]。
平台的称量模块、水分精确控制模块和估测的簸箕柳植株鲜质量和植株干质量,可以用来估算每株簸箕柳的日耗水量(即土壤水分蒸发蒸腾损失总量(Evapotranspiration,ET))和水分利用效率(Water use efficiency,WUE)。
ET=[mi,after-mFW,i-(mi+1,before-mFW,i+1)]/T
(3)
WUE=(mDW,i+1-mDW,i)/(TET)
(4)
式中ET——土壤水分蒸发蒸腾损失总量
WUE——水分利用效率
mi,after——第i次施水后总盆质量,g
mi+1,before——第i+1次施水前总盆质量,g
mFW,i——第i次称取的植株鲜质量,g
mFW,i+1——第i+1次称取的植株鲜质量,g
mDW,i——第i次称取的植株干质量,g
mDW,i+1——第i+1次称取的植株干质量,g
T——时间间隔,取2 d
3 干旱胁迫试验下植物表型参数分析
3.1 不同水分处理下植株的表型参数对比
图8为不同水分处理下簸箕柳的生长图像,正常水分和干旱胁迫下的植物都是从第12天开始进行形态表型参数的采集,采集周期为8 d。由于从扦插后的第52天开始,干旱胁迫植株出现明显枯萎状态,因此干旱胁迫植株从扦插后的第52天开始停止形态表型参数的采集。干旱胁迫植株一共进行了6次(分别是扦插后的第12、20、28、36、44、52天)形态表型参数的采集,正常施水植株一共进行了7次(分别是扦插后的第12、20、28、36、44、52、60天)形态表型参数的采集。因此正常施水植株进行了7次三维形态模型的重建,正常施水植株相应时期拍摄的实际真实图像与模型重建图如图8a所示,干旱胁迫植株进行了6次三维形态模型的重建,干旱胁迫植株相应时期拍摄的实际真实图像与模型重建图如图8b所示。由图8可见,株高、植物鲜质量、叶片数量等表型参数在干旱胁迫条件下比正常施水情况下明显小。

图8 2种不同水分处理条件下真实图像与模拟图像对比Fig.8 Comparisons of real and simulated images under two different water treatment conditions
对正常施水与干旱胁迫2种不同处理下的簸箕柳表型参数进行描述统计,数据如表1所示。由表1可知,正常施水植株的株高显著大于干旱胁迫植株的株高,且正常施水植株的投影叶面积也显著高于干旱胁迫植株,而2种处理下的簸箕柳叶片数量没有显著差别,说明正常施水植株的叶片增长速率高于干旱胁迫植株的叶片增长速率。正常施水植株日耗水量显著高于干旱胁迫植株,原因可能是正常施水植物生长较快增加了对水的需求,相应的蒸腾作用也会增强,同时干旱胁迫植株启动了对干旱胁迫的节水响应(如气孔关闭),从而降低水分的消耗。由于花盆表面的水分蒸发损失较大,2种水分胁迫下的植物日耗水量较多,因此水分利用效率均较低。
利用多源表型信息采集平台获取不同水分条件下的植株表型参数,绘制出各表型参数描述统计表(表1),更加直观地表现出干旱胁迫对植株形态及生理表型参数的影响,也为植株干旱评估提供了帮助与参考。
3.2 植株投影叶面积与地上部分鲜质量、干质量关联分析
采用试验阶段破坏性获取的方式得到簸箕柳鲜质量和干质量参数,并与植株投影叶面积进行关联分析。植株投影叶面积与植株鲜质量之间的相关性如图9a所示,植株投影叶面积与植株鲜质量之间呈现很高的线性正相关性,决定系数R2为0.89;植株投影叶面积与植株干质量之间的相关性图如图9b所示,植株投影叶面积与植株干质量之间呈现线性正相关性,决定系数R2为0.78;植株干质量与植株鲜质量之间的相关性如图9c所示,植株干质量与植株鲜质量之间呈现线性正相关性,决定系数R2为0.76。由此可见,通过多源表型信息采集系统采集到簸箕柳投影叶面积,可以估算出簸箕柳的鲜质量与干质量,从而计算出簸箕柳植株的含水率,以便评估簸箕柳干旱情况。
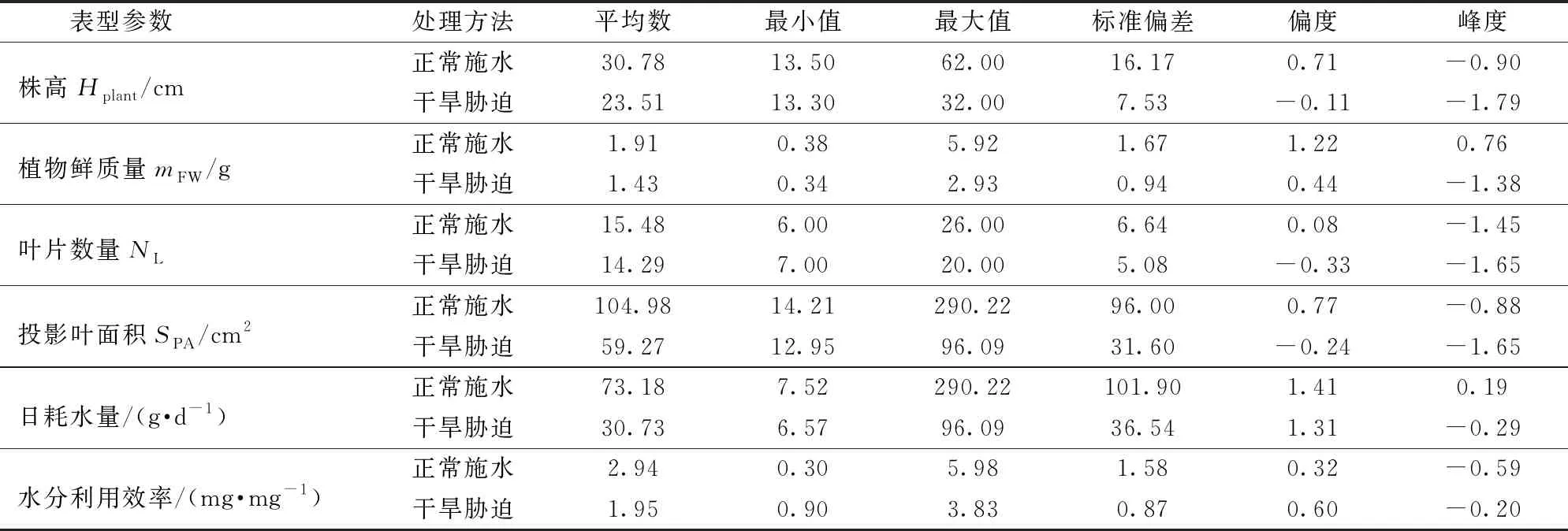
表1 不同处理方法下的簸箕柳表型参数描述统计Tab.1 Statistics of Salix suchowensis Cheng phenotypic parameters under different treatment conditions

图9 植株投影叶面积、植株鲜质量、植株干质量相关性分析Fig.9 Analysis diagrams of relationship between plant projected area and plant fresh weight and dry weight
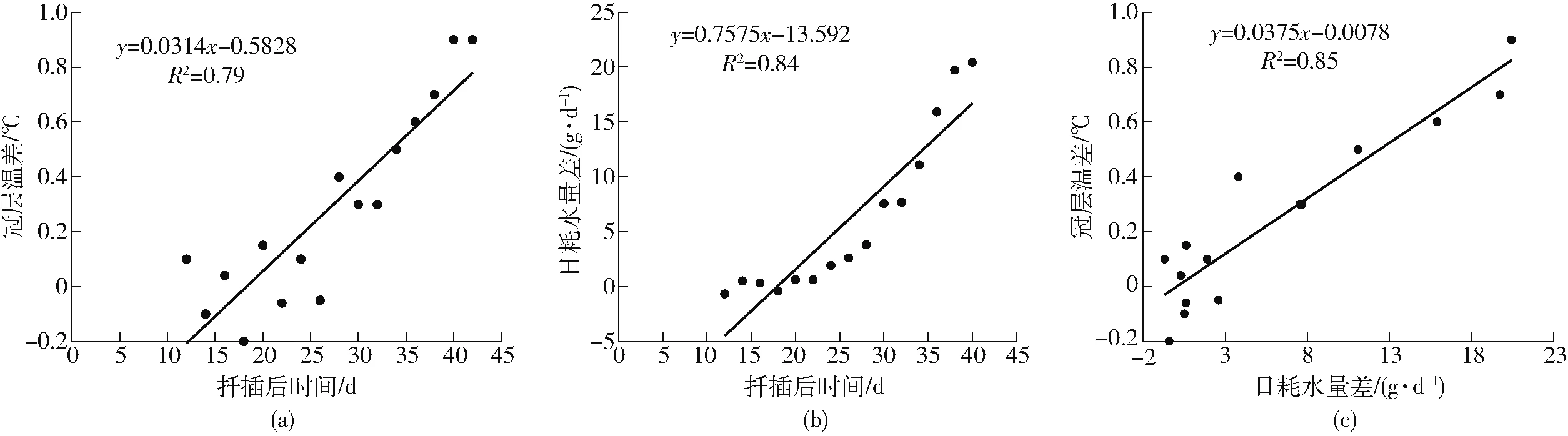
图10 冠层温差、时间、日耗水量差之间相关性分析Fig.10 Relationships between canopy temperature difference and time and daily water consumption difference
3.3 冠层温度与日耗水量关联分析
植物冠层温度反映冠层的能量平衡状况,与植物光合生理特性及产量形成密切相关,是一个很好的干旱监测指标,已有研究[26]表明植物蒸散量与冠层温度之间密切相关,基于能量平衡原理,可利用冠层温度估算植物蒸散量,并进行精量灌溉决策。植物的冠层温度受环境温度影响较大,在采集冠层温度时,无法保证环境温度完全相同,因此此处研究不同水分处理条件下植物的冠层温差随时间的动态变化。本文对不同水分处理条件下植物的平均冠层温差(干旱胁迫植株冠层温度减去正常施水植株冠层温度)与平均日耗水量差(干旱胁迫植株日耗水量减去正常施水植株日耗水量)和时间进行了关联分析,相关性如图10所示。由图10a可知,冠层温差与扦插后时间呈现正相关,决定系数R2为0.79;扦插初期,2种处理下的植物冠层温差并未明显变化,但是随着时间推移,冠层温差逐渐变大,最大温差达到0.9℃。冠层温度受土壤水分环境调控,随着干旱胁迫程度的加重,土壤中水分供应不足,植株蒸腾作用减弱,蒸腾耗热减少、感热通量增加,从而使得植株冠层温度上升;由图10b可知,日耗水量差与扦插后时间呈现出一定的正相关,决定系数R2为0.84。扦插初期,由于土壤水分含量较高,可以保证植物正常供水,叶片耗水量都处于正常范围。随着干旱胁迫的加剧,2种处理下的植物日耗水量差明显变大,最大达到20.4 g/d。原因可能是随着时间推移,干旱胁迫程度的加重,呼吸作用加强,干旱胁迫迫使植物产生应激反应(如气孔关闭),导致植物的蒸腾作用减弱,叶片温度上升,从而使得日耗水量差值变大。由图10c可知,日耗水量差与冠层温差呈现出一定的正相关,决定系数R2为0.85。可见,通过使用表型信息采集系统中的热成像模块获取2种不同水分条件下的冠层温差,可以对植株的受干旱胁迫程度进行监测与评估,也为评判植物中的水分输送利用提供了技术依据。
4 结论
(1)设计并构建了面向簸箕柳表型分析的多源表型信息采集平台。该平台主要由图像采集模块、热成像模块、自动称量模块、水分控制模块组成。实现了植物图像采集、冠层热成像图像获取、水分精确控制等功能。
(2)基于YOLO v3目标检测算法检测并分割出图像中的感兴趣区域,并利用图像处理技术提取了植物的形态结构表型参数(株高、投影叶面积、叶片数量)。其中叶片数量是通过将原有的霍夫直线检测算法进行改进,从而准确提取出簸箕柳的茎,使得簸箕柳的茎叶分离,进一步通过骨架提取算法提取得到叶片数量。通过对热成像仪得到的红外图像处理,获得更准确的冠层温度。
(3)设计了干旱胁迫试验,对不同处理下的簸箕柳表型信息参数进行了对比以及相关性分析。结果显示干旱胁迫下的簸箕柳株高、投影叶面积、日耗水量、水分利用效率都随时间推移显著低于正常施水下的簸箕柳,而冠层温度却高于正常施水下的簸箕柳。通过相关性分析得出:簸箕柳投影叶面积与鲜质量及干质量都呈现线性正相关,决定系数分别为0.89和0.78;植株鲜质量与植株干质量之间呈现线性正相关,决定系数为0.76;扦插后时间与平均冠层温差之间呈正相关,决定系数为0.79;日耗水量差与扦插后时间呈现出一定的正相关,决定系数为0.84;冠层温差与日耗水量差呈正相关,决定系数为0.85。
(4)面向植物抗旱性研究的多源表型信息采集平台能够扩大传感范围,使获取的目标植物表型信息更加丰富完善,提高系统可靠性的同时增强了表型信息的利用效率。在植物未遭受严重的干旱胁迫时,通过采集得到植株投影叶面积、热成像信息等表型参数,可以对植物的鲜质量、干质量、水分利用效率进行估算,实现对植株受干旱胁迫程度的评估。
