枣园生草处理对土壤养分和细菌群落的影响
2022-03-11王中堂沙建川解小锋孟晓烨赵登超
王中堂 沙建川 解小锋 孟晓烨 赵登超 彭 玲 张 琼,*
(1 山东省果树研究所,山东 泰安 271000;2 山东省林业厅经济林管理站,山东 济南 250014;3 山东省林业科学研究院,山东 济南 250014;4 滨州学院山东省黄河三角洲生态环境重点实验室,山东 滨州 256600)
枣(ZizyphusjujubaMill.)是原产我国的重要果树,果实营养价值极高,素有“活维生素丸”之美誉[1]。该树种具有耐瘠薄与抗盐碱的特性,是我国北部盐碱地和瘠薄山区的主要经济林树种之一,为当地产生了较好的经济效益、生态效益和社会效益。针对我国大部分枣园立地条件差,水肥流失严重的现状,目前果园生草是一种优良的土壤管理制度[2]。果园生草是指在果树行间(株间)或全园种植多年生禾本科或豆科草作为土壤覆盖的管理措施,具有改良土壤质量、减少水分流失、促进果树生长发育及改善果实品质、调节果园微域生态环境等优点,现已成为世界上许多国家和地区广泛采用的果园土壤管理模式[3-8]。近年来研究表明,与清耕相比,果园生草可优化土壤微生物群落,加快土壤有机质分解,其中土壤细菌在矿质养分转化和养分吸收中扮演着重要角色[9-11]。
我国果农主要采用节本增效的措施实现持续增收,因此明确适合我国国情的草种及其省力高效的生草模式,对果园生草技术的推广应用具有重要意义。陈学森等[12]在苹果园研究发现一年生豆科植物长柔毛野豌豆(ViciavillosaRoth.)与其他草种相比具有5个特点:一是具有固氮作用;二是适应能力强,在我国各地均有分布;三是具有冬小麦生长特性,即秋季播种后经一个冬季的冷冻处理,翌年春季可爬蔓生长,易盖满地面,不仅减少其他杂草生长,还可减少水分蒸发,具有保水保肥的作用;四是种子无休眠,浸水24 h后即可萌发生长;五是根系浅,茎木质化程度极低,且6月结豆荚后,植株很快腐烂,无需刈割。目前国内文献报道中长柔毛野豌豆仅在苹果园和梨园进行过种植效益评价[12-13],在枣园中的相关研究尚鲜见。基于此,本研究通过田间试验,探究了枣园种植长柔毛野豌豆对土壤综合肥力、土壤细菌群落丰度和结构多样性的影响,旨在为长柔毛野豌豆在枣园生草中的推广应用提供科学依据。
1 材料与方法
1.1 试验材料与试验地概况
试验于2019年10月—2020年9月在山东省农业科学院果树研究所泰东试验基地进行,试验地位于山东省泰安市(35°~36°N,116°~117°E),属北温带大陆性半湿润季风气候区,雨热同季,主要集中在6—9月,年降水量约697 mm,无霜期195 d。土壤主要为黏质壤土,含有机质9.18 g·kg-1,硝态氮32.56 mg·kg-1,铵态氮20.42 mg·kg-1,速效磷15.12 mg·kg-1,速效钾124.32 mg·kg-1,pH值7.7。
供试枣树为自主选育的金丝4号新品种,10月上旬完熟,树体5年生,株行距1.5 m×4 m,长势良好。长柔毛野豌豆种子购自山东天地园艺科技有限公司。
1.2 试验处理
试验共设置3个处理:(1)果园清耕,试验期间在清耕区及时清除杂草;(2)自然生草,当地自然生草群落主要为马唐[Digitariasanguinalis(L.)Scop]、丝茅(茅草)[Imperatakoenigii(Retz.)Beauv.]、葎草(拉拉秧)[Humulusscandens(Lour.)Merr.]、中华小苦荬(苦菜)[Ixeridiumchinense(Thunb.)]、荠菜[Capsellabursa-pastoris(L.)Medic.]及稗[Echinochloacrusgalli(L.)Beauv. var.crusgalli],试验期间刈割3次,保持自然杂草高度15~20 cm,维持自然生草覆盖;(3)种植长柔毛野豌豆,于2019年10月在枣树行间(距主干30 cm以上)开沟播种长柔毛野豌豆种子,播种密度为22.5 kg·hm-2,长柔毛野豌豆开花时间为每年4—6月,7月后倒伏枯萎,落地种子9月初长出第二茬,一次播种可连续多年生长,试验区域土壤为长柔毛野豌豆与少量自然杂草覆盖。每个处理的面积均为667 m2,栽植3行枣树,共110株,单行为1个重复。其他施肥、浇水、修剪以及整枝等管理措施均保持一致。
于2020年9月10日分别对各处理的行间和根际取土,行间取土利用土钻取0~30 cm土层,取土位置为两行枣树的中间点。根际取土为小心挖出枣树根系,收集附着在细根(≤2 mm)上的土,并去除较大颗粒土,即为根际土。每处理分别取3次重复,用于各指标测定。
1.3 测定项目与方法
1.3.1 土壤理化性质及酶活性测定 采用烘干法测定土壤含水量;采用残渣烘干法测定土壤全盐量;利用PHS-3C型pH计(雷磁,上海)测定土壤pH值;采用重铬酸钾外加热法测定土壤有机质含量(soil organic matter,SOM);采用凯氏定氮法测定土壤全氮含量(total nitrogen,TN);利用AA3流动注射分析仪(天津中通科技发展有限公司)测定土壤硝态氮含量(NO3-N);采用碳酸氢钠-钼锑抗比色法测定土壤速效磷含量(Olsen-P);采用火焰光度法测定土壤速效钾含量(available potassium,AK);采用环刀法测定土壤容重(bulk density,BD)[14]。土壤脲酶活性采用苯酚钠比色法测定,单位以NH3-N mg·g-1(37℃,24 h)表示;土壤蔗糖酶活性采用3,5-二硝基水杨酸比色法测定,单位以mg·g-1(37℃,24h)表示;土壤碱性磷酸酶活性测定采用磷酸苯二钠比色法,单位以mg·g-1·d-1(37℃,24 h)表示;土壤碱性过氧化氢酶活性采用高锰酸钾滴定容量法测定,单位以0.02 mol·L-1KMnO4mL·g-1(25℃,20 min)表示,蛋白酶活性测定采用铜盐比色法,单位以NH3-N mg·g-1(37℃,24 h)表示[15]。
1.3.2 果实品质测定 于果实完熟期(10月5日)在树体四周均匀选取果实,1株树上选取5个果实,每个处理共选取30个果实,果实放入干冰盒中,及时带回实验室。可溶性糖含量用蒽酮比色法测定,可滴定酸含量用NaOH 滴定法测定,维生素C(vitamin C,Vc)含量用钼蓝比色法,类黄酮含量用紫外光分光光度法,硬度用TA.XT plus型质构仪(英国Stable MicroSystems公司)测定。
1.3.3 土壤细菌的16S rRNA测序及多样性分析 土壤细菌的16S rRNA测序由北京诺禾致源科技股份有限公司完成。采用十六烷基三甲基溴化铵(cetyltrimethylammonium bromide,CTAB)法提取样本总基因组DNA。利用特异性引物扩增土壤细菌16S rRNA V3+V4区片段,采用通用引物338 F(5′-ACT CCT ACG GGA GGC AGCA-3′)和806 R(5′-GGA CTA CHV GGG TWT CTA AT-3′)扩增,产物用2%琼脂糖凝胶电泳检测,AXYGEN凝胶电泳切胶回收,扩增体系和反应程序参考Huws等[16]的方法,每个样品3个重复。根据所扩增的16S区域特点,构建小片段文库,基于Illumina NovaSeq测序平台对该文库进行双末端测序。经过Reads拼接过滤,可操作分类单元(operational taxonomic units,OTUs)聚类,进行物种注释及丰度分析。基于OUTs聚类注释结果,对其进行α多样性分析和β多样性分析。
1.4 数据分析
采用Excel 2019软件进行数据处理和作图,采用SPSS 17.0软件进行数据的统计分析(单因素方差分析),差异显著性检验用最小显著性差异法(least-significant difference, LSD),显著性水平设为0.05。
2 结果与分析
2.1 枣园生草对土壤理化性质和土壤酶活性的影响
由表1可知,与清耕处理相比,枣园生草处理降低了行间和根际土壤pH值、含盐量和容重,提高了行间和根际土壤含水量,以及全氮、有效磷、有效钾和有机质含量,均以长柔毛野豌豆处理作用最明显。行间和根际土壤pH值和有效磷含量在不同处理间无显著差异,行间和根际土壤有效钾和有机质含量在不同处理间存在显著差异。长柔毛野豌豆处理对土壤有机质和全氮含量较清耕处理的提高幅度最明显,行间土升幅为64.5%和71.9%,根际土升幅为57.3%和90.6%。

表1 枣园生草对土壤理化性质的影响Table 1 Effects of planting grass on soil quality characteristics in jujube orchard
如表2所示,枣园生草不同程度地提高了行间和根际土壤脲酶、蔗糖酶、过氧化氢酶、碱性磷酸酶和蛋白酶活性,均以长柔毛野豌豆处理提高幅度最高。与清耕处理相比,行间土分别提高了168.8%、16.6%、6.7%、52.3%、14.2%,根际土分别提高了104.5%、15.3%、10.3%、49.0%、17.1%,且行间和根际土壤的脲酶和碱性磷酸酶活性在不同处理间存在显著差异,行间土壤的过氧化氢酶在不同处理间无显著差异。可见长柔毛野豌豆有利于提高土壤脲酶和碱性磷酸酶活性。

表2 枣园生草对土壤酶活性的影响Table 2 Effects of planting grass on soil enzyme activities in jujube orchard
2.2 土壤细菌测序优化序列统计
土壤样本测序结果统计表明(表3),细菌共测定的有效序列条数为469 424,过滤掉低质量的序列后,得到优化序列总数为300 150,优化序列所占比例均大于61.0%。

表3 土壤样本细菌测序结果统计Table 3 Sequencing data statistics of soil samples of bacteria
稀释曲线表示高通量测序数据量是否涵盖样品中所有微生物类群,当曲线趋于平坦,说明测序数据量渐进合理,更多的数据量只会产生少量新的物种[17]。由图1可知,6个土壤样本的OTUs数目随测序数据量的增加呈先快速上升后缓慢上升的趋势,序列数量达36 220 时,各土壤样本稀释曲线均基本趋于平缓,表明测序深度合理,可以反映样品中的物种组成。
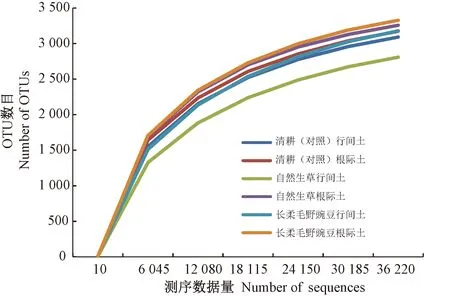
图1 土壤样本细菌稀释曲线Fig.1 Rarefaction curves of bacteria of soil samples
2.3 枣园生草对土壤细菌群落结构的影响
2.3.1 土壤细菌α多样性(Alpha diversity)分析 α多样性反映样本内的微生物群落的丰度和多样性,其中香农指数反映样品中的分类总数及其占比,辛普森指数反映样品群落的多样性,Chao1指数和ACE指数反映样品中群落的丰度[18-19]。由表4可知,与清耕处理相比,自然生草处理显著降低了行间土壤的香农指数、Chao1指数和ACE指数,但对根际土壤香农指数、Chao1指数和ACE指数无显著影响;长柔毛野豌豆处理提高了行间和根际土壤的Chao1指数和ACE指数,对香农指数无显著影响。辛普森指数在各处理间均无显著差异。各处理的覆盖率均达到了97%以上,说明测序样本所构建的基因库可以有效地反映其多样性,结果可靠。

表4 土壤细菌群落α多样性统计Table 4 Statistics of soil bacterial community α diversity
2.3.2 土壤细菌群落β多样性分析 无度量多维标定法(nonmetric multidimensional scaling,NMDS)统计是以Bray-Curtis距离来分析的非线性模型,将样本的物种信息以点的形式反映在二维平面上。二维平面上点与点间的距离可体现不同样本间的差异程度,反映出样本的组间和组内差异等。由图2可知,自然生草根际土和长柔毛野豌豆行间土样本组内差异相对较小,对照根际土样本组内差异相对较大。与清耕处理相比,生草处理土壤的细菌多样性产生了一定的差异,从行间土细菌多样性来看,自然生草行间土样本与清耕处理土壤的细菌多样性差异较大;从根际土细菌多样性来看,自然生草根际土样本与清耕处理土壤的细菌多样性差异较大。
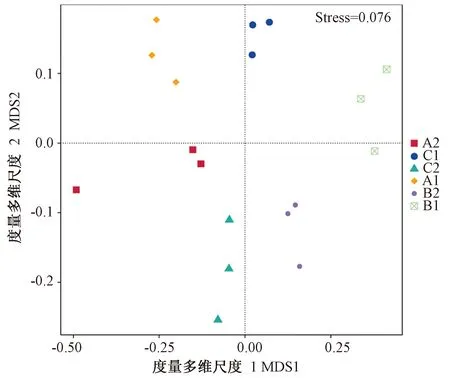
注: A1:对照行间土;A2:对照根际土;B1:自然生草行间土;B2:自然生草根际土;C1:长柔毛野豌豆行间土;C2:长柔毛野豌豆根际土。下同。Note: A1: Inter row soil of control. A2: Rhizosphere soil of control. B1: Inter row soil of natural grass. B2: Rhizosphere soil of natural grass. C1: Inter row soil of hairy vetch. C2: Rhizosphere soil of hairy vetch. The same as following.图2 土壤细菌群落β多样性分析Fig.2 Analysis of soil bacterial community β diversity
2.4 枣园生草对土壤细菌群落组成及相对丰度的影响
由图3可知,枣园生草对土壤细菌的群落组成及相对丰度影响较大。在门水平上,相对丰度排名前十的细菌类群为:变形菌门(Proteobacteria,23.7%~45.2%)、放线菌门(Actinobacteriota,12.2%~17.9%)、未明确细菌门(unidentified bacteria,10.6%~17.1%)、酸杆菌门(Acidobacteriota,6.5%~12.9%)、厚壁菌门(Firmicutes,3.3%~8.0%)、绿弯菌门(Chloroflexi,2.3%~5.9%)、粘球菌门(Myxococcota,1.4%~3.8%)、硝化螺旋菌门(Nitrospirota,1.2%~2.7%)、疣微菌门(Verrucomicrobiota,0.9%~3.0%)、泉古菌门(Crenarchaeota,0.5%~2.3%)。变形菌门、放线菌门和酸杆菌门为优势种群,其相对丰度之和占42.4%~76.0%。其中变形菌门以自然生草行间土壤样本最高(45.2%),放线菌门以长柔毛野豌豆行间土壤样本最高(17.9%),酸杆菌门以对照根际土壤样本最高(12.9%)。

图3 门分类水平土壤样本细菌群落相对丰度Fig.3 Relative abundance of soil bacterial community at phylum level
进一步从纲水平进行分析,土壤中主要细菌类群为:γ变形菌纲(Gammaproteobacteria,17.0%~38.8%)、放线菌纲(Actinobacteria,5.7%~12.2%)、α变形菌纲(Alphaproteobacteria,6.4%~8.5%)、芽孢杆菌纲(Bacilli,2.1%~8.5%)、未明确细菌纲(unidentified bacteria,2.8%~6.7%)、病毒杆菌纲(Vicinamibacteria,1.2%~5.9%)、嗜热油菌纲(Thermoleophilia,1.5%~4.7%)、酸杆菌纲(Acidobacteriae,2.1%~3.3%)、亚硝基球菌纲(Nitrososphaeria,0.5%~2.3%)、酸微菌纲(Acidimicrobiia,0.4%~2.9%)。优势菌纲为γ变形菌纲、放线菌纲、α变形菌纲和芽孢杆菌纲,其相对丰度占31.2%~68.0%。其中γ变形菌纲以自然生草行间土壤样本最高(38.8%),放线菌纲以自然生草行间和长柔毛野豌豆行间土壤样本最高(12.2%、12.0%),α变形菌纲和芽孢杆菌纲均以长柔毛野豌豆行间土壤样本最高(8.5%、8.4%)。
2.5 土壤细菌群落与土壤主要环境因子之间的关系
土壤细菌微生物群落结构与土壤理化性状的关系如图4所示。冗余分析(redundancy analysis)RDA1解释了土壤微生物群落差异的53.2%,不论行间土还是根际土的冗余分析,清耕与自然生草和长柔毛野豌豆处理在此轴分开,且pH值、容重与RDA1轴和清耕的夹角均为锐角,表明pH值和容重是清耕与自然生草和长柔毛野豌豆处理细菌微生物群落差异的主要影响因子。此外,长柔毛野豌豆和自然生草处理与脲酶、有机质、碱性磷酸酶、全氮、含水量、有效钾均呈锐角,表明这些因素对枣园生草(长柔毛野豌豆和自然生草)的细菌群落形成上发挥主导作用。

注:CK:清耕;NG:自然生草;HV:长柔毛野豌豆;TN:全氮;AK:有效钾;OM:有机质;BD:容重;SWC:含水量;URE:脲酶;PHO:碱性磷酸酶。Note: CK: Bare earth. NG: Natural grass. HV: Hairy vetch. TN: Total nitrogen. AK: Available potassium. OM: Organic matter. BD: Bulk density. SWC: Water content. URE: Urease. PHO: Phosphatase. 图4 不同生草处理土壤主要环境因子与细菌门水平的冗余分析Fig.4 Redundancy analysis of abundance phyla and main soil environmental factors
2.6 枣园生草对果实产量和品质的影响
由表5可知,与清耕相比,长柔毛野豌豆处理可显著提高果实单果重、可溶性糖含量、Vc含量,降低有机酸含量,但与自然生草相比无显著差异。果实纵横径和硬度在各处理间无显著差异;类黄酮含量在各处理间差异显著,以长柔毛野豌豆处理最高。

表5 枣园生草对果实产量和品质的影响Table 5 Effects of planting grass on fruit yield and quality in jujube orchard
3 讨论
我国3 000余年精耕细作的农耕文明及其“除草务尽”的传统理念,成为枣园生草技术不能在中国有效推广的主要障碍因素,将会导致果园土壤质量持续下降的恶性循环,因此筛选符合我国国情的草种对生草技术的推广将大有裨益。近几年,长柔毛野豌豆以茎木质化极低和无需刈割等突出优势,成为我国乃至世界范围内果园备受推崇的优势草种[20-21]。前人的研究多集中在长柔毛野豌豆对果园土壤养分[12]、病虫害防治[13]、树体生长和果实品质[22]的影响。本研究以传统清耕模式为对照,同时加入了自然生草处理,进一步探讨了枣园种植长柔毛野豌豆的土壤环境因子与土壤细菌群落之间的互作关系。结果表明,果园生草较清耕处理有效提高了土壤综合肥力和土壤细菌群落的丰度与多样性,其中种植长柔毛野豌处理较自然生草处理又进一步提高了土壤营养水平,表明长柔毛野豌豆可作为今后果园生草中的优势草种。进一步RDA分析表明,生草处理主要通过影响土壤全氮含量、有机质含量、含水量及碱性磷酸酶和脲酶活性,从而有利于增加土壤放线菌门、疣微菌门和变形菌门等有益细菌的丰度,这与姜莉莉等[23]在苹果园中研究认为生草有助于提高土壤有机质及养分含量,改善土壤微生态环境的结论一致。
土壤有机质含量是评价土壤营养的核心指标[24]。有机质含量提高可促进土壤其他指标同步提高,有利于果园的可持续发展。研究表明我国大部分果园有机质含量不足1%,氮肥利用率不足20%,若有机质含量达到1.5%,氮肥利用率可上升至30%[25]。本研究表明果园生草处理全面提高了土壤综合肥力,清耕与自然生草处理相比,长柔毛野豌豆处理显著提高了土壤有机质含量、全氮含量、有效钾含量和土壤酶活性,与陈学森等[12]在苹果园中的研究结论一致。但是在苹果园种植长柔毛野豌豆提高了土壤有效磷含量,而本试验中各处理间的有效磷含量未见显著差异,可能是由于磷素在土壤中迁移慢,极易被土壤吸附固定,大多形成不能被植物吸收的无效态磷形式存在[26],在苹果园中进行的是多年生草试验,而本研究一年生草试验尚不能改变土壤有效磷含量。土壤中的酶在土壤碳氮循环过程中发挥着重要作用,直接或间接参与了土壤中的物质代谢[27]。本研究发现果园生草不同程度地提高了土壤酶活性,其中长柔毛野豌豆显著提高了土壤脲酶和碱性磷酸酶活性,与吴玉森等[6]在梨园中的研究结论一致。人工生草避免了传统清耕制条件下施肥后的奢侈吸收和施肥间隔期间的养分匮乏,同时长柔毛野豌豆的根系密度是果树的几十倍,速效性肥料撒施后会被草根迅速吸收利用,避免了肥料淋溶损失,提高了养分生物贮存量以及无机态向有机态的转化,减少养分损失,因此可以充当养分的临时贮存库,随着根系死亡又会将养分释放到土壤中供果树根系吸收利用[28]。本试验是在枣果实完熟期进行土壤取样,可见种植长柔毛野豌豆使土壤养分供应稳定持久,这种稳定供肥的能力在保肥能力较差的山东省北部盐碱地枣园显得尤为重要。
土壤微生物群落结构及多样性对土壤生态系统的平衡起着重要作用,是评价土壤品质的重要生物学指标。有研究表明生草可显著提高果园土壤细菌数量[29]。本研究结果表明,长柔毛野豌豆处理较清耕和自然生草处理的物种丰度和多样性更高。其中长柔毛野豌豆处理的Chao1指数和ACE指数均高于清耕和自然生草处理,表明长柔毛野豌豆能够提高果园土壤细菌丰度。但清耕处理的ACE指数和Chao1指数略高于自然生草处理,可能因为试验处理时间较短,而且自然生草群落中夹杂恶性杂草,破换了土壤生态平衡,但是从长期来看,合理管理的自然生草制应优于清耕制度[30]。另外,本研究中3个生草处理的根际土细菌丰度均高于行间土,与Li等[31]认为土壤根际细菌数量和活性显著高于根外土壤,呈现特异根际效应的结论一致。进一步对各生草处理中的细菌在门和纲分类水平上进行物种注释,分析发现变形菌门、放线菌门和酸杆菌门为各处理下的土壤优势菌门,与王晓雯等[32]的研究结果一致。在纲分类水平上,优势菌纲为γ变形菌纲、放线菌纲、α变形菌纲和芽孢杆菌纲,多是变形菌的不同种类,可能变形菌主导了各分类水平下的优势菌种。与在雷竹林土壤[33]上的研究结论一致。同时生草处理也影响了土壤细菌群落的β多样性,生草处理下土壤细菌的物种组成和群落结构均异于清耕处理(对照),不论行间土还是根际土的细菌群落多样性与对照相比,生草处理均提高了土壤细菌群落多样性,自然生草差异较大,长柔毛野豌豆次之。
土壤微生物的群落组成及多样性极易受土壤自身及外界环境等综合因素的影响,是土壤质量变化的敏感指标[34]。RDA分析表明,土壤环境因子受长柔毛野豌豆处理影响较大,提高了土壤养分水平,从而有利于增加土壤放线菌门、疣微菌门和变形菌门等有益细菌的丰度。大部分放线菌门是一种腐生菌类,可分解木质素和纤维素,促进土壤中植物有机残体的降解[35];大部分疣微菌门有效参与了土壤氮循环的硝化作用,可促进有机氮向无机氮转化[36];变形菌门下面的一些亚纲(α、β、γ变形菌亚纲)具有吸收土壤有机碳的能力[37]。可见,枣园种植长柔毛野豌豆是一种可行的技术措施。由于本研究时间较短,后续可开展多年种植长柔毛野豌豆试验进一步挖掘其益处。
4 结论
与土壤清耕和自然生草处理相比,枣园种植长柔毛野豌豆可提高土壤营养水平,冗余分析表明其主要通过影响土壤全氮含量、有机质含量、含水量、碱性磷酸酶和脲酶活性来改善土壤细菌群落,从而提高果实产量和品质。种植长柔毛野豌豆具有省力、高效的特点,该植物将是枣园生草可推荐的适宜草种。
